Современная систематика
Как было показано выше, на протяжении большей части XX в. синапсиды считались одним из подклассов класса рептилий. Позднее широкое распространение представлений, характерных для филогенетической систематики, привело к пересмотру классификации синапсид. Они были выведены из состава класса рептилий, получив в конце XX в. статус самостоятельного класса (в этой связи стоит напомнить, что А. Бринк еще в 1963 г. предлагал поднять таксономический ранг Synapsida до класса с 2 подклассами — Pelycosauria и Therapsida).
В то же время распространение взглядов, характерных для радикальных представителей кладистики, привело к тому, что многие современные авторы используют название Synapsida в расширенном смысле и применяют его ко всей кладе синапсидообразных (Synapsidomorpha), включающей не только синапсид в классическом понимании, но и млекопитающих (по отношению к данной кладе, помимо названия Synapsidomorpha, иногда применяют также названия Theropsida или Pan-Mammalia). При этом сторонники эволюционной таксономии выделяют в составе синапсидообразных 2 класса: парафилетический класс Synapsida и дочерний монофилетический класс Mammalia. Млекопитающих также вводят и в состав Therapsida, а термин Pelycosauria не используют — как относящийся к парафилетической группе. В целом же синапсидообразные, вместе с сестринской группой завропсид (Sauropsida), образуют кладу амниот — крупнейшую по объему группу в составе надкласса четвероногих (Tetrapoda).
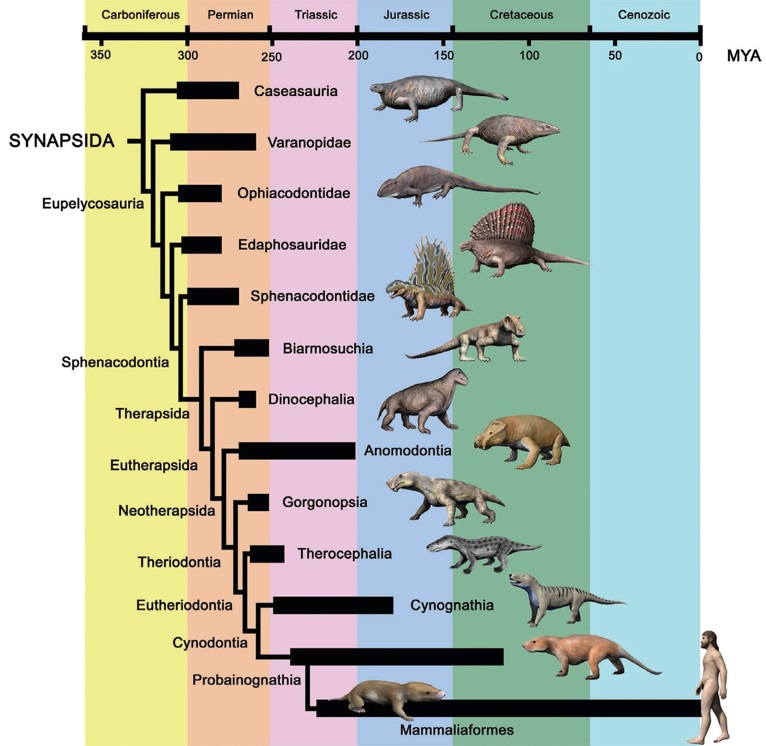
Кладограмма, представляющая родственные отношения синапсид согласно современным данным.
У других палеонтологов такой подход вызывает нарекания, поскольку для ссылок на таксоны Synapsida или Therapsida в их традиционном объеме приходится использовать громоздкие выражения "синапсиды, не являющиеся млекопитающими" (англ. nonmammalian Synapsida) и "терапсиды, не являющиеся млекопитающими" (англ. nonmammalian Therapsida); пеликозавров же при этом именуют "базальными синапсидами". Т. Кемп в этой связи отмечал, что парафилетичность Synapsida и Therapsida носит "технический" характер; при этом пеликозавры существенно отличаются от терапсид по многим признакам (а терапсиды — от млекопитающих), и в подобных ситуациях, когда виды, образующие парафилетическую группу, имеют по своему фенотипу достаточное сходство друг с другом при значительном отличии от представителей других групп, трактовка такой группы как таксона вполне законна. Сторонники данной точки зрения продолжают рассматривать Synapsida и Mammalia как отдельные классы.
Таким образом, на сегодняшний день в состав класса синапсид обычно включают 2 отряда — пеликозавров (Pelycosauria) и терапсид (Therapsida); в последний из них входят такие основные группы: биармозухии (Biarmosuchia), диноцефалы (Dinocephalia), аномодонты (Anomodontia) и териодонты, или зверозубые (Theriodontia). Некоторые авторы повышают ранг терапсид до подкласса, содержащего отряды Biarmosuchia, Dinocephalia, Anomodontia и надотряд Theriodontia; последний при этом, в свою очередь, включает отряды горгонопсов (Gorgonopsia) и настоящих териодонтов (Eutheriodontia). При этом оба отряда (как и весь класс Synapsida) являются парафилетическими группами. В основу разделения синапсид на крупные систематические категории принимается, как правило, строение черепа, нижней челюсти и зубной системы.
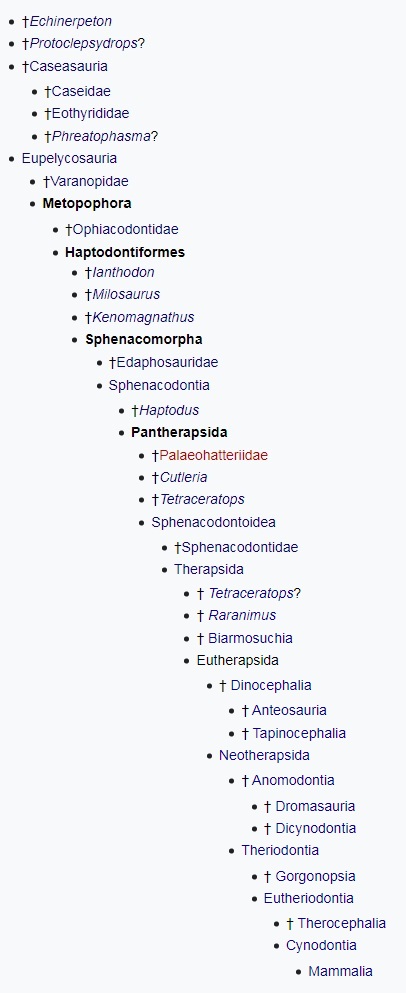
Систематика синапсид по современным данным.
Особая систематическая схема, в значительной степени эклектическая, была принята в коллективной монографии М.Ф. Ивахненко и ряда других российских авторов в 1997 г. Для подкласса Pelycosauria была использована схема Р. Рейса за 1986 г., но с перенесением «Varanopsidae» в Ophiacodontia. Подкласс Therapsida был разделен на 3 надотряда — Eotheriodontia, Theriodontia и Anomodontia. При этом наиболее неудачными надо признать принципы выделения надотряда Eotheriodontia, фактически в смысле схемы Э.К. Олсона за 1962 г., объединяющего все примитивные группы — Phthinosuchida (Phthinosuchoidea, Rhopalodontoidea, Niaftasuchida, Dinocephalia), поскольку жестких критериев разделения отрядов на тот момент не существовало.
Ревизия материалов по примитивным тероморфам Восточной Европы, проведенная М.Ф. Ивахненко в 1999 и 2000 гг., выявила, что темпоральная область черепов у эстемменозухид, фтинозухид и горгонопид построена по общей схеме — заглазничная кость не несет площадки налегания мускула, в отличие от Dinocephalia. С учетом строения темпоральной области черепа была предложена новая схема классификации отряда Eotheriodontia, противопоставленного отряду Eutheriodontia.
В 2001 г. и 2002 г. М.Ф. Ивахненко было показано, что в действительности Theromorpha в принятых схемах объединяются только по наличию синапсидного окна. По строению височных областей черепа Ophiacodontia (и Varanopidae) примитивно сохраняют полость между стенкой мозговой коробки и аддукторной ямой, латерально ограниченную структурами палатоквадратума (epipterygoideum, quadratum). У Sphenacodontia и примитивных групп (Gorgonopia, Dinocephalia, Anomodontia) латеральная стенка полости редуцирована с выходом аддукторной мускулатуры к боковой поверхности мозговой коробки. У высших групп (Therocephalia, Cynodontia) имеется эпиптериковая полость, однако в состав медиальной стенки аддукторной полости входит специальный фланг чешуйчатой кости. Поскольку у Sphenacodontia, Gorgonopia, Dinocephalia и Anomodontia чешуйчатая кость контактирует с заглазничной и подстилает теменной щит, эта конструкция обозначена как «параквадратобазальная», тогда как конструкция черепа Therocephalia и Cynodontia, у которых чешуйчатая кость не контактирует с заглазничной и перекрывает теменную, обозначена как «сквамозобазальная». Gorgonopia, Dinocephalia и Anomodontia были объединены в Eotheriodontia, высшие териодонты — в Eutheriodontia.
Новая широкая ревизия Theromorpha, проведенная М.Ф. Ивахненко в 2003 г., заставила его признать, что только наличие синапсидного окна как апоморфии по преформованной структуре не может быть принято в качестве надежного систематического признака. Ряд групп пеликозавров (Ophiacodontia и Casesauria) может быть противопоставлен всем Theromorpha sensu Ivakhnenko, 2003 вследствие отсутствия у них субапсидной вырезки и периангулярной полости на угловой кости. Аддукторная полость у Ophiacodontia и Casesauria (офиакодонтная конструкция по М.Ф. Ивахненко) отделена эпиптериковой щелью от латеральной стенки мозговой коробки. Медиальная стенка аддукторной полости сформирована пластиной палатоквдратума, представленной оссификациями широкого эпиптеригоида и высокой квадратной кости. Это соответствует строению данной области у большинства групп примитивных тетрапод, но не у Theromorpha.
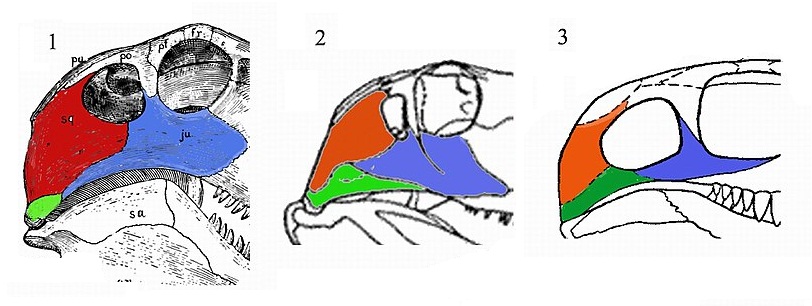
Схематическое сравнение черепов «пеликозавров»: 1 — сфенакодонт, 2 — офиакодонт, 3 — казеид. Квадратноскуловая кость — зеленая, чешуйчатая — красная, скуловая — синяя.
В то же время обычно принимаемые особенности строения, якобы объединяющие Pelycosauria и отличающие их в целом от Therapsida, по мнению М.Ф. Ивахненко, вполне могут являться результатом независимых и неравномерно проходивших преобразований в эволюции различных групп синапсид. Они соответствуют перестройкам челюстной мускулатуры (I), зубной системы (II) и области послехоанного воздухопроведения (III). С (I) связано увеличение относительных размеров височного окна и соответственно значительная редукция надвисочной кости; со (II) — увеличение размеров верхних клыкообразных зубов с увеличением высоты лицевой пластины верхнечелюстной кости и соответствующим удлинением септомаксиллы и дорсального отростка премаксиллы; с (III) — формирование носоглоточного желоба и соответствующее поперечное расширение сошников, уменьшение межптеригоидной вырезки, преобразование небных озубленных бугров и удлинение базикраниальных отростков птеригоидов.
Основываясь на упомянутых аргументах, М.Ф. Ивахненко выводит из состава Theromorpha «офиакодонтных пеликозавров» — отряд Ophiacodontia (надсемейство Varanopoidea с семействами Varanopidae и Watongiidae, а также надсемейство Ophiacodontoidea с семейством Ophiacodontidae) и отряд Casesauria (с семействами Eothyrididae и Caseidae), объединив их в едином таксоне Ophiacomorpha, условно ранга подкласса. «Истинные пеликозавры» Edaphosauridae и Sphenacodontidae были оставлены в составе примитивных Theromorpha.
Помимо этого, М.Ф. Ивахненко допускает, что таксон Ophiacomorpha может являться родственным предкам Archosauromorpha. Данное предположение обусловлено тем, что окно синапсидного типа, первично ограниченное заглазничной, чешуйчатой и скуловой костями (например, у Ophiacodon), у Varanopidae имеет тенденцию разрастаться по всей щеке, доходя до квадратно-скуловой кости (например, у Varanops). Топографически при этом оно перестает отличаться от нижнего височного окна диапсид. Поскольку никто не доказал, что оба окна диапсидной конструкции возникают одновременно, или верхнее опережает нижнее по времени возникновения, или нижнее первично является для всех групп субапсидным, то лишь наличие синапсидного окна может означать для группы и принадлежность к примитивным группам, близким к диапсидам, но до появления верхнего окна.
Кроме того, следует заметить, что офиакодонтная группа имеет ряд черт, удивительно сближающих их с примитивными архозавроморфами. Например, форма орбит у офиакодонтов со специфической дорсальной вырезкой верхнего века, как у всех диапсид. На парокципитальном отростке мозговой коробки образуется специальный фланг, формирующий у архозавроморф тимпанальную полость между затылком и стерноклейдомастоидной мускулатурой. Треугольное вдавление перед глазницей у Varanodon, ограниченное сзади косым гребнем, практически не отличается от начальных стадий формирования предглазничного окна примитивных архозавроморф (Ornithosuchus, Riojasuchus) и даже динозавров (Lesothosaurus). Наличие рудимента спиракулярной щели (апопарейальность) над каудальным отростком заглазничной кости достаточно легко может привести к формированию парапсидного окна, узкого и щелевидного у примитивных Archosauromorpha (Proterosuchus). Разумеется, это не означает, что Ophiacomorpha можно принимать в качестве предков Archosauromorpha, имеющих диапсидную силовую конструкцию черепа. У высших офиакоморф (Varanopoidea и Casesauria) череп имеет синапсидную конструкцию, с единым большим окном синапсидного типа. Однако примитивные Ophiacodontoidea вполне могут быть близки по строению к морфотипу, исходному для Archosauromorpha.
Для Ophiacomorpha М.Ф. Ивахненко принял следующее определение: апопарейальные ангуститабуляры с синапсидным височным окном; параквадратобазальные по типу строения темпоральной области черепа, надвисочная (=чешуйчатая) кость значительно редуцирована. Theromorpha определяются М.Ф. Ивахненко как апопарейальные ангуститабуляры с синапсидным и субапсидным височными окнами и периангулярной полостью на угловой кости нижней челюсти. Указанные дефиниции обозначают обладание четко определенными морфологическими признаками.
1. Ангуститабуляры (angustitabularia). Маркирующий признак — наличие контакта таблитчатой и теменной костей. Синдром связан с наличием раздельных экзопокровов межкапсулярной крыши и крыши затылочного кольца, поскольку tectum synoticum находится топографически под задней частью parietale, a tectum posterius полностью перекрыт postparietale.
2. Апопарейалы (apopareialia). Маркирующий признак — отсутствие специального широкого фланга чешуйчатой кости, подстилающего надвисочную; сохраняется примитивная конструкция с бесшовным прилеганием слабо скрепленных костей в области контакта крыши черепа и щеки, где проходила спиракулярная щель.
3. Субапсиды (subapsida). Маркирующий признак — отсутствие контакта скуловой и квадратно-скуловой костей. Связано с примитивной фенестрацией черепа, образованием окна (fenestra subapsida) или вырезки (incisura subapsida) в районе стыка швов скуловой, чешуйчатой и квадратно-скуловой костей. Для Theromorpha субапсидность черепа определяется наличием вырезки (incisura subapsida), что выражается в отсутствии контакта jugale и quadratojugale.
4. Синапсиды (synapsida). Маркирующий признак — наличие синапсидного височного окна в области рудиментарной спиракулярной щели между крышей и щекой. Окно расположено ниже каудального отростка заглазничной кости, скуловую дугу образует скуловая кость.
5. Периангулярии (periangularia). Маркирующий признак — наличие желоба на наружной поверхности угловой кости нижней челюсти. Желоб прикрыт снаружи тонким костным выростом (ala angularis), топографически продолжающим внешний гребень (lamina reflexa). Возможно, полость преформирована унаследованным костным желобом инфрадентального сейсмосенсорного канала и связана с аппаратом звукопроведения.
Для подкласса Theromorpha М.Ф. Ивахненко в 2003 г. принял разделение на 2 группы в ранге инфраклассов: Eotherapsida и Eutherapsida (=Eutheriodontia sensu Tatarinov, 1974; =Therosauria sensu Kemp, 1982).
Инфракласс Eotherapsida характеризуется редукцией эпиптериковой полости и реперным диагностическим признаком наличия fenestra anterosynapsida (параквадратобазальная конструкция). Эпиптеригоид тонкий, медиальной стенкой аддукторной ямы является латеральная стенка мозговой коробки, куда входят передняя пластина prooticum и единая вертикальная пластина (periotica) из срастающихся tectum posterius, tectum synoticum и opisthoticum. Параквадратобазальная конструкция определяется наличием чешуйчатой кости, лежащей топографически ниже crista parotica и дорсально подстилающей теменную кость. Имеется контакт каудального отростка заглазничной кости и чешуйчатой кости по верхнему краю височного окна.
Инфракласс Eutherapsida характеризуется сохранением эпиптериковой полости и реперным диагностическим признаком наличия fenestra posterosynapsida (сквамозобазальная конструкция). Эпиптериковая полость разделяет латеральную стенку мозговой коробки и медиальную стенку аддукторной ямы, состоящую из расширенного эпиптеригоида (= alisphenoideum) и переходящую в специальный фланг чешуйчатой кости. Сквамозобазальная конструкция определяется наличием чешуйчатой кости, лежащей топографически выше crista parotica и дорсально перекрывающей теменную кость. Контакт заглазничной и чешуйчатой костей отсутствует.
По мнению М.Ф. Ивахненко, это разделение должно было произойти весьма рано, в самый начальный период формирования височных окон, и могло быть связано с некоторыми отличиями в преимущественном развитии первичного окна — сзади вперед, с ограничением сзади "настоящей squamosum" (fenestra posterosynapsida), или спереди назад, с выходом к "paraquadratum - squamosum" (fenestra anterosynapsida).
Принимаемые М.Ф. Ивахненко альтернативные синдромы отобраны им по принципу наличия или отсутствия и не могут рассматриваться в развитии или неполном выражении. Действительно, иные соотношения экзо- и эндоструктур, чем у angustitabularia, будут означать принадлежность к иной группе; син- и апопарейальность означает возможность или невозможность фенестрации в районе рудимента спиракулярной щели, и даже при отсутствии реализации этой возможности, например, у стегальных капториноморф они остаются apopareialia. Следующие синдромы (subapsida, synapsida, periangularia) связаны с наличием соответствующих структур, а отсутствие любой из них будет также означать принадлежность к иной группе. Например, наличие только одного синапсидного окна (как у Ophiacodontia), также определяет синапсидную конструкцию. Но присутствие субапсидной вырезки и синапсидного окна одновременно (Theromorpha) определяет иную конструкцию, которую можно обозначить как синсубапсидную (synsubapsida). То же относится и к уровню разделения Eotherapsida и Eutherapsida (paraquadratobasalia — squamosobasalia). Возможно, в данном случае сохранение или редукция первичной палатоквадратной стенки cavum epiptericum и определяет строение обрамляющего экзоскелета.
В 2008 г. М.Ф. Ивахненко в очередной раз уточнил филогенетическую систему подкласса Theromorpha, подразделенного им на инфраклассы Eotherapsida и Therosauria. Эотерапсиды включают надотряды Sphenacomorpha и Dinomorpha. Последний разделяется на отряды Gorgodontia (с подотрядами Dinocephalia и Gorgonopia) и Anomodontia (с подотрядами Ulemicia и Dicynodontia). Терозавры, в свою очередь, включают отряды Therocephalia и Cynodontia.
Отметим, что воззрения М.Ф. Ивахненко на систематику синапсид распространения не получили. Ряд исследователей вслед за М.Ф. Ивахненко отмечает некоторую необычность форм, помещаемых им в Ophiacomorpha, однако установление для них особого подкласса большинство считает преждевременным. Л.П. Татаринов также высказал сомнение в правомерности выделения с точки зрения филогенетический системы в особый инфракласс Therosauria.