Органы чувств
В толще кожи на клюве и, в меньшей степени, на щитках позади него залегают рецепторы электрической и тактильной чувствительности, представляющие собой специализированные эпидермальные структуры с подходящими к ним нервными окончаниями. При этом большая часть механорецепторов сосредоточена в продольных канавках на клюве, тогда как 40 000 электрорецепторов распределены по его поверхности более равномерно, со средней плотностью около 30 на 1 кв. мм. Максимальной чувствительностью обладают не все рецепторы, а лишь определенная их часть. Электро- и механорецепторы обслуживаются парой очень сильно развитых тройничных (V-х черепных) нервов; периферические синапсы отсутствуют.
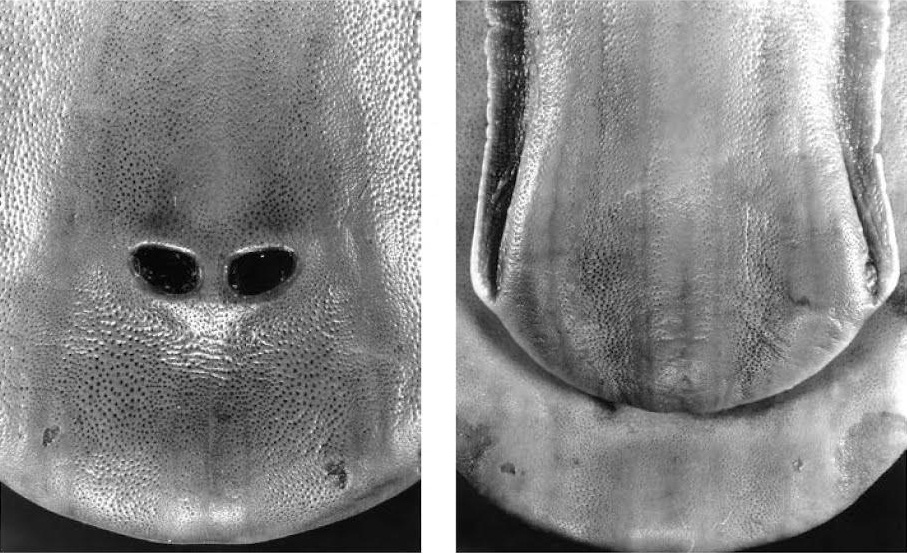
Клюв утконоса (Ornithorhynchus anatinus) крупным планом, вид сверху и снизу.
Полагают, что электрорецепторы являются специализированными эккриновыми железами. Они чередуются с несенсорными эккриновыми железами, которые не иннервированны и используются для создания на поверхности клюва слизистой пленки. Судя по всему, эта пленка способствует лучшей электропроводности и предохраняет рецепторы от высыхания, когда животное находится на суше. Механорецепторы выглядят как расположенные перпендикулярно к поверхности клюва стержневидные образования из выстроенных в ряд уплощенных чувствительных клеток. Они находятся в мелких порах, тогда как через более крупные поры на клюве открываются потовые железы. Иннервированные серозные железы расположены на внешней и внутренней поверхности клюва, группируясь главным образом по его краям и вблизи щечной области.
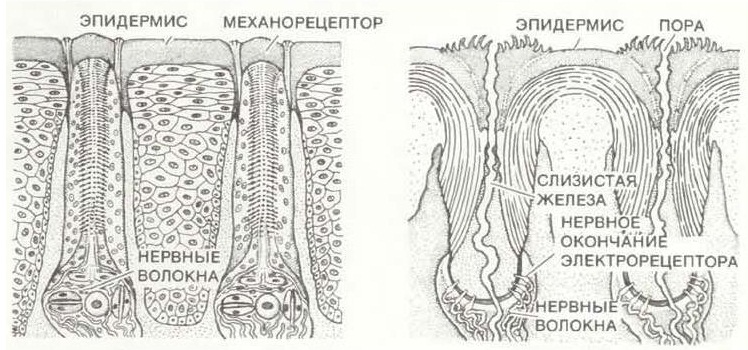
Рецепторы, залегающие в коже клюва утконоса (Ornithorhynchus anatinus): слева — механорецепторы, справа — электрорецепторы.
Любое живое существо генерирует вокруг себя слабое электромагнитное поле за счет сокращения своей мускулатуры. На это поле (напряжением от нескольких десятков микровольт до нескольких милливольт и с частотой колебаний около 140 Гц) и реагируют электрорецепторы охотящегося в воде утконоса, вследствие чего он плывет по направлению к источнику стимула. Судя по результатам экспериментов, животное привлекает даже искусственный объект, через который пропущен незначительный электрический ток. Вероятно, утконос определяет нужное для броска направление, анализируя различия в полярности и интенсивности сигналов, получаемых разными электрорецепторами. Это объясняет характерные движения головы животного из стороны в сторону во время поисков корма (примечательно, что аналогичные движения совершают молотоголовые акулы (Sphyrnidae) и веслоносые (Polyodontidae), также обладающие расположенными на голове электрочувствительными органами).
Выявляемые электрорецепторами поля позволяют утконосу отличать одушевленные объекты от неодушевленных, а тактильные раздражения от их движений в воде непрерывно стимулируют механорецепторы. Все сигналы передаются в соматосенсорную область коры головного мозга и в нужный момент челюсти производят быстрое движение, схватывая добычу. По-видимому, расстояние до нее утконос определяет, учитывая разницу во времени между моментами получения генерируемых жертвой электрических стимулов (распространяющихся в воде с большей скоростью) и вызванных ее движениями импульсов механического давления (относительно более медленных). Высказано предположение, что утконос способен улавливать даже очень слабые электрические сигналы. При большом количестве испускающих их источников сигналы суммируются, достигая требуемого порога для раздражения сенсорной коры головного мозга.
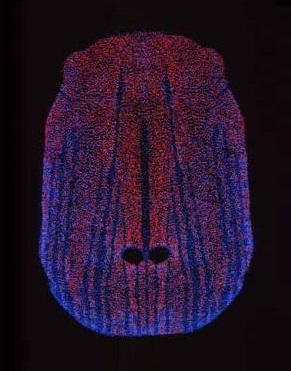
Электрорецепторы (показаны в виде красных точек) и механорецепторы (синие точки) на клюве утконоса (Ornithorhynchus anatinus).
Установлено, что даже при слабом тактильном раздражении механорецепторы клюва посылают в головной мозг нервные импульсы, вызывающие электрический потенциал на гораздо более обширном участке его новой коры, чем импульсы от конечностей, глаз и ушей. Если же воздействовать на клюв утконоса электрическим полем, то возбуждается область новой коры, почти полностью перекрывающаяся и выходящая за пределы той, которая возбуждается при раздражении механорецепторов. Более того, часть корковых нейронов возбуждается от сигналов как электро-, так и механорецепторов, что является дополнительным подтверждением тесной связи между обоими видами чувствительности. При этом нервные импульсы от рецепторов на правой стороне клюва поступают на левую сторону мозга и наоборот.
В целом клюв утконоса преобладает на соматотопической схеме его мозга точно так же, как кисти рук человека доминируют у «сенсорного гомункулуса». При этом большинство корковых нейронов представляет в коре очень ограниченные участки поверхности клюва. Двигательные области коры частично совпадают с чувствительными областями.
Считается, что электрическая чувствительность у предков утконоса развилась в связи с необходимостью поиска пищи в мутной воде и может быть связана с утратой зубов. Существует также допущение, что утконос использует электрические поля, создаваемые водными течениями, для определения своего положения относительно дна.
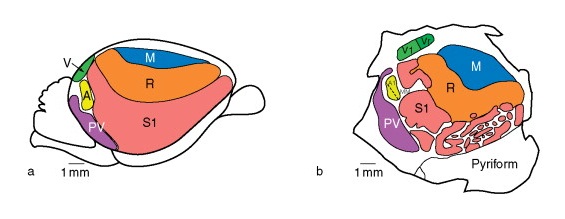
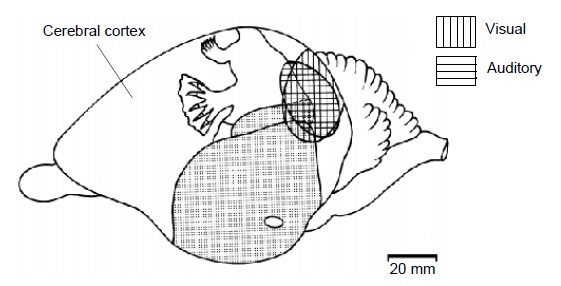
Вверху: схема распределения зон в больших полушариях головного мозга утконоса (Ornithorhynchus anatinus), сбоку (а) и спереди в разрезе (b).
Внизу: соматотопическая схема чувствительной коры больших полушарий головного мозга утконоса (Ornithorhynchus anatinus).
Зрение у утконоса довольно слабое. Сферическое глазное яблоко имеет небольшие размеры, достигая около 0,6 см в диаметре. Как и у ехидн, в задней части склеры расположено чашевидное образование из хряща. Зрачок круглый, с гладкомышечным сфинктером. Хрусталик сильно уплощен. Сосудистая оболочка очень толстая и глубоко пигментированная. Сетчатка содержит палочки и двойные колбочки с бесцветными капельками жира, однако чувствительность глаза к интенсивности света и степень его цветового зрения не определены. Рядом с глазным яблоком расположены слезная и гардерова железы. Подобно завропсидам и в отличие от большинства териев, имеется мигательная перепонка, которая очищает и смазывает глазную поверхность, двигаясь и рассеивая выделения желез. Снаружи глаз прикрывают 2 века, каждое из которых оснащено тарзальной пластинкой. В оптической хиазме перекрещивается большинство зрительных нервных волокон; лишь малая их часть продолжает идти по той же стороне мозга. Область коры, получающая и обрабатывающая зрительные сигналы, расположена в задней части полушария и имеет сравнительно небольшие размеры.
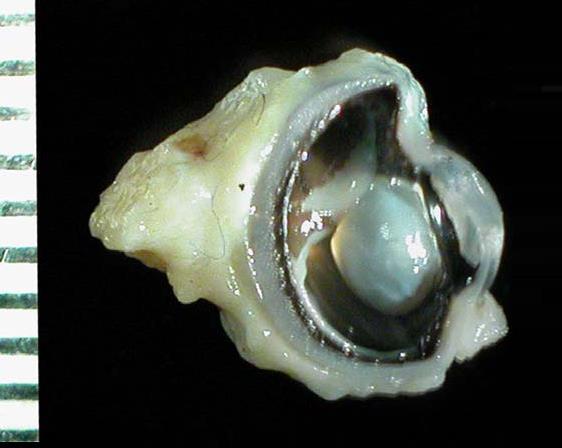
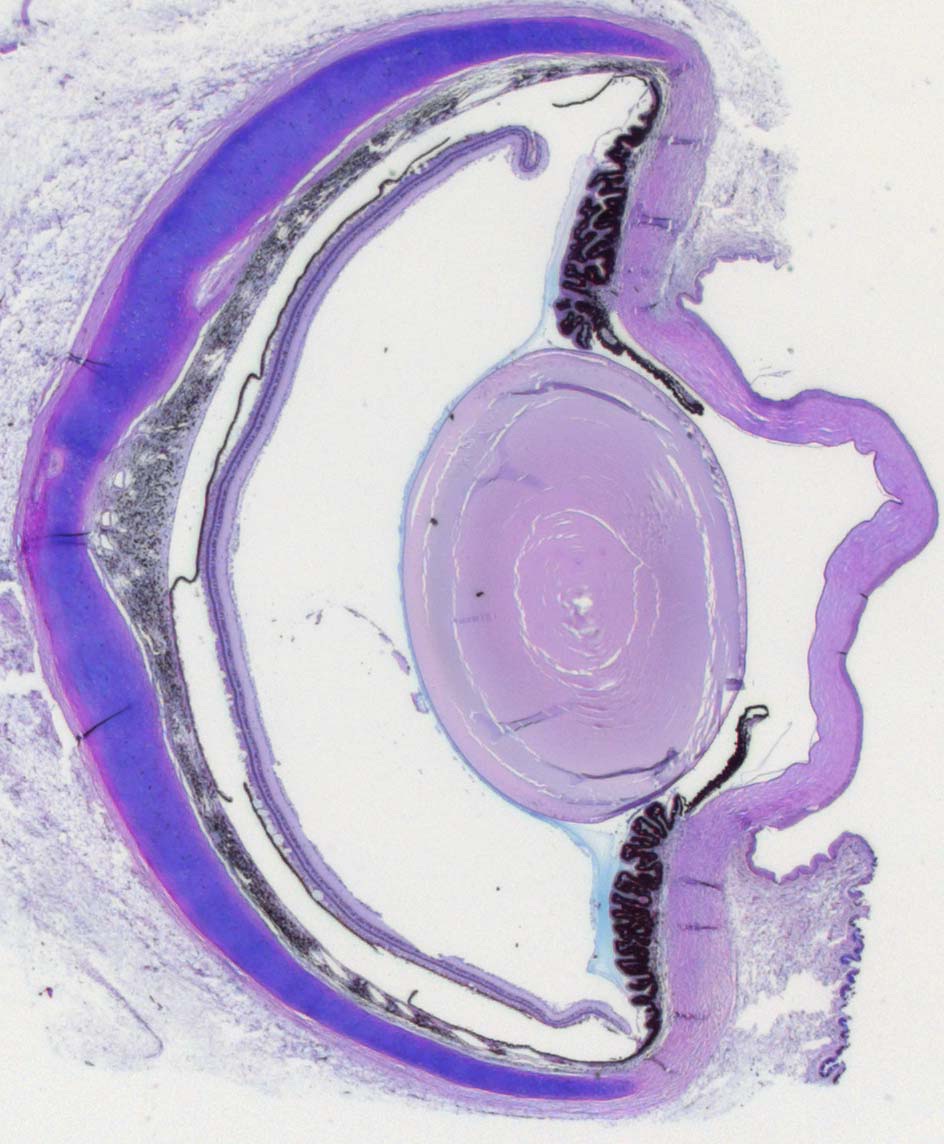
Слева — глазное ложе утконоса (Ornithorhynchus anatinus), справа — глазное яблоко утконоса в разрезе.
Так как глаза при нырянии прикрываются плотно сомкнутыми веками и краями глазно-ушных желобков, можно предположить, что зрение под водой зрение практически не используется. Наблюдения в прозрачном аквариуме показали, что ныряющий утконос на короткие мгновенья все-таки открывает глаза, но вряд ли это играет существенную роль, поскольку животное это активно в основном ночью и часто кормится в мутной воде, так что зрение оказывается фактически бесполезным. Так как относительно небольшие глаза утконоса расположены на дне желобков позади широкого лобного щитка высоко по бокам головы, он плохо видит прямо перед собой, когда находится на поверхности воды и на суше. Поскольку у животного не развита бинокулярность, отсутствует связанное с ним визуальное восприятие глубины. Тем не менее, в воздушной среде глаза весьма чувствительны к движениям, издали замечая потенциальную опасность. Таким образом, зрительный канал получения информации продолжает сохранять немаловажное значение.
Имеются указания на то, что у предков утконоса зрение играло гораздо более существенную роль. У утконоса роговица и прилегающая к ней передняя поверхность хрусталика относительно плоские, а его задняя поверхность круто изогнута. Данные свойства, а также месторасположение на сетчатке области наибольшей чувствительности (area centralis), весьма напоминают таковые у хорошо видящих под водой млекопитающих, например, выдр (Lutrinae) и морских львов (Otariinae). Височная концентрация (с ушной стороны) ганглионарных клеток сетчатки, важная для бинокулярности, тоже может указывать на преобладание в прошлом охоты на достаточно крупную добычу; наличествующей сегодня остроты зрения для подобных действий явно недостаточно. Кроме того, ограниченная острота зрения соответствует в головном мозге низкой степени кортикального увеличения, небольшому латеральному коленчатому телу и крупному верхнему двухолмию; это свидетельствует о том, что для зрения крыша среднего мозга играет более важную роль, чем зрительная кора больших полушарий, подобно некоторым грызунам. Перечисленные признаки подтверждают, что утконос последовательно приспосабливался к сумеречной активности и поискам пищи в мутной воде, развивая электрорецепцию в ущерб зрению.
Слух у утконоса сравнительно плохой. Звуковые волны попадают в ухо через довольно длинный (около 4 см) наружный слуховой проход, который простирается от поверхности тела до барабанной перепонки. По-видимому, способность косточек среднего уха воспринимать звуки в воздушной среде развита лучше, чем у ехидн, но все же очень ограничена. Когда животное ныряет, отверстие слухового прохода замыкается, так что слух вряд ли используется в водной среде. Высказано предположение, что наполненный воздухом слуховой проход может резонировать с передающимися в воде звуками, функционируя в качестве дополнительного органа слуха, наподобие плавательного пузыря рыб. Однако подтверждающие это экспериментальные данные отсутствуют. Ушная улитка способна улавливать звуковые волны частотой от 500 Гц до 20 кГц (диапазон почти такой же, как и у человека). Как и у ехидн, внутреннее ухо наиболее чувствительно к сравнительно низким колебаниям частотой около 4 кГц, хорошо проникающим сквозь толщу воды и кости черепа. Область коры головного мозга, получающая информацию по слуховому нерву, относительно небольшая и перекрывается с почти всей зрительной областью. В связи с полуводным образом жизни особенно хорошо развит вестибулярный аппарат.
Обоняние у утконоса развито хорошо. Количество обонятельных раковин (турбиналий) меньше, чем у ехидн. С каждой стороны в продырявленной пластинке (lamina cribrosa) имеется по одиночному отверстию, тогда как у прочих современных млекопитающих (включая ехидн) они множественные. Запахи улавливаются обонятельными рецепторами носовых раковин (около 700 видов), а также вомероназальными рецепторами типа 1 (V1R, порядка 950 видов) и типа 2 (V2R, около 80 видов). Полагают, что вомероназальные рецепторы не только воспринимают запах находящейся во рту пищи, но могут также иметь определенное значение для распознавания феромонов или запаховых веществ, растворенных в воде. Вполне вероятно, обоняние играет важную роль при общении самца с самкой и матери с детенышами, а также в ходе других социальных взаимодействий.