Строение и функции
По своей величине синапсиды сильно варьируют: общая длина составляет от 15 см до 6 м, вес — от 10 гр до 5 т.
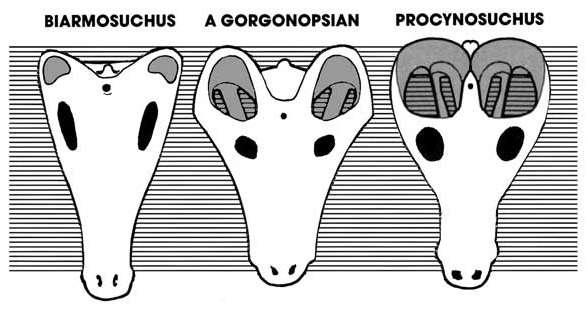
Ввиду интенсивной и длительной эволюции, существует мало общих для синапсид признаков строения скелета. Наиболее характерным из них является наличие с каждой стороны их черепа единственного височного окна, ограниченного снизу скуловой дугой. Эта дуга сложена, как правило, скуловой и чешуйчатой костями, а в некоторых случаях (Varanopidae, Cotylorhynchus) — скуловой и квадратно-скуловой костями. Первоначально височное окно было небольшим и занимало низкое положение на черепе. Над ним смыкались заднеглазничная и чешуйчатая кости; у более прогрессивных форм, однако, оно увеличилось и распространилось наверх, до теменной кости. Насколько известно, лишь у специализированного ядовитого тероцефала Euchambersia скуловая дуга редуцирована и височное окно открыто снизу.
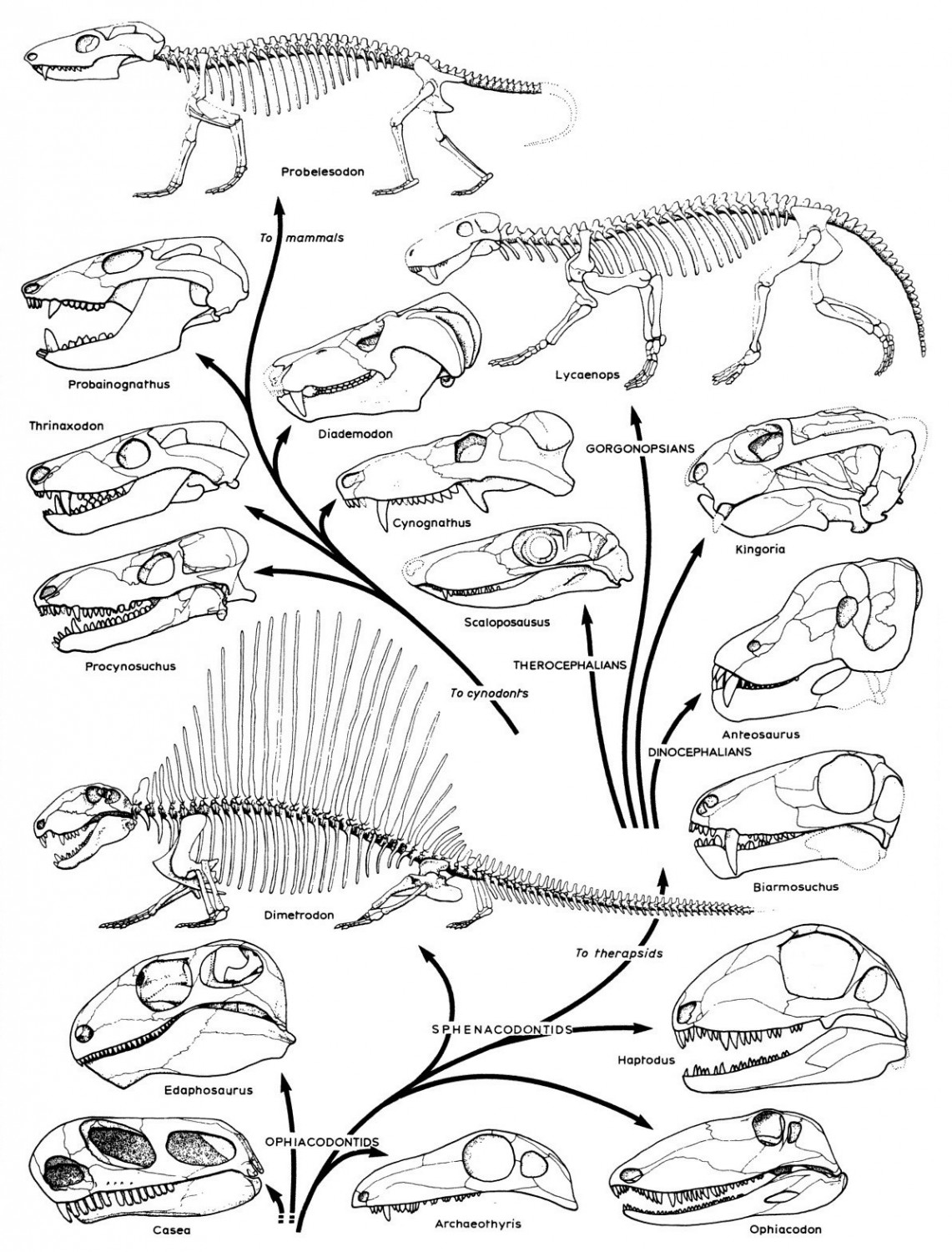
Филогенетическое древо синапсид, демонстрирующее эволюцию их черепа и скелета. В нижней части схемы показаны пеликозавры, посередине и вверху справа терепсиды, вверху слева цинодонты.
Череп синапсид никогда не бывает сильно уплощенным. Как правило, морда округлая. Ноздри терминальные. Глазницы обращены наружу, обычно с умеренно развитым кольцом склеротикальных косточек. У ряда продвинутых форм заглазничная дуга исчезает. Пинеальное отверстие большое у примитивных синапсид, маленькое или совсем исчезает у продвинутых. У современных рептилий в этом отверстии расположен пинеальный орган, который воспринимает интенсивность света (но не может давать изображение) и работает как эндокринная железа, участвуя в регуляции многих суточных и сезонных ритмов и, возможно, в терморегуляции.
Черепные элементы редуцируются сравнительно незначительно, но кожные супраоцципиталии (в большинстве случаев сросшиеся в верхнюю затылочную кость) были вместе с табличатыми костями оттеснены вниз к задней стороне черепа, а у прогрессивных форм сократилось число элементов, окружающих глазницу. Лобные кости достигают верхних краев глазниц. У примитивных форм заглазничная кость контактирует с надвисочной. У более продвинутых синапсид надвисочная кость обычно редуцирована или утрачена. Прогрессивные формы утрачивают и заднелобную кость, а квадратная и квадратноскуловая кости у них сильно редуцируются.
Прогрессивное увеличение височных окон у синапсид. Автор — Д. Петерс.
Затылок широкий; таблитчатые кости и непарная заднетеменная кость значительно разрастаются по его поверхности. Верхняя затылочная кость широкая и достигает таблитчатых; задневисочные отверстия маленькие. Заднеушная кость обычно не срастается с боковой затылочной. Околозатылочные отростки заднеушных костей массивные и в большинстве случаев несколько загибаются на конце вниз, по направлению к верхнему концу чешуйчатой кости. Затылочный мыщелок одинарный, двойной или тройной, с различной степенью разделенности. У высших синапсид мыщелок двойной.
Септомаксилла в большинстве случаев выходит на поверхность черепа позади ноздри. Почти всегда имеется морфологически подвижное базиптеригоидное сочленение. Небо вогнутое, у архаичных форм по типу близкое к небу капториноморф, с хорошо развитыми межптеригоидными отростками на базисфеноиде. У терапсид межптеригоидные ямы уменьшаются, а иногда совсем утрачиваются (горгонопиды) и птеригоиды широко соприкасаются по средней линии. Птеригоиды обычно с хорошо развитыми боковыми отростками и "флангами". Наружный птеригоид сохраняется. Верхний птеригоид хорошо развит, и у прогрессивных терапсид он значительно расширен. Сфенэтмоид обычно развит сравнительно хорошо.
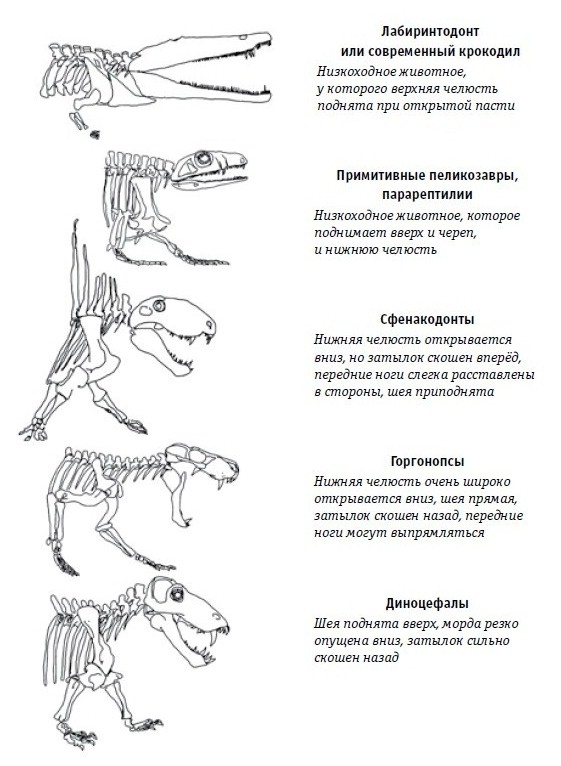
Схема, показывающая, как расположен затылок и как открывается пасть у различных тетрапод (по Богданову, 2018 г.).
У многих терапсид образуется зачаточное вторичное небо. Оно существенно укрепило верхнюю челюсть и надежно обособило носовую и ротовую полости, позволяя непрерывно дышать при пережевывании пищи. Кроме того, обработка пищи во рту сокращало количество времени, необходимое для пищеварения. У диноцефалов и примитивных горгонопов на небе появились костные выросты крыловидных костей — они поперек перекрывали ротовую полость при закрытой пасти. Растительноядные диноцефалы благодаря этому приспособились пережевывать пищу передними зубами. Но уже у горгонопид, а также у продвинутых дицинодонтов и тероцефалов мягкие ткани разрослись на небе, отделив ротовую полость от дыхательных путей, а твердое небо высокое. У высших тероцефалов и цинодонтов, независимо друг от друга, вторичное небо развилось очень хорошо, будучи образовано смыкающимися по средней линии отростками челюстных и небных костей. Обычно развитие вторичного неба связывают с гомойотермностью.
Какой бы то ни было ушной вырезки нет. Овальное окно слуховой капсулы расположено низко у основания черепа. Слуховая косточка (стремя) прободенная, обычно массивная и выступает наружу, проходя от овального окна к квадратной кости. Барабанная перепонка появилась, по-видимому, лишь у самых продвинутых терапсид, помещаясь в вырезке угловой кости близ задней стороны нижней челюсти.
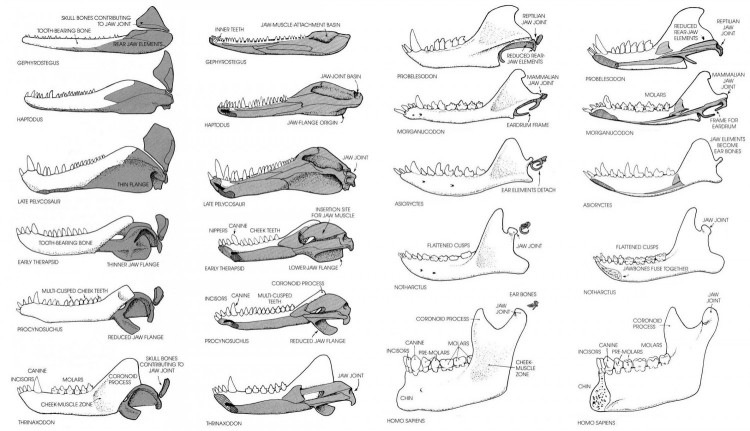
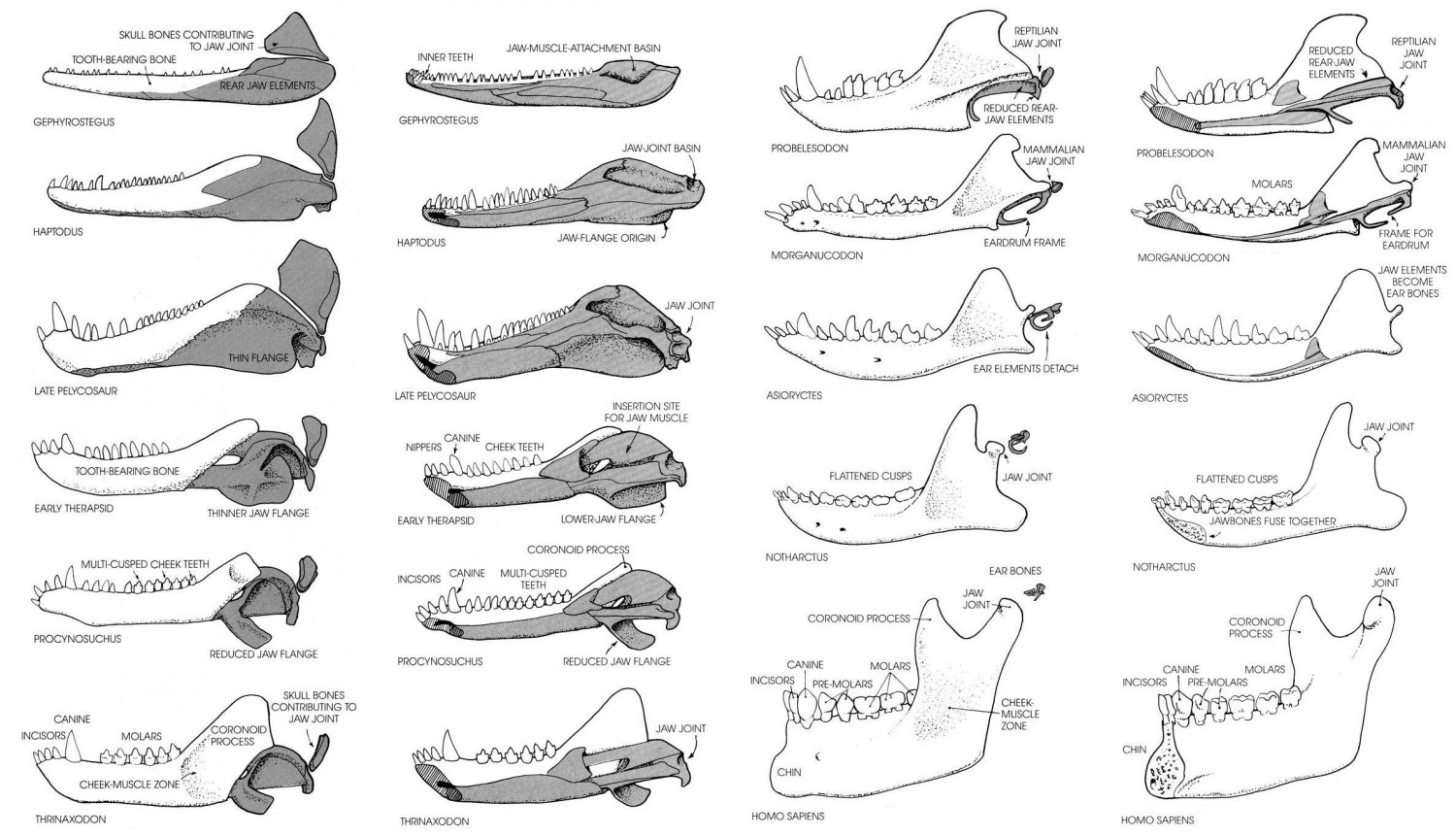
Эволюция костей нижней челюсти и среднего уха.
Нижняя челюсть у синапсид имеет полный набор костей, характерный для ранних амниот. С черепом она сочленяется челюстным суставом, образованным (как и у других амниот, но не у млекопитающих) квадратной и сочленовной костями. У терозавров зубная кость резко разрастается назад и образует мощный венечный отросток, повышая прочность нижней челюсти, а ее остальные кости (угловая, надугловая и сочленовная) постепенно редуцируются до совсем маленьких размеров. Лишившись своей исконной роли, задние нижнечелюстные кости со временем исчезают или начинают выполнять другие функции: например, сочленовная кость в ходе эволюции стала молоточком в составе среднего уха. Очевидно, в перестройке челюстного аппарата сыграла роль его миниатюризация и реорганизация жевательной мускулатуры. У цинодонтов Tritheledontidae доросшая до черепа зубная кость вступает в контакт с чешуйчатой костью, расположенной спереди от квадратной, и может образовывать с ней дополнительное челюстное сочленение. У млекопитающих на смену квадратно-сочленовному суставу, типичному для синапсид, приходит вторичный челюстной сустав, образуемый чешуйчатой и зубной костями. Традиционно именно наличие единственного чешуйчато-зубного сустава рассматривают как ведущий признак, позволяющий провести грань между классами синапсид и млекопитающих.
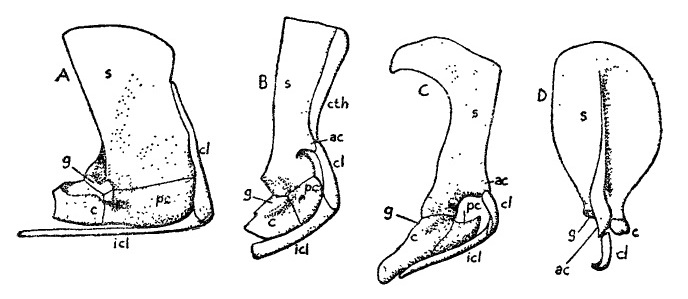
Прорисовки плечевого пояса синапсид и млекопитающих: А — пеликозавр
Ophiacodon, В — терапсид
Kannemeyeria, С — однопроходное
Ornithorhynchus, D — сумчатое
Didelphis. Сокращения: ac — акромиальный отросток (acromion), c — коракоид (coracoideum), cl — ключица (clavicula), cth — клейтрум (cleithrum), g — гленоидная ямка, pr — прокоракоид (procoracoideum), s — лопатка (scapula).
Зубы субтекодонтные у наиболее примитивных представителей (некоторые пеликозавры), текодонтные — у более высоко организованных. У примитивных пеликозавров и многих растительноядных форм (эдафозавров, дромазавров и др.) зубы гомодонтные. Однако уже у более продвинутых пеликозавров они обнаруживают начальные этапы дифференцировки: зубы, располагавшиеся на предчелюстных костях, напоминают по форме резцы, далее следуют хорошо выраженные клыки и сохраняющие исходную коническую форму заклыковые зубы. Более выраженный характер дифференцировка зубов приобретает у терапсид; четко выражена она у терозавров, причем если у горгонопов заклыковые зубы остаются простыми коническими (а иногда вовсе исчезают), то у цинодонтов они имеют сложно устроенную жевательную поверхность. У некоторых диноцефалов функцию измельчения пищи принимают не заклыковые зубы, а резцы, или вернее, предклыковые зубы. У большинства синапсид отмечается многократная и нерегулярная смена зубов на протяжении всей жизни (полифиодонтия), но у высших терозавров намечается переход к двукратной смене (дифиодонтии). У пеликозавров и низших терапсид имеются зубы на небе, иногда очень сильно развитые (особенно у эдафозавров). У дицинодонтов зубы замещаются роговым клювообразным чехлом.
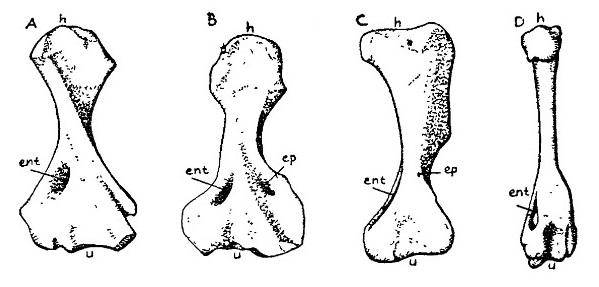
Прорисовки плечевой кости синапсид и млекопитающих, сверху: А — пеликозавр Dimetrodon; В — пеликозавр Edaphosaurus; С — цинодонт ?Diademodon; D — олигоценовый амфиционид Daphoenus. Сокращения: ent — энтепикондиляоное отверстие (foramen entepicondylare); ер — эктепикондиляоное отверстие (foramen ectepicondylare); h — головка; u — сочленованная поверхность для локтевой кости (ulna).
Обычно насчитывается 26-27 предкрестцовых позвонков. Число шейных позвонков составляет 6-7, но задние из них слабо отличаются от спинных. Как у большинства примитивных рептилий, атлант сохраняет самостоятельные невральные дуги; более необычно сохранение самостоятельных невральных дуг проатланта. Обычно обособливается и поясничный отдел с укороченными и прирастающими к позвонкам ребрами. Поясничный отдел особенно хорошо выражен у высших терапсид и, очевидно, свидетельствует о развитии у них мышечной диафрагмы. В крестце от 2-3 (у примитивных форм) до 8 позвонков. Хвост длинный, средней длины или укороченный.
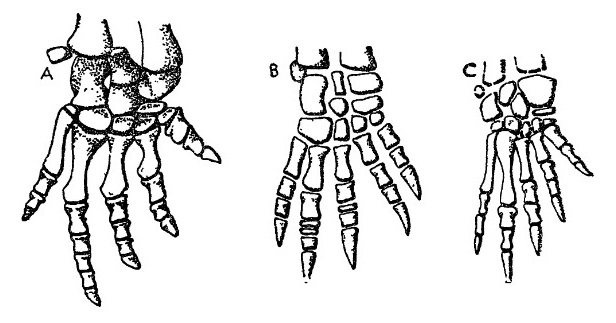
Прорисовки скелета кисти синапсид: А — пеликозавр Ophiacodon; В — горгоноп Lycaenops; С — тероцефал Ericiolacerta.
Позвонки изначально амфицельные, у примитивных представителей — прободенные, а у некоторых высших синапсид — почти платицельные. У пеликозавров между позвонками обычно сохраняются свободные интерцентры. Зигапофизы наклонены вниз; остистые отростки расширенные в передне-заднем направлении и высокие, а у некоторых пеликозавров они резко разрастаются и выступают над спиной, образуя т. н. "парус". Ребра двухголовчатые (имеют двойное соединение с позвоночником), но на задних туловищных позвонках обе головки сливаются.
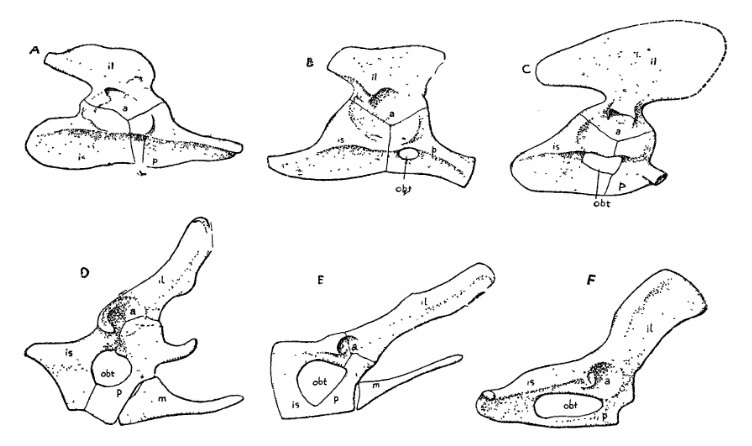
Прорисовки тазового пояса синапсид и млекопитающих: А — пеликозавр Dimetrodon; B — горгоноп Lycaenops; С — цинодонт Cynognathus; D — однопроходное млекопитающее Ornithorhynchus; Е — сумчатое млекопитающее Didelphys; F — миоценовый амфиционид Daphoenodon. Сокращения: a — acetabulum; il — ilium; m — сумчатая кость; obt — foramen obturatorium, увеличивающийся до размеров окна у терапсид и млекопитающих; р — pubis.
У пеликозавров постановка пконечностей сохраняет примитивный характер, проксимальные их элементы отходят от тела в горизонтальном направлении. У более высоко стоящих терапсид конечности по существу были уже приведены к передне-заднему положению, свойственному млекопитающим, в связи счем конечности и их пояса испытали ряд морфологических изменений. Плечевой пояс с 2 коракоидами и Т-образной межключицей, у примитивных синапсид сохраняется и клейтрум. На лопатке развивается акромиальный отросток. Подвздошная кость обычно расширенная, разрастается в переднем направлении. Вентральная часть тазового пояса пластинчатая, без вырезки между лонной и седалищной костями, но у высших синапсид в этом месте развивается тироидное отверстие. Конечности обычно массивные, но удлиненные. Плечевая кость с энтэпикондилярным, а иногда и с эктэпикондилярным отверстиями, локтевая кость с олекраноном. Дистальные суставы конечностей формируются на границе предплечья и кисти (предплечно-запястный), голени и стопы (голеностопный). Астрагал комплексный, полностью сформированный. Фаланговая формула у пеликозавров и большинства терапсид типа 2 - 3 - 4 - 5 - 3, но у аномодонтов и высших терозавров число фаланг становится таким же, как у млекопитающих — 2 - 3 - 3 - 3 - 3. В целом можно утверждать, что на протяжении всей истории синапсид совершенствовалась их локомоция, но лишь квадрупедальная; развития в сторону бипедализма здесь, в противоположность архозавроморфам, не наблюдалось.
Значительная массивность элементов скелета, особенно характерная для крупных синапсид (диноцефалов, дицинодонтов), может быть во многом связана с несовершенством костной ткани — меньшая прочность компенсировалась большей толщиной. У некоторых синапсид на верхней части туловища имелись кожные окостенения. Нередко сохраняются гастралии.
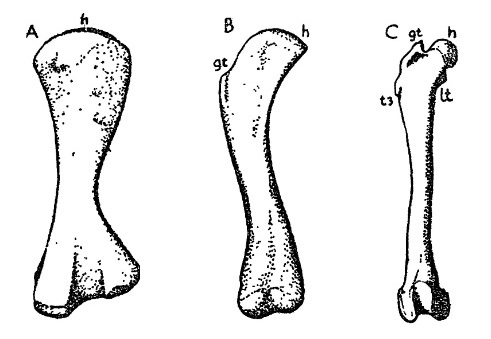
Прорисовки бедренной кости синапсид и млекопитающих: А — пеликозавр Ophiacodon; В — цинодонт; С — олигоценовый амфиционид Daphoenus. Сокращения: gt — большой трохантер; h — головка бедренной кости; lt — меньший трохантер; t3 — третий трохантер.
Физиология синапсид почти не известна. Неясно, например, значение у них теменного глаза, или роль спинного "паруса" многих пеликозавров. Высказываются предположения, что они служили целям терморегуляции.
Среднее ухо синапсид с системой выравнивания импедансов развилось совершенно иным путем, нежели у предков современных рептилий и птиц. У сфенакодонтных пеликозавров отогнутая пластина угловой кости, по-видимому, функционировала как барабанная перепонка. Ее вибрации передавались внутреннему уху через сочленовную, квадратную кости и стремя. У терапсид кости на заднем конце нижней челюсти не присоединялись к зубной швами и способны были двигаться самостоятельно, что облегчало проведение колебаний. Постдентальные кости постепенно редуцировались, но остались частью нижней челюсти даже у ранних млекопитающих. Форма и относительное расположение этих элементов недвусмысленно указывают, что квадратная, сочленовная и угловая кости рептилий гомологичны соответственно наковальне, молоточку и барабанной кости современных млекопитающих.
Вероятно, синапсидная ветвь тетрапод исходно отличалась строением кожных покровов от завропсид. Во всяком случае, надежных доказательств наличия настоящей твердой чешуей у пеликозавров не обнаружено. Кожа примитивных терапсид была гладкой и несла в своей толще большое количество желез. Возможно, выделяемый этими железами секрет загустевал и покрывал кожу мелкими шелушащимися "чешуйками". Вопрос о времени возникновения волосяного покрова у терапсид до сих пор не решен окончательно. Вибриссы ("усы") могли появиться достаточно рано; не исключается их наличие даже у дицинодонтов. Первоначально шерстный покров мог защищать железистую кожу от высыхания, а уже затем стал обеспечивать термоизоляцию. Согласно другой точке зрения, волосы изначально могли появиться в качестве средств осязания — таких, как вибриссы.
Есть основания полагать, что уже в конце перми у терапсид наметился переход к гомойотермности, и прогрессивные триасовые их представители обладали, по-видимому, такими характерными для млекопитающих особенностями, как относительная теплокровность, мышечная диафрагма, четырехкамерное сердце, шерстный покров и т. д. Размножение происходило путем откладывания яиц.
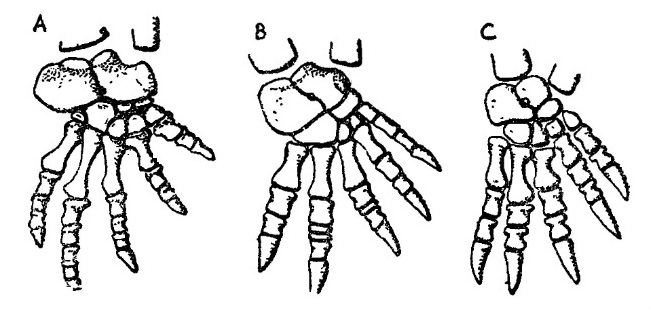
Прорисовки скелета стопы синапсид: А — пеликозавр Ophiacodon; В — горгоноп Lycaenops; C — тероцефал Theriognathus.
Образ жизни
Большинство синапсид — плотоядные; особенно интересна ветвь крупных горгонопид — древнейших саблезубых хищников. Однако часть пеликозавров (эдафозавриды и казеиды), а позднее и терапсид (часть диноцефалов и аномодонты) перешли к растительноядности. Зубочелюстной аппарат цинодонтов тритилодонтид отдаленно напоминает таковой грызунов. Известны тероцефалы с "ядовитыми" зубами.
Синапсиды заселяли преимущественно наземные местообитания, некоторые формы (офиакодонты, некоторые горгонопы, диноцефалы и дицинодонты) были полуводными, обитая по берегам пресноводных бассейнов, болот и лагун. Некоторые дицинодонты, судя по строению их конечностей, вели роющий образ жизни.