Внутренние органы
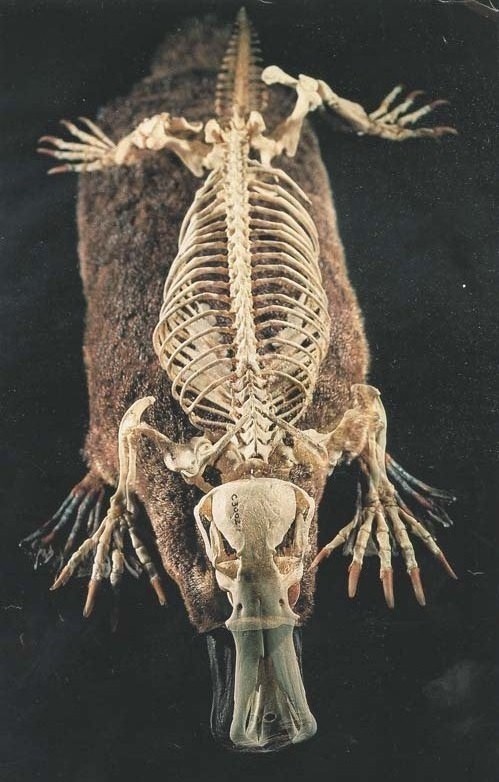

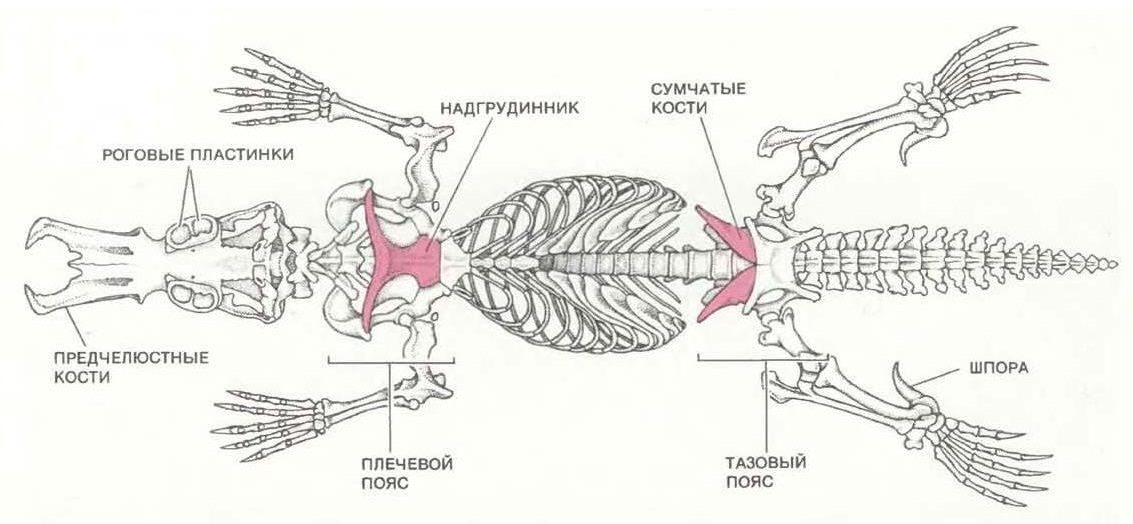
Скелеты утконоса (Ornithorhynchus anatinus) и прорисовка скелета, снизу.
Спереди и по бокам верхняя челюсть утконоса выступает над нижней. Верхнечелюстные кости широко расходятся в стороны. Септомаксиллярные и расположенные под ними межчелюстные кости значительно превосходят верхнечелюстные кости по длине и лирообразно загибаются внутрь. Маленькая гантелевидная кость находится в центре между ветвями клюва. Ее назначение точно не выяснено, однако отверстия вомероназального органа открываются с 2 сторон на сужении между расширенными участками этой непарной кости. Наружные ноздри по отношению к гантелевидной кости расположены антеро-латерально. Между отклоняющимися ветвями верхней челюстей находится хрящевая пластинка.
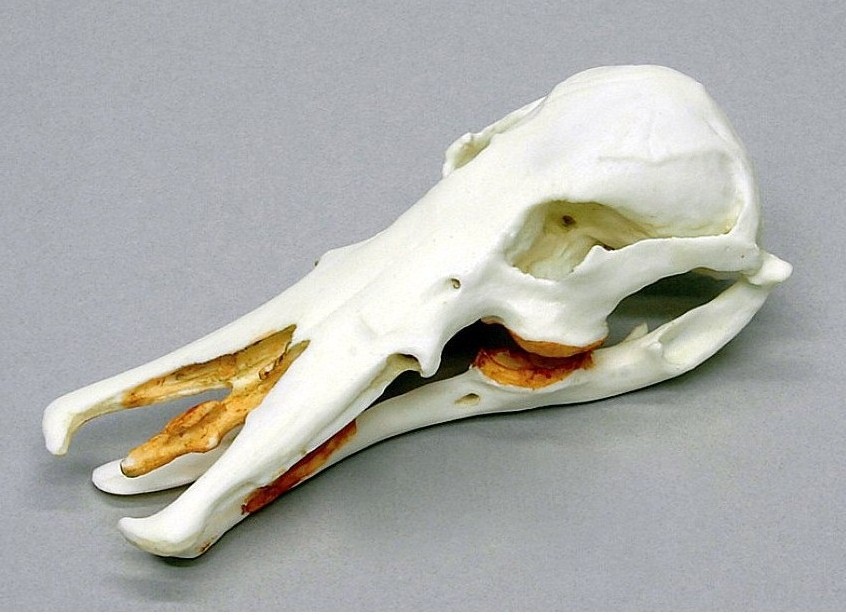
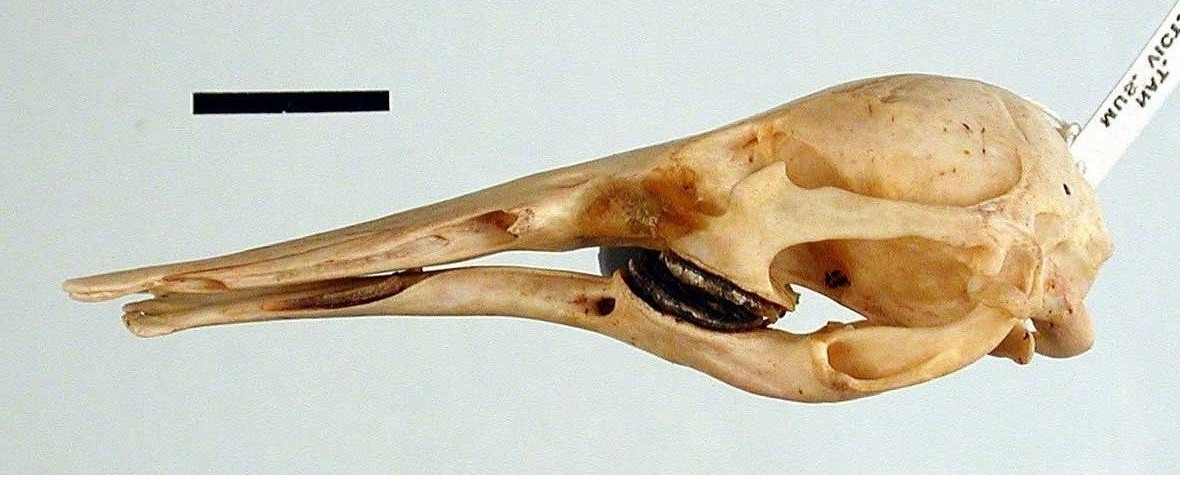
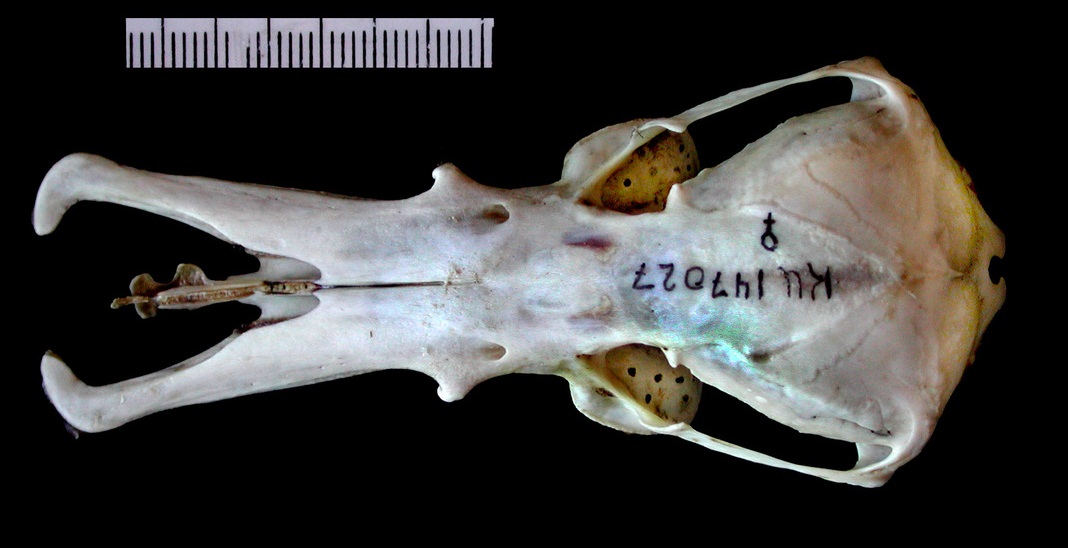
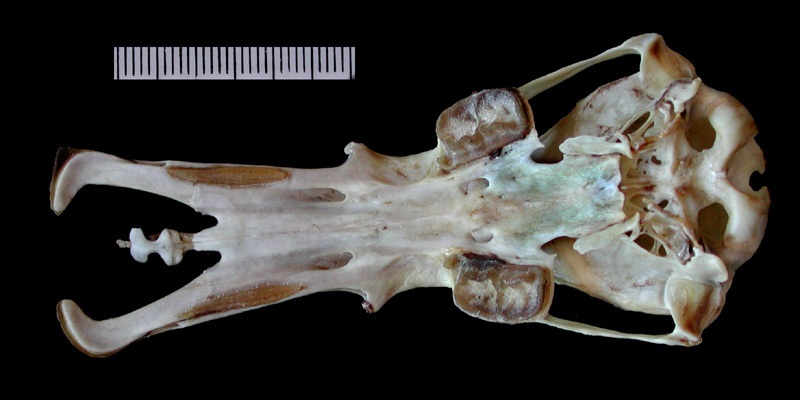
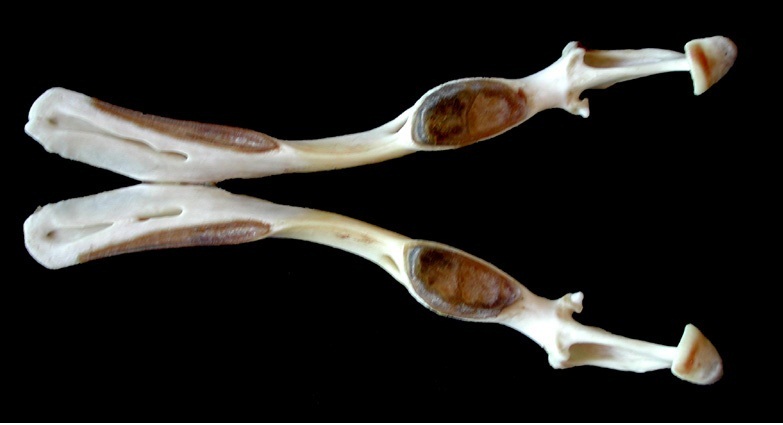

Череп и нижняя челюсть утконоса (Ornithorhynchus anatinus).
Сверху и по бокам верхней челюсти имеются отверстия для прохождения ветвей тройничного нерва. Небо очень длинное и тоже пронизано большими отверстиями для ветвей тройничного нерва, а равно и основание черепа. Носовые ходы очень длинные. Плоские, продырявленные зубные ложа, образованные расширениями верхнечелюстных костей, представляют основания для щечных зубов и заменяющих их роговых пластинок. Маленькие глаза расположены высоко на черепе — особенность, характерная для животных, плавающих у водной поверхности. Скуловые дуги тонкие и расположены близко к черепу, соответствуя слабой жевательной мускулатуре. Все же, дуги гораздо толще и массивнее, чем у ехидн. Основание скулового отростка чешуйчатой кости пронизано широким височным каналом для затылочной артерии. Мозговая коробка большая и округлая, хотя пропорционально и меньше, чем у ехидн.
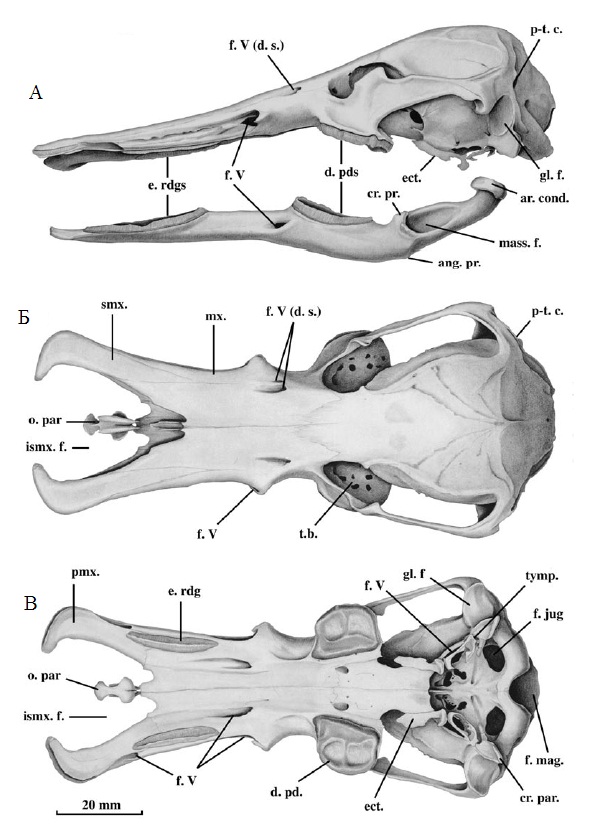
Прорисовки черепа и нижней челюсти утконоса (Ornithorhynchus anatinus); сбоку (А), сверху (Б) и снизу (В): ang. pr. — рудиментарный угловой отросток; ar. cond. — суставной мыщелок; cr. par. — гребень на каменистой части височной кости; cr. pr. — венечный отросток; d. pd.(s) — роговые пластинки; ect. — эктоптеригоид; e. rdg (s). — роговые гребни; f. jug. — яремное отверстие; f. mag. — большое затылочное отверстие; f. V — отверстия для прохождения ветвей V-го черепного нерва, иннервирующих клюв; f. V (d. s.) — отверстия для прохождения ветвей V-го черепного нерва, иннервирующих лобный щиток; gl. f. — сочленовная ямка; ismx. f. — межсептомаксиллярное окно; mass. f. — массетерная ямка; mx. — верхнечелюстная кость; o. par. — гантелевидная кость; pmx. — межчелюстная кость; p-t. c. — задневисочный канал; smx. — септомаксиллярная кость; t.b. — зубное ложе; tymp. — барабанная кость.
Барабанная полость снизу полностью открыта, костный слуховой барабан не образуется. Тонкие, пластинчатые эктоптеригоиды, прикрепленные к небным костям, упираются в кольцеобразные барабанные кости. По сравнению с ехиднами, элементы среднего уха не так неподвижны: барабанная кость, наковальня и молоточек соединены плотно, но без срастания, так что легко могут быть отделены друг от друга. Молоточек и наковальня значительно меньше (хотя молоточек больше наковальни примерно в 3 раза) и изящнее, чем у ехидн. Само среднее ухо не плотно присоединено к костям черепа, а занимает как бы плавающее положение. Возможно, у утконоса наблюдается внутривидовая изменчивость в появлении стременного отверстия в эмбриональном развитии, так как иногда оно упоминается, а в других случаях указывается, что стременное отверстие отсутствует на протяжении всего развития. В любом случае, у взрослого животного стременного отверстия нет. Утконос, по-видимому, единственное современное млекопитающее, сочетающее отсутствие стременного отверстия с развитой стременной артерией во взрослом состоянии, причем артерия проходит позади стремени.
Хрящевой наружный слуховой проход поднимается, изгибаясь, с нижней стороны черепа вверх до глазницы, где крепится к верхней части скуловой дуги позади глаза. К нижней челюсти проход прикрепляется в месте расположения барабанной перепонки. Благодаря такому строению слухового прохода утконос может улавливать передаваемые по воздуху звуковые колебания, находясь в это время в полупогруженном состоянии под водной поверхностью (примечательно, что сходным образом по сравнению с предковым изменено ухо современных крокодилов).
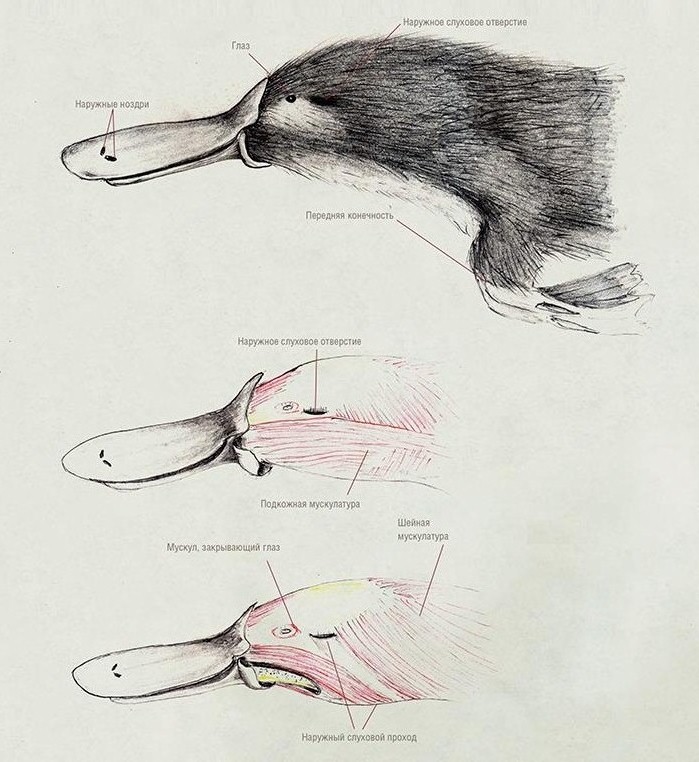
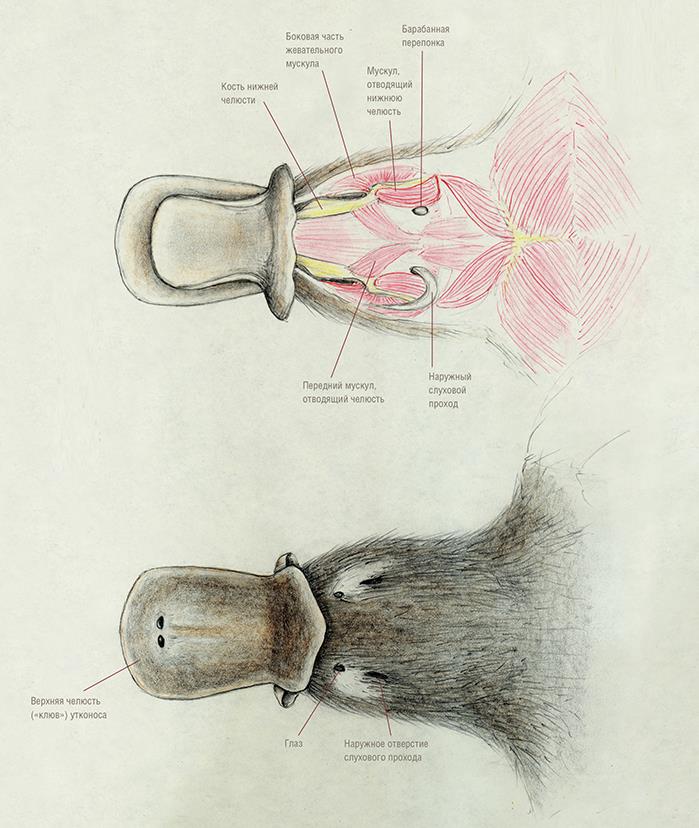
Мускулатура головы утконоса (Ornithorhynchus anatinus).
Как и в верхней челюсти, удлиненные зубные кости впереди длинного симфизного сращения значительно расходятся в стороны; между ними залегает хрящевая пластинка. Суставной отросток довольно хорошо развит, тогда как венечный отросток утрачен, у угловой отросток может иметься или отсутствовать у различных особей. Массетерная ямка преобразована в глубокий карман.
Нижнюю челюсть опускает парная m. detrahens mandibulae, с каждой стороны прикрепляющаяся к нижней части ее внутренней поверхности. В то же время, у териев за это действие отвечает парная двубрюшная мышца (m. digastricus), которая состоит из 2 мышечных брюшек, соединенных общим сухожилием. M. detrahens mandibulae является производным жевательной мышцы (m. masseter), сместившейся назад к челюстному суставу и взявшей на себя функцию открывания рта. M. detrahens mandibulae и переднее брюшко m. digastricus иннервируется ветвью тройничного (V-го черепного) нерва, тогда как заднее брюшко — лицевым (VII-м черепным) нервом.
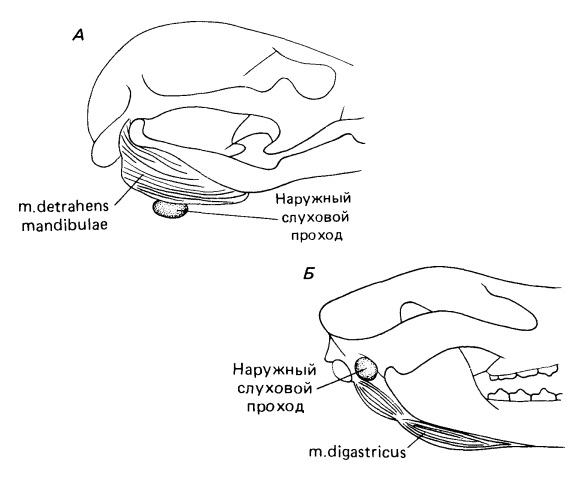
Наружный слуховой проход и мышцы, опускающие нижнюю челюсть: А — утконос (Ornithorhynchus anatinus), наружный слуховой проход находится под m. detrahens mandibulae; Б — виргинский опоссум (Didelphis virginiana), слуховой проход находится над m. digastricus.
Зубы присутствуют лишь у молодых особей, до достижения ими приблизительно 1/3 веса взрослых. Они закладываются в следующем числе: I0/i5, С1/с1, Р2/р2, М3/m3 = 34. Однако в итоге у детенышей прорезываются только II-й предкоренной и I-й коренной сверху и с I-го по III-й коренные снизу, так что зубная формула составляет I0/i0, С0/с0, Р1/р0, М1/m3 = 10. Не совсем понятно, какое поколение представляют зубы утконоса — молочное или постоянное. Настоящие коренные по определению должны быть постоянными, не иметь предшественников и не выпадать. С другой стороны, даже у прорезывающихся предкоренных и остальных непрорезывающихся зубов утконоса не обнаружено предшественников. Возможно, его зубы все же гомологичны постоянным зубам вымерших однопроходных и прочих млекопитающих. Другой допустимый вариант — все зубы до коренных молочные, а коренные постоянные.
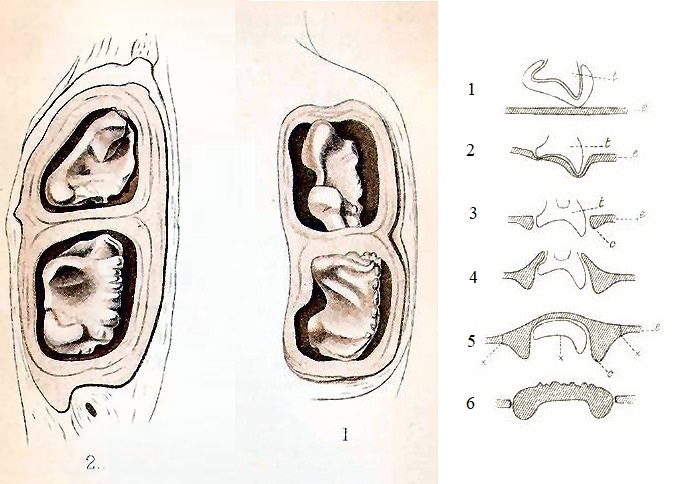
Слева — прорисовки щечных зубов утконоса (Ornithorhynchus anatinus), справа — схема их прорезания и замены роговыми пластинками: 1 — зуб, покрытый эмалевым органом и ротовой эпителий под ним; 2 — зуб перед прорезанием; 3 — прорезавшийся зуб; 4 — края эпителия ороговевают; 5 — образование роговой пластинки, в которой лежит зуб; 6 — зуб исчезает, пластинка отделяется от окружающей ткани.
Прорезавшиеся зубы мелкие по размерам и не полностью заполняют зубные ложа; их строение существенно варьирует у разных особей. Зубы кальцинированы, покрыты тонким слоем эмали и имеют многочисленные короткие корни — по меньшей мере, по 2 с наружной и внутренней стороны каждого зуба. Выступающие из десен коронки напоминают по своей форме плоские, неправильно округленные блюдечки. P1 и m3 небольшие по размерам и имеют по 1 основному широкому и низкому бугорку. На остальных зубах бугорки на язычной и губной сторонах жевательной поверхности соединены друг с другом поперечными гребнями-лофами, образуя двойные лопасти. По периметру каждый зуб усажен несколькими более мелкими бугорками. Отличие от ископаемых обдуродонов (Obdurodon) состоит в сильнее развитых бугорках, более наклонно ориентированных лофах, лучше выраженных цингулюмах и утратой цингулюма на М1.
Зубы у утконоса очень непрочные и после перехода к самостоятельному питанию быстро стираются. Вслед за этим рассасываются их отделы, расположенные в челюстях. Позднее зубные ложа покрываются сверху твердыми роговыми пластинками, берущими начало из эпителия ротовой полости. На полпути между зубными ложами и концом клюва находится еще один набор пластинок, заменяющий отсутствующие резцы и клыки. Они имеют вид длинных, узких и острых гребней и используются для удержания и измельчения пищи, тогда как широкие и плоские задние пластинки действуют в качестве раздавливающих и перетирающих поверхностей. Пластинки в значительной мере повторяют строение исчезнувших зубов: на месте верхних коренных они разделены на 2 доли, а на месте нижних в дополнение к 2 долям имеют маленькую заднюю дольку, соответствующую колышковидному m3. Подобная структура отчасти аналогична дифференцированной зубной системе большинства млекопитающих. Роговые пластинки постоянно стираются от грубой пищи и вновь обновляются до самого конца жизни.
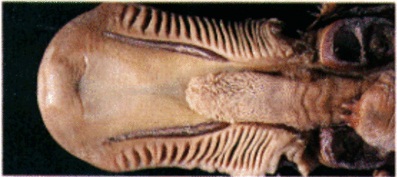
Нижняя половина клюва и язык утконоса (Ornithorhynchus anatinus).
Вдоль боковых краев нижней челюсти расположено примерно по 20 поперечных гибких роговых зазубрин, которые используются для отцеживания несъедобных частиц пищи. Язык короткий и мясистый. Сверху на его передней части расположены 2 продольных роговых гребня, помогающих захватывать добычу и заменяющих в данном отношении отсутствующие резцы и клыки. На язычном основании находится круглая площадка, у переднего конца которой расположены 2 роговых шипа, трущихся о небо и помогающих измельчать пищу. Задняя часть языка утолщается в виде своеобразного желвака, который совершенно замыкает ротовую полость. Позади клюва в нее открываются 2 объемистых защечных мешка. Имеются небольшие слюнные железы.

Грудная клетка и прорисовка плечевого пояса утконоса (Ornithorhynchus anatinus), снизу.
Для пищеварительного тракта характерно простое строение и сравнительно небольшая длина. У самки длиной 44 см его протяженность составила 140 см. Небольшой тонкостенный желудок лишен пищеварительных желез и служит для накопления тщательно измельченного в ротовой полости корма. На стенках короткой тонкой кишки отсутствуют ворсинки, но имеется множество поверхностных складок. Группы пищеварительных желез открываются в общие канальцы, устья которых расположены между этими складками. В переднем отделе тонкой кишки имеется скопление бруннеровых желез. Толстая кишка также короткая, с небольшой слепой кишкой. Прямая кишка по своему диаметру превосходит остальную часть кишечника. О процессах пищеварения практически ничего неизвестно, хотя характер поедаемой пищи предполагает высокую протеолитическую активность секретов поджелудочной железы и желез стенок кишечника.
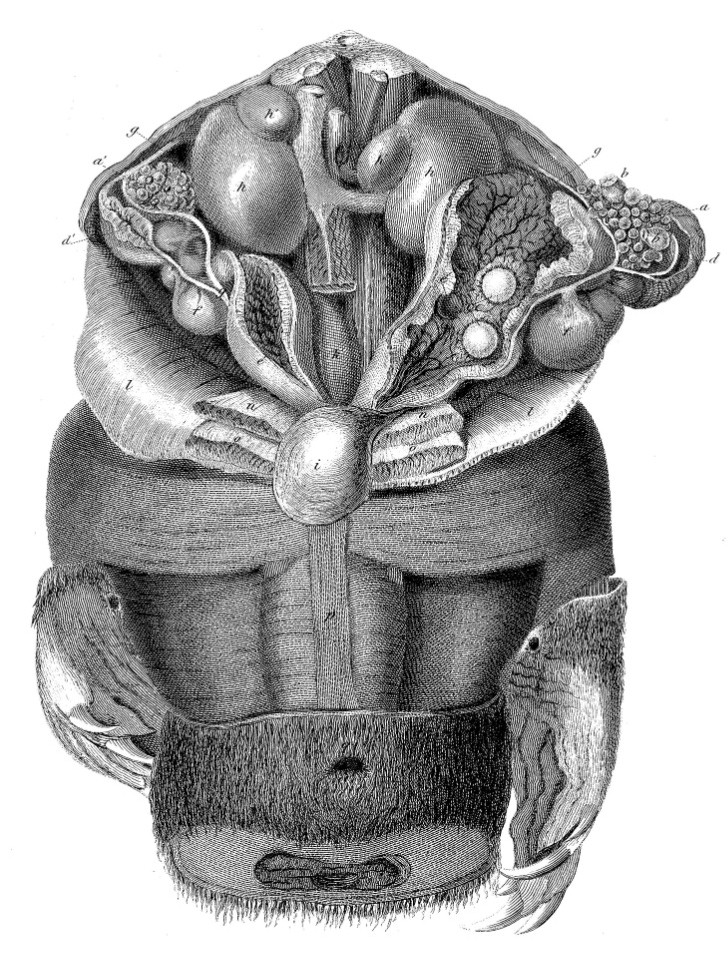
Старинный рисунок органов брюшной полости утконоса (Ornithorhynchus anatinus).
Строение и функционирования почек изучено плохо. Водный обмен весьма высок (480 мл/день), что вполне ожидаемо для кормящегося в водоемах животного.
Подобно некоторым рептилиям и многим птицам, у самок функционирует только один яичник — левый. Пенис самца расположен внутри клоакальной камеры в тонкостенном препуциальном мешке. В состоянии эрекции он выходит из отверстия клоаки наружу. Длина пениса составляет 5-7 см, он заканчивается головкой с 2 лопастями. Левая лопасть развита гораздо больше, что объясняется нефункциональностью правого яичника самки. При эякуляции сперма выбрасывается через левую лопасть, тогда как правая сомкнута.
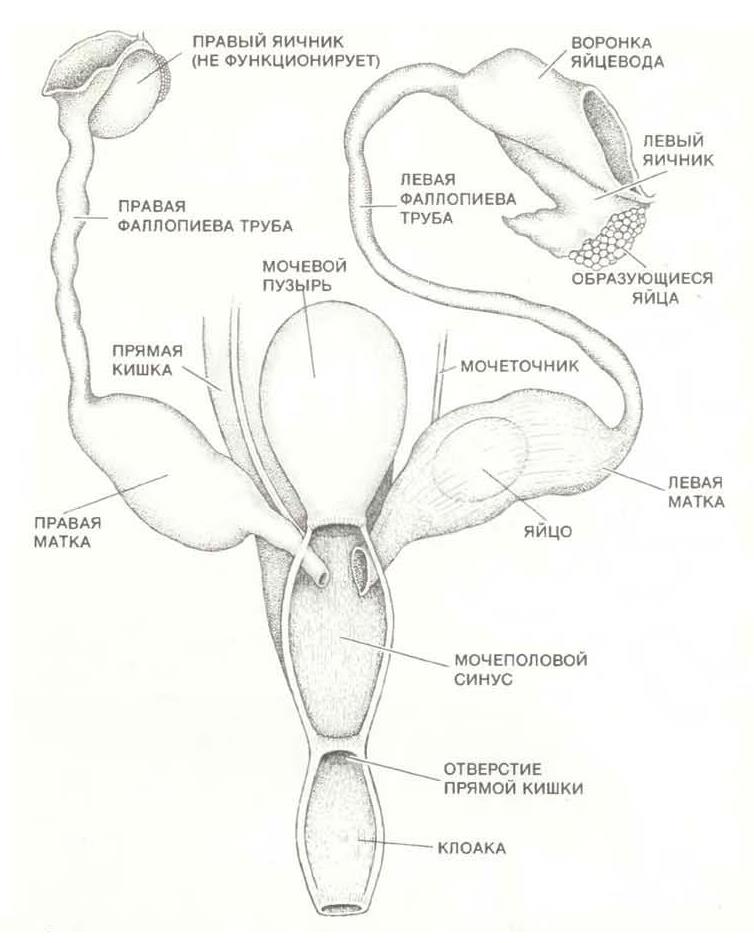
Репродуктивная система самки утконоса (Ornithorhynchus anatinus).
Головной мозг утконоса имеет архаичное строение. Обонятельные луковицы относительно небольшие и составляют 0,8 % объема головного мозга. Новая кора занимает порядка 48 % поверхности мозга, но большие полушария гладкие (лиссэнцефалия) и лишь по их сторонам имеются хорошо выраженные сильвиевы борозды. Корковые зоны для приема и обработки соматосенсорных, зрительных и слуховых импульсов сгруппированы в задней части и по бокам полушарий, проникая также в лобную область. Древняя кора, или обонятельная доля, относительно меньше, чем у ехидн. Мозжечок, напротив, сравнительно больше.
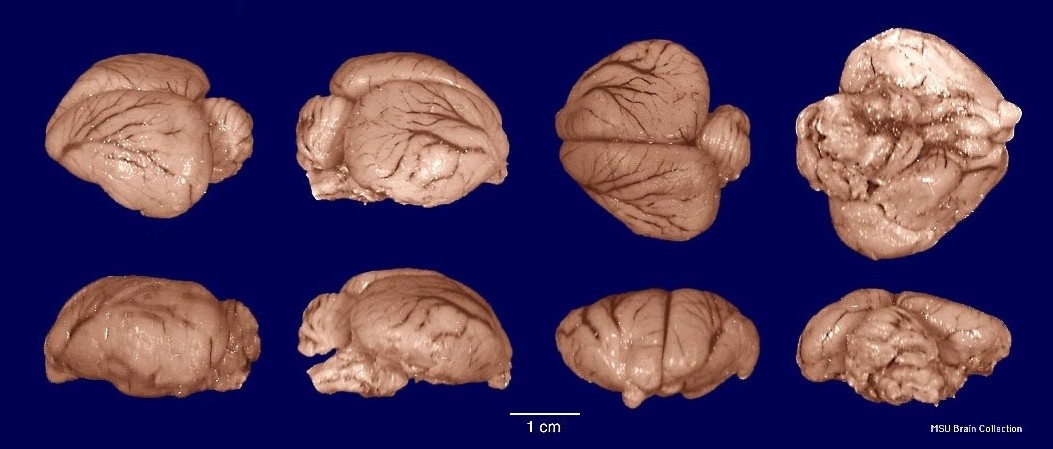
Головной мозг утконоса (Ornithorhynchus anatinus).
Мозг спящего утконоса последовательно проходит фазы медленного и быстрого сна. Внешне фаза быстрого сна проявляется подергиванием век, дрожанием головы и шеи. Также спящее животное может совершать передними конечностями плавательные движения.
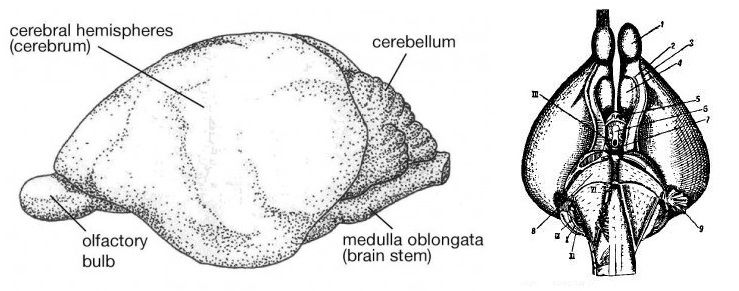
Прорисовки головного мозга утконоса (Ornithorhynchus anatinus), сбоку и снизу: 1 — обонятельная луковица (bulbus olfactorius); 2 — грушевидная доля; 3— обонятельная бороздка (fovea rhinalis); 4 — обонятельный бугорок (tuberculum olfactorium); 5 — конечная пластинка (lamina terminalis); 6 — серый бугор (tuberculum cinereum); 7 — сосковидные тела (corpora mammilaria); 8 — мост; 9 — придатки (flocculi); I-XII — черепно-мозговые нервы.