Строение и функции
Хоаны удлиненные; у примитивных терапсид они занимают переднее положение и разделены сошником, который в большинстве случаев становится непарным. У прогрессивных форм за счет выростов максилл и небных костей развивается вторичное небо, отодвигающее хоаны далеко назад. Обособление носовой полости от ротовой обеспечивает бесперебойное прохождение воздуха в легкие, обусловливая возможность длительной обработки пищи во рту. Чешуйчатая кость разрастается наружу. Верхняя крыловидная кость обычно несколько расширена, у продвинутых форм входит в стенки мозговой коробки. Фланги крыловидных костей хорошо развиты. Базисфеноид обычно с маленькими базиптеригоидными отростками. Небные зубы редуцируются и обычно исчезают. Крыловидные кости тесно примыкают к основанию черепа, широко соприкасаются впереди, и межптеригоидные ямы зачастую полностью закрываются. Квадратные ветви крыловидных костей тонкие, часто редуцируются и теряют связь с квадратной костью. Квадратная и квадратноскуловая кости малы. Передняя ушная кость увеличивается и соединяется с нисходящими отростками теменной кости. Слуховая косточка (стремя) короткая, со слабым верхним отростком. Челюстное сочленение обычно расположено впереди или на одном уровне с затылочным мыщелком.
У прогрессивных форм зубная кость сильно увеличивается и образует на верхнем краю высокий венечный отросток, увеличивающий площадь крепления челюстной мускулатуры. Задние кости нижней челюсти (сочленовная, угловая, надугловая, венечная), называемые постдентальными, уменьшаются в размерах. Угловая кость с хорошо выраженной вырезкой. У наиболее продвинутых форм задний конец зубной кости входит в контакт с чешуйчатой костью, таким образом, образуя дополнительное челюстное сочленение.
Зубная система обычно дифференцированная: отчетливо выражены резцы, клыки и заклыковые (щечные) зубы. Эта геродонтность связана с выполнением зубами различных функций: схватывания добычи, ее удержания, размельчения и т. д. Изначально простые конические заклыковые зубы у продвинутых форм приобретают дополнительные бугорки на коронке и иногда дифференцируются на предкоренные и коренные. У продвинутых форм наблюдается однократная смена зубов, как у большинства млекопитающих. Иногда зубы могут отсутствовать.
Позвонки амфицельные или платицельные, обычно отверстия для хорды нет. Обычно имеется около 26 предкрестцовых (в том числе 5-7 шейных и порядка 20 туловищных) и 3-5 крестцовых позвонков. У прогрессивных форм дифференцируется поясничный отдел позвоночника, теряющий ребра. Интерцентры, как правило, отсутствуют, но иногда сохраняются между шейными позвонками. Хвостовой отдел позвоночника претерпевает редукцию.
Клейтрума обычно нет. Сохраняется короткая, но широкая межключица. Суставная ямка плечевого сустава становится правильно вогнутой; передний коракоид лишь в незначительной степени участвует в ее образовании; задний коракоид большой; супрагленоидное отверстие отсутствует. Грудина обычно окостеневает. Пластина подвздошной кости широкая. Лобковая кость укороченная, тироидное отверстие в большинстве случаев хорошо развито. Плечевая кость с овальной проксимальной головкой и слабо скрученным телом. Энтэпикондилярное отверстие имеется, но эктэпикондилярное может утрачиваться. Задняя конечность смещена под туловище (у продвинутых форм занимает парасагиттальное положение), ее движение осуществлялось в передне-заднем направлении. Проксимальная головка бедренной кости смещается несколько внутрь от тела кости, IV-й вертлуг редуцируется. В стопе имеется лишь 1 центральная кость. Фланговая формула обычно редуцирована до 2, 3, 3, 3, 3, но иногда сохраняется и большее, нормальное для рептилий число фаланг. Гастралии почти всегда отсутствуют.
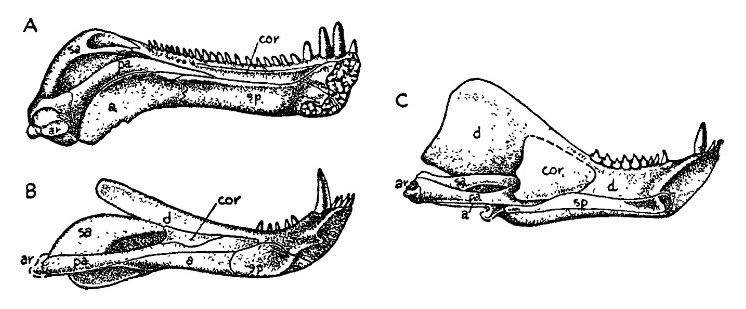
Прорисовки реконструированных нижних челюстей синапсид, изнутри: А — сфенакодонт Dimetrodon; В — горгонопид Scylacognathus; С — цинодонт Cynognathus.
Нерешенным вопросом для почти всех групп терапсид является способ функционирования их ушного аппарата, В настоящее время большинством исследователей принимается отсутствие, по крайней мере, у примитивных терапсид, постквадратной барабанной перепонки и подвижной слуховой косточки и формирование у потомков, млекопитающих, структур среднего уха преимущественно из костей нижней челюсти. Л.П. Татаринова пришел к выводу о вероятном возникновении барабанной перепонки лишь у высших цинодонтов в специальной вырезке угловой кости нижней челюсти, формируемой ангулярным лепестком. По мнению Л.П. Татаринова, у примитивных терапсид барабанной перепонки могло не быть, полость среднего уха соединялась с особым мандибулярным дивертикулом, располагающимся по нижнему краю нижней челюсти и связанным с полостью, прикрывавшейся имеющимся только у терапсид выростом угловой кости — ангулярным лепестком. Это тонкая костная пластинка, косо сзади-сверху вниз-вперед проходящая по наружной поверхности угловой кости.
Видимо, ангулярная пластинка терапсид — разрастание заднего края внешнего гребня (lamina reflexa), прикрывающего у сфенакодонтных пеликозавров небольшое коническое вдавление. Под эту пластину многие исследователи помещают часть птеригоидной мускулатуры. Однако исследование пластины у наиболее примитивных терапсид (Syodon и Estemmenosuchus) показывает, что полость под ней разделена гребнем на 2 части — узкую, каналообразную верхне-переднюю и широкую нижне-заднюю. Нижне-задняя в той или иной мере выступает ниже нижнего края челюсти, открыта с внутренней стороны и вполне могла быть связана с птеригоидной мускулатурой. Верхне-передняя открывается на верхнем краю ангулярного лепестка полукруглой вырезкой, продолжаясь пологим желобом вдоль верхнего края нижней челюсти. Именно эта полость значительно варьирует в относительных размерах и форме у различных групп, вплоть до сохранения от нее только передней стенки вырезки (Gorgonopidae).
Судя по приводимым в литературе данным, именно данная полость преформует в будущем тимпанальную часть слухового аппарата млекопитающих. Остальные элементы будущего слухового аппарата млекопитающих у терапсид имеют так же мало подходящее для этого строение — массивная слуховая косточка неподвижна и тесно скреплена с квадратной костью, квадратная кость (=incus) и сочленовная (=malleus) образуют обычный блок челюстного сустава. Видимо, расшифровка возможностей этой необычной конструкции как звукопередающего аппарата является делом специальных исследований, а пока следует отметить следующее: 1) сама вариабельность строения области ангулярного лепестка у различных групп заставляет предполагать активное функционирование структуры; 2) у примитивных групп, даже со слабо развитым пахиостозом скуловой области (например, у Biarmosuchus) над описанной выше вырезкой и продолжающим ее коротким желобом нависает сильно утолщенный, ругозистый край скуловой дуги (особенно хорошо это заметно у форм с сильным пахиостозом данной области — например, у Ulemosaurus); внутренний край "карниза" гладкий, как бы продолжает желоб от вырезки к нижнему краю затылочного фланга чешуйчатой кости, где он, соосно с челюстным мыщелком поворачивая внутрь и вверх, попадает в положение, в котором обычно реконструируют наружный слуховой проход у высших терапсид. Возможно, все эти данные позволяют полагать, что воздушную систему звукопроведения имели даже самые примитивные группы терапсид, пусть пока и неясно ее функционирование.

Остатки кожного покрова головы эстемменозуха (Estemmenosuchus uralensis): 1 — внешний вид мелкобугорчатого рельефа эпидермиса; 2 — внешний вид кориума с чечевицами, расположенными на соединительной основе; 3 — продольный разрез двух чечевиц с разрушенными верхними участками; 4 — диаметральный разрез чечевицы с участком внутреннего сегментированного слоя.
Существенно важным представляется вопрос о возможном строении кожи терапсид. Кожа маммального типа у прямых потомков терапсид не позволяет предполагать наличие у последних настоящего рептильного изолирующего фолидоза. Однако, по-видимому, в эволюции группы происходили изменения и в строении кожного покрова. Так, для примитивных групп (Eotitanosuchidae из диноцефалов, Estemmenosuchida и примитивных Gorgonopidae из горгонопов) отмечены своеобразные округлые ямки на некоторых участках поверхности костей черепа. У горгонопа Estemmenosuchus описаны залегающие в толще кожи линзовидные образования из кальцита, трактуемые как сохранившиеся остатки кожных желез. Специальный просмотр образцов в коллекции позволил выявить связь этих линзовидных образований с отмеченными ямками на поверхности кости. И "остатки кожи", и ямки имелись только на тех участках костей, где кожа должна была плотно прилегать к ним (особенно в районах лицевых пластин максилл, а также на рогообразных выростах), и полностью отсутствуют на внутричерепных поверхностях костей или в местах прикрепления мускулатуры, что в значительной мере подтверждает вывод о сохранении каких-то структур кожи. Пока трудно судить, что представляли собой эти кальцитизированные линзочки, но, во всяком случае, они не могли иметь отношения к нормальной рептильной чешуе.
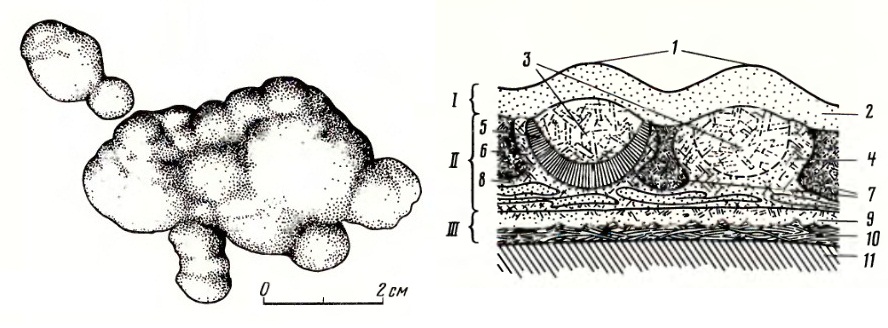
Слева — кожные окостенения эстемменозуха (Estemmenosuchus uralensis), с наружной стороны. Справа — предполагаемый схематический разрез кожи головы эстемменозуха (Estemmenosuchus uralensis): I — эпидермис, II — кориум, III — подкожная соединительная ткань, 1 — поверхностные бугорки эпидермиса, 2 — недифференцированный темно-коричневый эпидермис, 3 — полости чечевиц, выполненные светлым кальцитом, 4 — недифференцированная стенка чечевицы, 5 — внутренний сегментированный слой золотистого кальцита, 6 — наружный слой серого кальцита, 7 — участок между смежными чечевицами, выполненный темно-коричневым золотистым кальцитом, 8 — слой серого кальцита в основании чечевицы, 9 — слой светло-коричневого, иногда неясноволокнистого кальцита, 10 — слои параллельноволокнистого серого или золотистого кальцита, 11 — компактная костная ткань.
У примитивных титанозухий Syodontidae лицевые поверхности верхнечелюстных костей покрыты мелкими "морщинами", образованными многочисленными мелкими отверстиями, связанными анастомозными желобками. У высших диноцефалов и у аномодонтов поверхности лицевых пластин максилл гладкие, с немногочисленными крупными отверстиями, от которых отходят короткие прямые желобки, редко ветвящиеся или анастомозирующие; ближе к границе с предчелюстной костью число отверстий увеличивается, появляются ветвящиеся и анастомозирующие желобки. У горгонопий могут быть ямки скульптуры (например, у Suchogorgon) или такие же отверстия с желобками (например, у Sauroctonus). Хорошо развитые анастомозные сети сосудов на передних краях верхнечелюстных костей описаны у тероцефалов и цинодонтов, и интерпретированы как отпечатки сосудистой сети, питавшей вибриссы.

Мумифицированные остатки и детали строения кожных покровов Lystrosaurus.
Очень интересна скульптура боковых поверхностей костных выростов "рогов" на черепах Estemmenosuchus и бурнетиида Proburnetia. Крупные сосуды, выходя из отверстий на теменных костях, оплетают их ветвящимися желобами, образуя картину, очень напоминающую ветвление сосудов на пантах оленей.
У многих дицинодонтов (например, Kannemeyeriidae) на носовых костях имеются пахиостозные бугры со своеобразной микроструктурой поверхности, образуемой многочисленными анастомозирующими сосудами, очень похожие по строению на бугры основания рога современных носорогов. У дицинодонта Lystrosaurus вокруг глазниц имеются костные валики, что весьма напоминает структуры, например, у некоторых полорогих, как бы выдвигающие глаза из густой шерсти. Разумеется, все это не свидетельствует непременно о наличии у дицинодонтов настоящей шерсти, однако у рептилий подобные структуры не наблюдаются.

Мумифицированная кисть Lystrosaurus.
Все перечисленные факты, хотя и косвенно, все же свидетельствуют в пользу предположения об отсутствии рогового фолидоза рептильного типа даже у наиболее примитивных терапсид. Возможно, детальное изучение более обширного материала позволит обоснованно восстановить картину эволюции кожного покрова терапсид к "маммальному" состоянию. Сейчас можно лишь высказать предположение о вероятном наличии в той или иной мере развитого покрова, близкого по структуре к шерстному, у терозавров, высших аномодонтов, и, может быть, даже у высших горгонопов и диноцефалов, тогда как у наиболее примитивных представителей этих групп, видимо, кожа имела несколько иное строение, и была покрыта многочисленными тесно расположенными кожными железами.
Конечно, для сколько-нибудь обоснованных реконструкций данных явно мало, однако предварительную гипотезу высказать можно. Вряд ли следует предполагать, что эти железы выделяли слизистый секрет, как у лиссамфибий. Скорее, может быть, они продуцировали кератиноподобное вещество, бляшки которого сплошным паркетом покрывали тело, защищая от высыхания. Усиление функционирования таких желез могло привести сначала к появлению волосовидных сенсорных "вибрисс" на определенных участках тела, а затем и к формированию сплошного шерстного покрова. В сочетании с развитием потовых желез образовался "промаммальный" покров. На каком эволюционном уровне это происходило, сказать пока трудно, однако кажется вероятным, что исчезновение сплошного защитного "фолидоза" у разных групп терапсид связано, скорее всего, не с ослаблением защитных функций кожи против высыхания, а с их совершенствованием, и именно по пути "маммализации" кожи. Возникшие сначала остевая шерсть и потовые железы создавали воздушно-паровую прослойку. Образование подшерстка могло быть связано уже с появлением гомойотермности и соответствовать маммальному уровню. Соответственно, терапсиды, особенно апикальные группы эволюционных трендов, могут рассматриваться как потенциально высоконаземные группы животных.
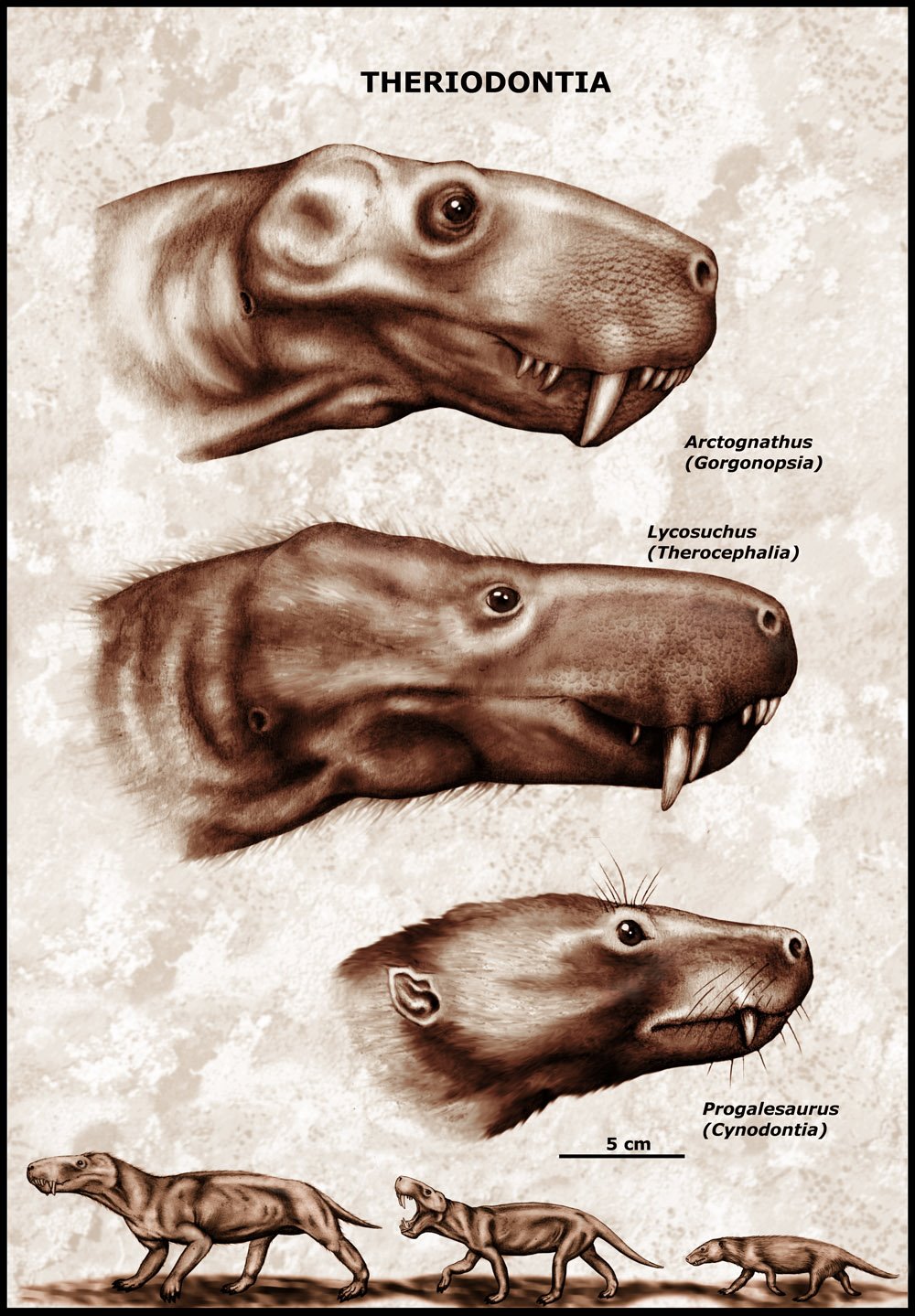
Реконструкция внешнего облика некоторых терапсид.
Значительные размеры и масса крупнейших синапсид (диноцефалов, эстемменозухов, дицинодонтов) позволяют предположить, что эти животные были гигантотермическими. Они имели низкую площадь поверхности к массе тела, в результате чего внешние слои ткани изолировали внутренние слои. Это увеличивало среднюю температуру животного, повышая уровень его метаболизма до близкого к гомойотермному существу, без обычных для этого адаптаций. Для охлаждения крупные терапсиды могли погружаться в воду или отводить избыточное тепло через различные выросты на голове, пронизанные множеством сосудов и покрытые мягкой кожей. Именно для этого могли предназначаться рога, вздутия и гребни на черепах диноцефалов, эстемменозухов и бурнетиоидов. Проходящая по ним кровь остужалась. Возможно, черепные выросты даже "потели". Однако главным назначением этих образований все-таки являлось привлечение самок, поскольку они больше развиты у самцов.
Подсчет содержания изотопов кислорода в костях пермских и триасовых терапсид из Южной Африки показал, что по-настоящему гомойотермными (на уровне современных однопроходных) могли быть только дицинодонты и цинодонты, да и то лишь триасовые. Но настоящая гомойотермия появилась только у млекопитающих (хотя, вероятно, не с самого начала).
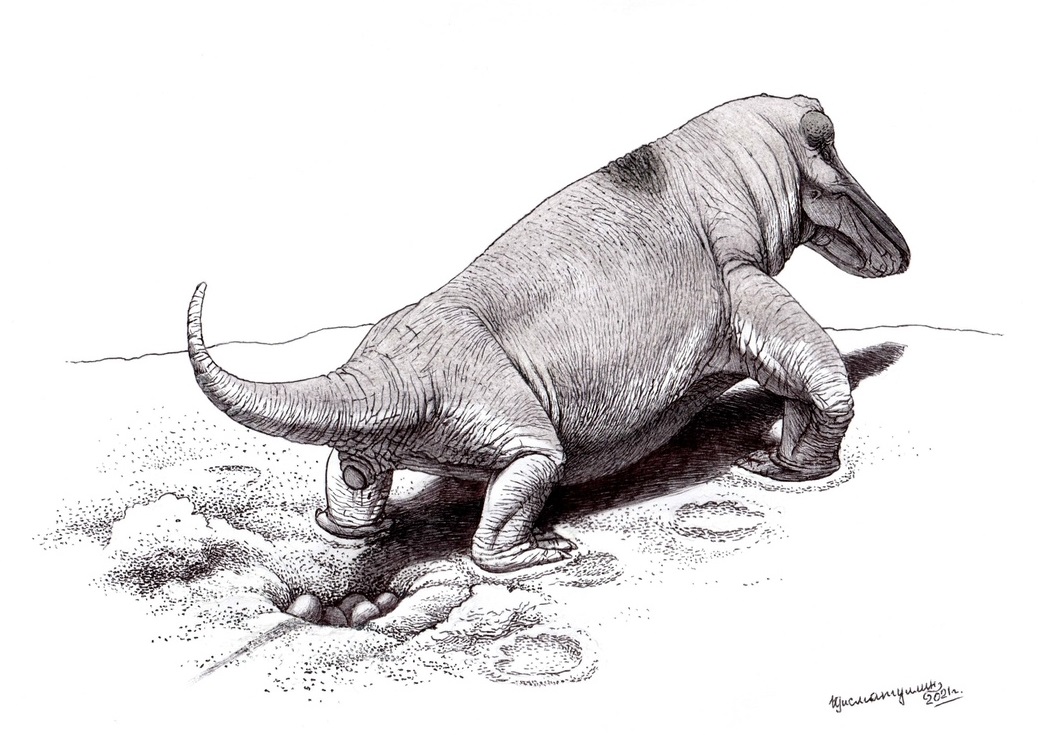
Струтиоцефал (Struthiocephalus whaitsi) откладывает яйца. Автор — И. Гисматтулин.
Есть основания предполагать, что у высших терапсид уже были сформированы мягкие, снабженные зачаточной мускулатурой губы, способствующие удержанию пищи в ротовой полости при ее пережевывании (наличие губ иногда принимается в качестве показателя перехода к сосанию молока). Утрата поясничными позвонками ребер предполагает приобретение диафрагмы и переход к маммальному механизму вентиляции легких. Передние конечности (как до этого задние) смещаются под туловище, в парасагиттальную плоскость, что говорит о способности к длительной локомоторной активности.
Большую роль в становлении млекопитающих сыграли норы. Их рыли триасовые цинодонты, мелкие пермские дицинодонты, а также Lystrosaurus. По-видимому, они откладывали там яйца и там же выхаживали потомство. Когда началось молочное вскармливание детенышей, неизвестно. Обычно его предполагают для цинодонтов, порой также для тероцефалов.

Дииктодоны (Diictodon feliceps) у входа в нору. Автор — Ntvtiko.