в оглавление
назад
Зубы (продолжение)

Череп с нижней челюстью сумчатого льва (Thylacoleo carnifex).
Итак, в зависимости от образа жизни и характера питания зубная система млекопитающих претерпела значительные адаптивные изменения. Исходный сплошной ряд из сравнительно простых, слабо дифференцированных зубов в известной мере сохранился лишь у насекомоядных (в широком смысле), рукокрылых и еще некоторых групп, да и то с определенными оговорками в каждом отдельном случае. Например, у землероек (Soricidae), отличающихся вытянутой узкой мордой, зубные ряды с выдвинутыми вперед резцами образовали своеобразный «пинцет», которым удобно захватывать и удерживать мелкую добычу (насекомых, червей и др.), а бугорчатыми с острыми вершинами коренными зубами дробить их хитиновый покров. Мало дифференцированы зубы у питающихся преимущественно мягкими плодами обезьян: их клыки не велики, а коренные имеют бугорчатые или плоские жевательные поверхности.
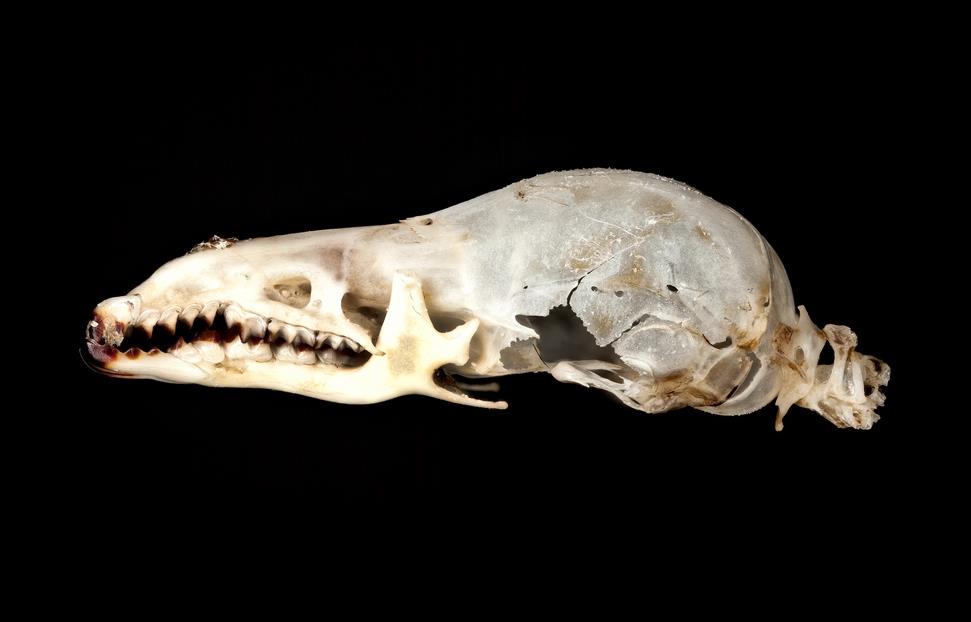
Череп с нижней челюстью и передняя часть шейного отдела позвоночника масковой бурозубки (Sorex cinereus).
Большую дифференцировку испытал зубной ряд плотоядных млекопитающих. Их резцы очень острые, а клыки значительно увеличены. Иногда верхние клыки достигают просто громадных размеров — развивается т. н. саблезубость. Это характерно для тилакосмилов (Thylacosmilidae), некоторых арктоционов (Arctocyonidae), махероидов (Machaeroidinae) и особенно для кошкообразных (Feliformia) — нимравов (Nimravidae), барбурофелисов (Barbourofelidae), махайродонтов (Machairodontinae). Очень своеобразны сумчатые львы (Thylacoleonidae), у которых орудием нападения служили не клыки (из которых сохранились только сильно уменьшившиеся верхние), а сильно увеличенные резцы.
Зубной ряд тюленя-крабоеда (Lobodon carcinophagus).
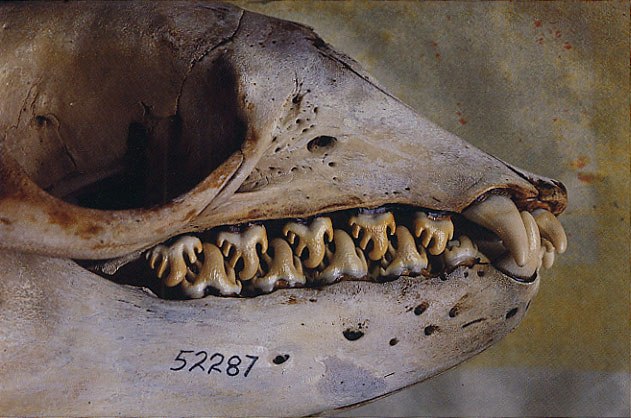
Относительно очень большие плоские щечные зубы свойственны калану (Enhydra lutris), пищу которого составляют морские ежи, моллюски и ракообразные — животные с твердыми скелетными образованиями. Очень массивны зубы гиен (Hyaenidae), дробящих трубчатые кости крупных животных. У ластоногих (Pinnipedia) в связи с заглатыванием неразмельченной пищи произошло вторичное упрощение зубов, однако клыки у многих из них хорошо развиты. Так, у моржа (Odobenus rosmarus) клыки преобразованы в мощные, постоянно растущие бивни, достигающие у матерых самцов 82 см в длину. Они выполняют множество функций: используются при добывании корма на морском дне, помогают выбираться из воды на лед, служат для защиты от белых медведей, но в первую очередь служат для внутривидовых демонстраций и являются турнирным оружием. Коренные зубы у моржа утрачены, а предкоренные преобразованные в «жернова» для дробления и перетирания раковин моллюсков. У тюленя-крабоеда (Lobodon carcinophagus) фестончато-бугорчатые щечные зубы, которые при смыкании плотно прилегают друг к другу и образуют своеобразное сито для фильтрации мелкого антарктического криля.
Череп с нижней челюстью обыкновенного вампира (Desmodus rotundus).
Далеко зашедшая специализация — питание кровью птиц и млекопитающих — имеет место у летучих мышей-вампиров (Desmodontinae). Их увеличенные резцы выдаются вперед и имеют очень острые режущие края. У шерстокрылов (Cynocephalidae) верхние резцы располагаются по бокам челюсти, а нижние направлены вперед и прорезаны глубокими узкими щелями, делающими эти зубы похожими на гребешок. Полагают, что нижние резцы служат шерстокрылам для ухода за шерстью. Сходным образом используют свои узкие нижние резцы и многие лемуры.

Нижняя челюсть шерстокрыла.
У грызунов число резцов сокращается до 2 в верхней и нижней челюсти, клыки исчезают, предкоренные также частично или полностью утрачиваются, а коренные (вместе с сохранившимися предкоренными) приобретают бугорчатую, гребенчатую или складчатую жевательную поверхность с перегородками (валиками) из эмали. Такое строение зубной системы связано с характером питания: резцы сгрызают или скоблят растительность, а коренные, подобно жерновам, перетирают даже самые грубые ее части. Резцы приобретают форму стамесок и растут своей корневой частью в течение всей жизни, так что их стирание на концах непрерывно возмещается нарастанием у основания. Например, у серой крысы (Rattus norvegicus) скорость роста резцов составляет примерно 3 см в месяц. При повреждении одного из резцов противостоящий ему резец, не встречая препятствий, вырастает у грызунов до такой степени, что иногда образует уродливый завиток или кольцо, мешающее питанию животного, что может привести к гибели.

Слева: зубная система восточного гофера (Geomys sp.). Зубы растут всю жизнь, чтобы компенсировать активное снашивание и стирание во время активного измельчения растительного материала.
Справа: ненормальный рост верхнего резца сурка-байбака (Marmota bobak), как следствие повреждения противостоящего ему и в норме стирающего его нижнего резца.
Благодаря тому, что эмаль имеется только на передней поверхности постоянно растущих резцов грызунов (а также зайцеобразных, вомбатов (Vombatidae) и ряда других млекопитающих), их снашивание идет неравномерно, поскольку задняя поверхность стирается быстрее передней. Благодаря этому обеспечивается постоянная острота резцов. У домовой мыши (Mus musculus) на задней поверхности верхних резцов имеется тонкий слой эмали, задерживающий стирание. Поэтому, в отличие от других мышей (лесной (Apodemus uralensis), полевой (Apodemus agrarius)), на этих резцах у нее образуется характерная выемка. Это явление рассматривается как приспособление к питанию мелкими семенами: в выемке удерживается мелкое семечко, обрабатываемое нижними резцами. Дикобразы (Hystrix) дополнительно затачивают зубы о камни.
Череп с нижней челюстью калимантанского орангутана (Pongo pygmaeus).
У ряда грызунов, например, у тушканчиков (Dipodidae), землекопов (Bathyergidae), слепышей (Spalacinae), слепушонок (Ellobius) и др. резцы играют важную роль при разрыхлении плотной почвы во время прокладывания ходов. С этим связаны крупные размеры резцов и их положение: они сильно выступают вперед, а у некоторых видов (слепышей и др.) находятся вне рта, так как на губах имеются специальные выступы, которые смыкаются позади резцов и ограничивают ротовое отверстие спереди. Благодаря этой особенности земля, разрыхляемая резцами, не попадает в рот зверька. Поддержание постоянного роста резца требует большого размера его альвеолярной части. Иногда этот объем увеличивается настолько, что резец простирается через всю нижнюю челюсть и выдается на наружной поверхности в ее задней части вблизи основания сочленовного отростка, образуя альвеолярный бугор. У форм, роющих с помощью резцов (слепыши, слепушонки), стирающая нагрузка на резец особенно сильна, поэтому объем его альвеолы исключительно велик, и альвеолярный бугор даже может выступать в виде довольно длинного отростка. Между резцами и коренными зубами у грызунов образуется широкая диастема. Коренные зубы хорошо развиты, хотя их количество имеет тенденцию к сокращению. У видов, питающихся грубыми частями растений, коренные не имеют корней и, подобно резцам, непрерывно растут по мере снашивания коронки от перетирания пищи.
Зайцеобразные позади пары нормальных резцов в верхней челюсти имеют пару недоразвитых, принимающих на себя удары нижних резцов при скусывании твердых стеблей. Подобно некоторым землероям из грызунов, у арктического беляка (Lepus arcticus), вынужденного прогрызать при добывании корма корку плотного снега, резцы сильно выдаются вперед.
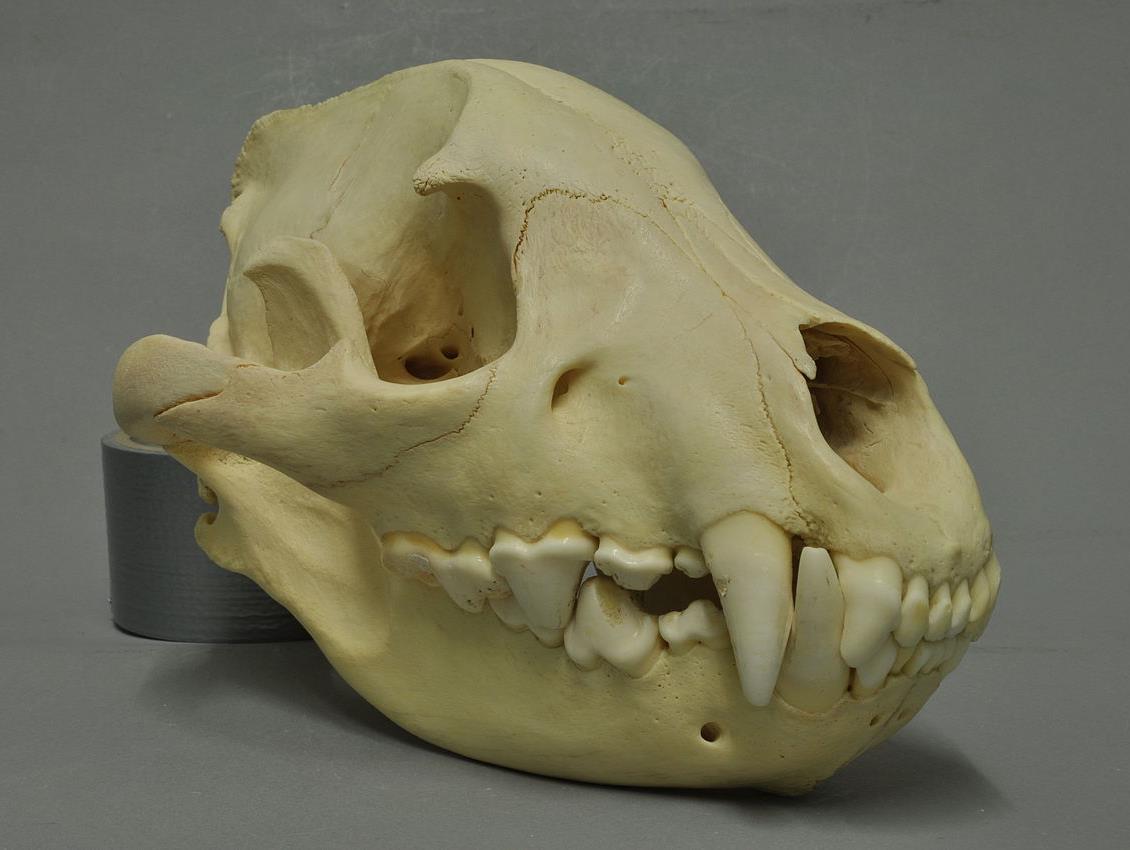
Череп с нижней челюстью пятнистой гиены (Crocuta crocuta).
В зубной системе копытных первенствующее значение принадлежит щечным зубам, резцы же и клыки в значительной степени редуцируются. Сильное развитие верхних и нижних клыков у свиней (Suidae) в первую очередь связано с добыванием корма из-под земли. У бородавочников (Phacochoerus) верхние клыки могут достигать 60 см в длину; с их помощью эти животные даже роют норы. Важна также оборонительная функция клыков. Они больше у самцов и помимо прочего используются ими в качестве турнирного оружия. Этой же цели служат увеличенные верхние клыки самцов оленьков (Tragulidae), кабарог (Moschidae) и некоторых оленей (Cervidae). Предкоренные многих копытных моляризуются и вместе с коренными отделяются от остальных зубов диастемой. Копытные, питающиеся жесткими степными травами (например, те же лошади (Equidae)), обладают щечными зубами с высокими коронками прямоугольной или цилиндрической формы. Наоборот, копытные, поедающие мягкую сочную растительность (например, бегемоты (Hippopotamidae)), имеют низкокоронковые щечные зубы.

Череп с нижней челюстью гиппопотама (Hippopotamus amphibius).
Очень своеобразна зубная система слонов (Elephantidae). Из резцов у современных их форм сохраняются лишь верхние крайние, которые превратились в огромные (до 3,5 м длиной) постоянно растущие бивни, служащие для выкапывания корней, сдирания коры, служащие турнирным оружием, защитой от хищников и пр. Кроме бивней, имеется только четыре коренных зуба — по 1 в каждой половине верхней и нижней челюсти. Они исключительно массивны, а их высокие коронки обладают набором поперечно стоящих дентиновых пластин, облицованных эмалью и чередующихся с прослойками цемента. Такие коренные легко справляются даже относительно толстыми ветвями деревьев. Помимо слонов, увеличение одной верхней пары резцов и редукция прочих их пар наблюдалась у продвинутых мастодонтов (Mammutidae) и гомфотериев (Gomphotheriidae). Наоборот, у дейнотериев (Deinotheriidae) разрасталась одна из нижних пар резцов. У сирен резцы либо редуцируются (ламантины (Trichechidae)), либо превращаются в бивни (дюгони (Dugongidae)), клыки и предкоренные редуцируются, однако общее число коренных возрастает.

Череп с нижней челюстью дюгоня (Dugong dugon).
В результате упрощения и вторичных преобразований зубная система некоторых млекопитающих стала гомодонтной. У броненосцев (Cingulata) и ленивцев (Folivora) резцы и клыки отсутствуют, а все щечные в виде колышков, без эмали и с открытыми корнями. У зубатых китов (Odontoceti) зубы также имеют простую коническую форму. Как будет показано далее, число гомодонтных зубов может быть очень большим.

Череп с нижней челюстью девятипоясного броненосца (Dasypus novemcinctus).
Интересную особенность имеют зубы нарвала (Monodon monoceros). У его самцов (гораздо реже — у самок) из всех зубов прорезывается только левый клык, который образует огромный направленный вперед бивень длиной до 3 м. Назначение бивня до сих пор точно не прояснено, но, насколько известно, он не служит орудием нападения и не используется для пробивания корки льда при устройстве продухов. Предполагалось, что он необходим при брачных играх, для привлечения самок. Существует также версия, что бивни используются самцами во время брачных турниров, — было замечено, что они иногда трутся бивнями. Данная точка зрения подтверждается тем, что в тканях головы самцов иногда находят обломанные фрагменты бивней их вероятных соперников. Наконец, высказано мнение, что бивень чувствителен к изменениям давления, температуры и относительной концентрации взвешенных частиц в воде. Бивень лишен эмали, его внешний слой образует пористый цемент, а расположенный внутри дентин пронизан микроскопическими канальцами. В дентине залегает пульпа, где собрано множество нервных окончаний. Предполагается, что через пористую структуру зуба они получают информацию об окружающей среде, что может способствовать успеху в поиске пищи и обнаружении готовых к спариванию самок. Действительно, исследователями была обнаружена связь между различным содержанием соли в воде и соответствующими изменениями частоты сердечных сокращений нарвала. В ходе экспериментов пресная и соленая вода по очереди контактировали лишь с внешней поверхностью бивня. Все же, функциональность бивня как сенсорного органа (если она действительно имеет место) скорее всего, является вторичной.

Смонтированный скелет самца нарвала (Monodon monoceros).
Для большинства рептилий, зверообразных и млекопитающеобразных примитивнее синоконодона (Sinoconodon rigneyi) характерна многократная смена зубов, или полифиодонтия (хотя у ряда зверообразных уже имелись постоянные зубные группы, а у синоконодона темпы замещения зубов снизились). У ранних настоящих млекопитающих процесс замещения зубов редуцировался до однократной их смены — развилась дифиодонтия. В дифиодонтной зубной системе сначала появляются молочные зубы (dentes decidui), которые лишены корней и держатся шейкой за десну и края альвеолы. Развивающиеся под ними более крупные постоянные, или дефинитивные, зубы (dentes permamentes) обладают корнями, покрыты слоем зубного цемента и соединяются со стенками альвеол. По мере роста постоянный зуб выталкивает молочный, т. е. смена зубов происходит в вертикальном направлении. Обычно лишь часть постоянных зубов имеет своих молочных предшественников. Как правило, это резцы, клыки и предкоренные. Коренные прорезаются сразу в постоянной форме.
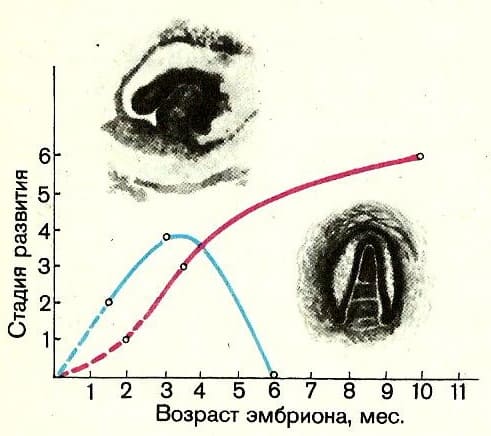
У современных взрослых усатых китов (Mysticeti) зубов нет, но они находятся в толще челюстей эмбрионов на начальных стадиях развития и затем, к 6 месяцам, бесследно рассасываются. У зубатых китов (Odontoceti) зубы сохраняются всю жизнь, хотя развиваться они начинают несколько позже, чем у усатых китов.
На первый взгляд может показаться, что зубная система млекопитающих состоит из набора молочных выпадающих зубов, в которых не входят коренные, и полного постоянного набора. Однако при более внимательном рассмотрении развития зубов создается другое представление. Молочные зубы типичных млекопитающих (в число которых входит и человек) обычно развиваются последовательно один за другим спереди назад — от резцов до молочных предкоренных, или «молочных коренных» (клыки с длинными корнями могут прорезаться с задержкой). После появления последнего «молочного коренного» начинается последовательное прорезание 3 настоящих коренных зубов. Тем временем еще до окончания этой первой волны передние зубы по очереди заменяются вплоть до предкоренных включительно. Но этот процесс дальше не продолжается, и коренные не заменяются. Отсюда ясно, что имеется один полный набор и второй неполный; коренные, хотя они и постоянные, принадлежат к тому же поколению, что и молочные зубы. Вот почему I-й коренной из постоянных зубов прорезается у человека первым. Таким образом, у взрослых млекопитающих во рту имеются части 2 зубных рядов; коренные принадлежат к первому ряду, а более передние зубы — ко второму. Это представление объясняет различные необычные факты вроде того, что коренные во многих случаях имеют значительно большее сходство с недолговечными «молочными коренными», чем с постоянными предкоренными, с которыми они соседствуют в течение большей части жизни.
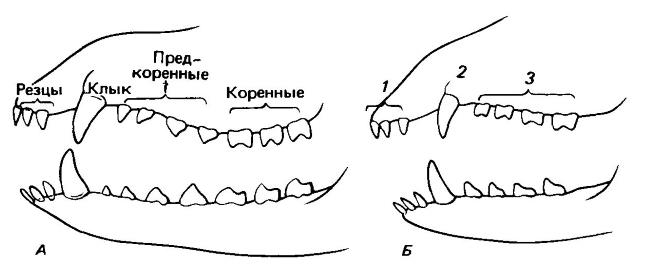
Схема, показывающая состав молочного и постоянного набора зубов на примере генерализованного плацентарного млекопитающего: А — постоянные зубы; Б — выпадающие зубы; 1 — выпадающие резцы; 2 — выпадающий клык; 3 — «молочные коренные» (выпадающие предкоренные).
Замена зубов — весьма целесообразное приспособление. Молодое животное уже рано, до полного вырастания, должно питаться самостоятельно и нуждается в определенном зубном наборе. Если бы прорезающиеся у него зубы сразу достигали полной величины, как у взрослого млекопитающего, они были бы чрезмерно велики. Напротив, зубы, пригодные по величине для молодого животного, не могли бы обеспечивать питание животного, выросшего в два и более раза, так как зубы в отличие от костей не могут перестраиваться и сохраняют первоначальную величину и форму. Замена мелких зубов крупными, а в дальнейшем — прорезание позади них коренных, удлиняющих зубной ряд, дает возможность нормально питаться как молодому, так и взрослому животному.

Постоянные и молочные (заштрихованы) зубы кошки (Felis silvestris): i — резцы (incisivi); с — клыки (canini); p — предкоренные (praemolares); m — коренные (molares).
При упрощении функции те или иные зубы могут утрачивать одно поколение и сохраняться на всю жизнь в той форме, в какой первоначально прорезались. Так, у основной массы сумчатых замене подвержены лишь III-и предкоренные — т. е., наблюдается неполная монофиодонтия. У некоторых сумчатых также выявлены нефункциональные, не прорезывающиеся зачатки молочного поколения резцов и клыков, хотя у других ничего подобного не обнаружено. Из-за полного отсутствия следов молочного поколения зубов у некоторых и отсутствия предшественников I-х и II-х коренных у всех сумчатых не совсем понятно, какие именно зубы функционируют у них в качестве постоянных: молочные или настоящие постоянные. Любопытно, что сменяемые III-и предкоренные у сумчатых развиваются и прорезываются раньше остальных зубов (у плацентарных наблюдается почти обратная ситуация — у них, за редким исключением, не меняются I-е предкоренные).

Спил челюстей человеческого ребенка перед потерей молочных зубов.
У землероек (Soricidae), многих кротов (Talpidae), некоторых рукокрылых, ластоногих, а из выражено гетеродонтных млекопитающих у полосатых скунсов (Mephitis) прорезывается и работает только постоянный набор зубов, а молочные зубы зачаточные и нефункциональные. Таким образом, их зубная система монофиодонтная. Эта особенность вообще характерна для огромного большинства млекопитающих с гомодонтными зубами, за исключением броненосцев. У последних (а также у ленивцев и трубкозубов) происходит ускоренная компенсация износа постоянного поколения дифиодонтных зубов, поскольку они однослойны (эмаль утрачена) и процесс прироста ограничен лишь откладкой дентина.

Возрастные изменения зубов у домашней лошади (Equus ferus caballus). Цифры сбоку указывают возраст животного.
Набор зубов строго специфичен и высококонсервативен, в связи с чем является важным компонентом характеристики таксонов. Для компактного его описания употребляют т. н. зубные формулы, в которых указывают число зубов одной половины верхней и нижней челюстей. Типы зубов сокращенно обозначаются начальными буквами их латинского названия: резцы — i, клыки — с; предкоренные — р; настоящие коренные — m. Зубы верхней челюсти обозначаются большой буквой, нижней — маленькой. Для обозначения молочных зубов прибавляется буква d. Приведем примеры зубных формул:
виргинский опоссум (Didelphis virginiana) — I3/i4, C1/c1, P3/p3, M4/m4 = 50;
волк (Canis lupus) — I3/i3, C1/c1, P4/p4, M2/m3 = 42;
енот-полоскун (Procyon lotor) — I3/i3, C1/c1, P4/p4, M2/m2 = 40;
лесная куница (Martes martes) — I3/i3, C1/c1, P4/p4, M1/m2 = 34;
калан (Enhydra lutris) — I3/i2, C1/c1, P3/p3, M1/m3 = 32;
дикая кошка (Felis silvestris) — I3/i3, C1/c1, P3/p2, M1/m1 = 30;
благородный олень (Cervus elaphus) — I0/i3, C0-1/c1, P3/p3, M3/m3 = 32-34;
тур (Bos primigenius) — I0/i3, C0/c1, P3/p3, M3/m3 = 32;
дикая лошадь (Equus ferus) — I3/i3, C1/c1, P3/p3, M3/m3 = 40;
ночной крылан (Alionycteris paucidentata) — I1/12, C1/c1, P3/p3, M1/m2 = 26;
человек разумный (Homo sapiens) — I2/i2, C1/c1, P2/p2, M2-4/m2-4 = 28-36;
дикий кролик (Oryctolagus cuniculus) — I2/i1, C0/c0, P3/p2, M2-3/m3 = 26-28;
обыкновенный бобр (Castor fiber) — I1/i1, C0/c0, P1/p1, M3/m3 = 20;
обыкновенная белка (Sciurus vulgaris) — I1/i1, C0/c0, P2/p1, M3/m3 = 22;
домовая мышь (Mus musculus) — I1/i1, C0/c0, P0/p0, M3/m3 = 16;
бобровая крыса (Hydromys chrysogaster) — I1/i1, C0/c0, P0/p0, M2/m2 = 12;
саванный слон (Loxodonta africana) — I1/i0, C0/c0, P3/p3, M3/m3 = 26 (на протяжении жизни, одновременно обычно действует 6).
Как можно видеть, количество зубов в разных случаях сильно варьирует. У большинства ранних форм общее число зубов превышает таковое у более поздних. Так, у млекопитающеобразного синоконодона (Sinoconodon rigneyi) зубная формула составляет I5/i5, C1/c1, P2/p2, M5/m5 = 52. Еще больше зубов — 54 — имелось у морганукодонов (Morganucodon). Нижнечелюстные зубы триконодонтов (Triconodonta) имеют формулу i4, c1, p4, m5, у ряда других мезозойских групп число коренных в нижнем зубном ряде достигает 8. В каждой половине челюсти сумчатых может иметься до 4 нижних и 5 верхних резцов, часто бывает по 4 коренных, но в большинстве случаев присутствует не более 3 предкоренных — в общей сложности порядка 50 зубов. Их количество увеличивается у сумчатого муравьеда (Myrmecobius fasciatus), зубная формула которого составляет I4/i3-4, C1/c1, P3/p3, dP1-2/dp1-2? M4/m4 = 48-56. С другой стороны у хоботноголового кускуса (Tarsipes rostratus) происходит редукция зубного ряда — I2/i1, C1/c0, P1/p0, M2/m2 = 18.
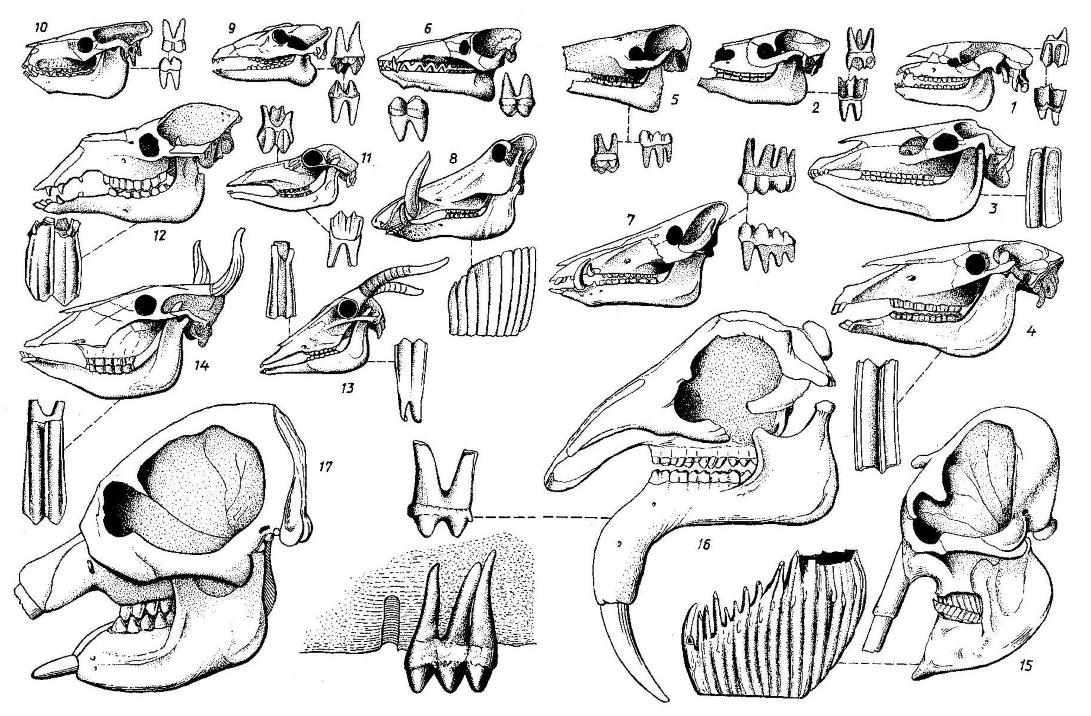
Прорисовки черепов с нижними челюстями и щечных зубрв различных копытных и хоботных: 1 — палеотерий; 2 — анхитерий; 3 — гиппарион; 4 — лошадь; 5 — палеохер; 6 — энтелодон; 7 — свинья; 8 — бородавочник; 9 — гиопотам; 10 — ореодон; 11 — олень; 12 — верблюд; 13 — антилопа; 14 — бык; 15 — слон; 16 — дейнотерий; 17 — мастодонт.
На заре возникновения плацентарных млекопитающих у них установился зубной набор I3/i3, C1/c1, P4/p4, M3/m3 = 44. Данное число принимается для группы в качестве исходного, а такая формула называется полной. Однако лишь немногие животные сохранили ее в точности; у большинства других она претерпела дальнейшие изменения. При этом прибавление каких-либо дополнительных зубов в полную формулу наблюдается редко. Например, у большеухой лисицы (Otocyon megalotis) происходит увеличение числа коренных, что в целом дает от 46 до 50 зубов — I3/i3, C1/c1, P4/p4, M3-4/m4-5. Только у форм, перешедших к пище, не требующей предварительного измельчения, число зубов, параллельно с их упрощением, может значительно увеличиваться, что облегчает их роль как захватывающего аппарата. К примеру, у гигантского броненосца (Priodontes maximus) насчитывается 80-100 зубов, а у лаплатского дельфина (Pontoporia blainvillei) — от 200 до 242. Как видно из приведенных примеров, количество зубов может существенно варьировать не только в пределах крупной систематической группы, но и у различных особей одного и того же вида.
Гораздо чаще исходное число зубов уменьшается, причем иногда очень значительно. Так, у грызунов исчезают клыки и в той или иной степени редуцируются предкоренные. Наконец, в случае полной ненадобности, зубы могут совсем исчезнуть. Их нет у панголинов (Manidae) и муравьедов (Vermilingua), практически полностью перешедших к питанию колониальными насекомыми. У некоторых млекопитающих, лишенных зубов во взрослом состоянии, последние закладываются в период зародышевого развития (усатые киты (Mysticeti) или развиваются в первые недели жизни (утконос (Ornithorhynchus anatinus)), однако затем их развитие останавливаются и они рассасываются, покрываясь сверху роговыми пластинками. У ехидн (Tachyglossidae) нет даже эмбриональной закладки зубов.

Конвергентное сходство в строении черепа и зубов некоторых сумчатых (А-В) и плацентарных (Г-Е) млекопитающих: А — вомбат (Vombatus ursinus); Б — сумчатый волк (Thylacinus cynocephalus); В — тилакосмил (Thylacosmilus atrox); Г — бобр (Castor fiber); Д — волк (Canis lupus); Е — эвсмил (Eusmilus sicarius).
вперед
в оглавление