в оглавление
назад
Зубы (продолжение)
Подобно резцам и клыкам, сложные щечные зубы млекопитающих развились из однородного ряда конических зубов примитивных зверообразных. Однако у более продвинутых зверообразных, а затем и у ранних млекопитающих, возникло такое разнообразие их форм, что установить последовательность преобразований в каждом отдельно взятом случае просто не представляется возможным.
Уже ранними исследователями относительно механизма происхождения щечных зубов высказывались 2 прямо противоположных мнения. Согласно теории конкреции, сложный зуб образовался путем слияния между собой соседних конических зубов, которые со временем преобразовались в отдельные бугорки и гребни. По теории дифференциации, разработанной Э. Копом и Г. Осборном, такой зуб возник в результате последовательного усложнения отдельного конического зуба. В дальнейшем, после привлечению онтогенетических и палеонтологических данных, подтвердилась вторая точка зрения. Что касается последовательности формирования рельефа коронок, то она окончательно не установлена и является предметом споров. В свое время для объяснения данного процесса охотно апеллировали к уже упомянутой теории Копа-Осборна. Позднее завоевала популярность теория амфикона. Сегодня представляется вероятным, что обе эти точки зрения частично верны, причем теория Копа-Осборна применима по отношению к нижнечелюстным коренным, а теория амфикона — к верхнечелюстным. Не вдаваясь в тонкости и противоречия обеих теорий, изложим обобщенный взгляд на проблему.
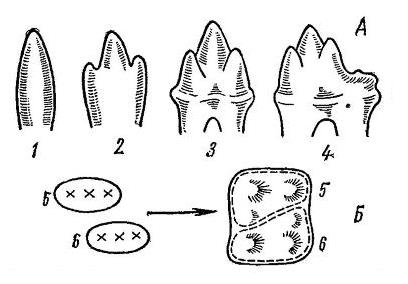
Происхождение коренных зубов по теории дифференциации (А) и по теории конкреции (Б): 1 — гаплодонтная форма; 2 — триконодонтная форма: 3 — тритуберкулярная форма; 4 — туберкуло-секториальный зуб; 5-6 — образование многобугорчатого зуба путем слияния 2 триконодонтных зубов.
Итак, изначально у зверообразных имелись лишь простые конические, или гаплодонтные, зубы с единственной вершиной. Эта исходная вершина на верхнечелюстных зубах именуется эоконом (eocon), а на нижнечелюстных является гомологом протоконида более поздних форм. Гаплодонтные зубы годились только для отрывания кусков пищи, их жевательная функция была абсолютно не развитой.
У цинодонтов (Cynodontia) щечные зубы претерпевают усложнение. Прежде всего, с внутренней стороны верхнечелюстных зубов появляется маленькая дополнительная вершинка — протокон (рrotoсonus), при чсыкании челюстей попадающая между соседними нижними зубами. Затем эокон начинает разделяться на переднюю и заднюю вершины; образуется наполовину раздвоившаяся вершина — амфикон (amphicon). В переднем и заднем углу наружного отдела развиваются дополнительные возвышения — соответственно, парастиль (parastyle) и метастиль (metastyle). В то же время, дополнительные вершинки — гомологи будущих параконида и метаконида — появляются и на основном валике нижнечелюстных зубов, впереди главной вершины и позади нее. Основной валик, или воротничок (cingulum), также является приобретением цинодонтов. У них он представляет собой выступ эмали с внутренней стороны нижней части коронки. Судя по всему, воротничок появился для защиты десен. Данная стадия усложняющегося конического зуба называется протодонтной.
Дальнейшее расщепление амфикона и сильное увеличение протокона приводят к образованию на верхнечелюстных зубах тригона (trigon), объединяющего 3 вершины — центральный протокон, занимающий теперь главенствующее положение, и полученные из амфикона переднюю и заднюю дополнительные вершины — паракон (paraconus) и метакон (metaconus). На нижнечелюстных зубах главная вершина тригонида (trigonidum) именуется протоконидом (рrotoconid), а передняя и задняя дополнительные вершины — соответственно параконидом (paraconid) и метаконидом (metaconid). И верхние, и нижние дополнительные вершины постепенно вырастают, и зубы становятся трехвершинными, причем все вершины распределяются на одной продольной линии. Зубы такого типа именуются трехзубчатыми, или триконодонтными. Они действовали по типу ножниц, помогающих разгрызать корм. Подобные зубы характерны для ряда примитивных мезозойских хищников — например, для триконодонтов (Triconodonta).
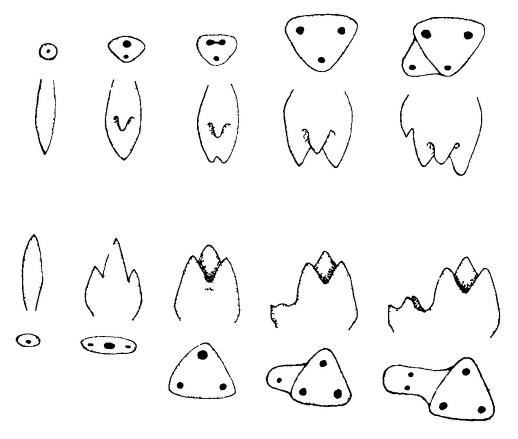
Схемы, иллюстрирующие эволюционное развитие структурных особенностей верхних и нижних коренных зубов. В верхнем и нижнем рядах зубы показаны с окклюзионной стороны; основные вершины обозначены черными точками. На всех изображениях щечный край обращен вверх, а передний — вправо. В 2 средних рядах эти же зубы показаны с языковой стороны, они тоже обращены своей передней стороной вправо. На верхних зубах первичная вершина (эокон) разделяется на паракон и метакон; протокон является новообразованием, а пятка (талон) мала или отсутствует. На нижних зубах к первичной вершине (протокониду) вначале прибавляются передняя и задняя вершины (параконид и метаконид), затем зуб становится треугольным и, наконец, сзади добавляется пятка (талонид) с 2 дополнительными вершинами (гипоконидом и энтоконидом).
Последующая эволюция зубной системы идет по пути усложнения трехвершинного зуба; от общей линии развития уклоняются лишь аллотерии (Allotheria) — архаичная вымершая группа, происхождение которой до конца не выяснено. Ее наиболее характерные представители, многобугорчатые (Multituberculata), имели удлиненные коренные с 2-3 продольными рядами вершинок, разделенными продольными бороздами. Вместе с тем, последний нижний предкоренной в большинстве случаев сильно удлинен и очень высокий, с продольным режущим срединным лезвием, зубчатым на вершине, и с вертикально бороздчатыми боковыми сторонами; передние предкоренные значительно меньше, иногда также со срединным режущим лезвием.
В трехвершинном зубе постепенно смещается линия вершин: на верхнечелюстных зубах средняя вершина — протокон — сдвигается внутрь, а передняя и задняя — паракон и метакон — наружу. В свою очередь, на нижнечелюстных зубах протоконид отодвигается наружу, а параконид и метаконид — внутрь, т. е. происходит процесс, обратный наблюдаемому в зубах верхней челюсти. Таким образом, коронка коренного приобретают общие очертания прямоугольного треугольника, по углам которого распределены острые вершины. На верхних коренных вершина треугольника обращена к языку (лингвально), на нижних — к щеке (буккально). Такую коронку называют трибосфенической, или тритуберкулярной (трехбугорчатой). При смыкании челюстей коренные не встречаются с коренными им противолежащими, а вершинами своих треугольных площадок вклиниваются между ними. При этом острые гребни сторон треугольников, проходя вплотную вблизи друг друга, действуют как разрезающий аппарат. И у мезозойских, и у более поздних хищников зубы с вершинами, соединяющимися между собой острыми гребнями, именуются бугорчато-режущими (туберкуло-секториальными), или секодонтными.
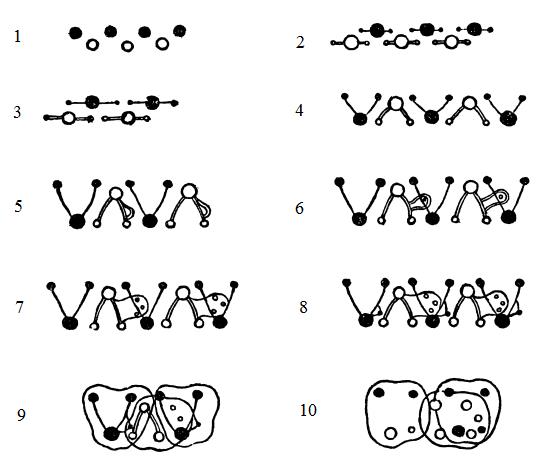
Схема эволюции коронок щечных зубов млекопитающих в представлении Осборна; вершины зубов верхней челюсти отмечены черными кружочками, вершины зубов нижней челюсти — белыми кружочками: 1 — гаплодонтная стадия; 2 — протодонтная стадия; 3 — триконодонтная стадия; 4 — тригонодонтная стадия; 5 — переход зубов в нижней челюсти к туберкуло-секториальной стадии; 6 — в нижней челюсти туберкуло-секториальные зубы, на талониде гипоконид, в верхней — тригонодонтные; 7 — на пятке зубов нижней челюсти образуются дополнительные бугорки; 8 — в верхней и нижней челюсти туберкуло-секториальные зубы; 9 — переход к четырехбугорчатым зубам; 10 — четырехбугорчатые зубы.
Трибосфенические зубы свойственны многим мезозойским млекопитающим — например, т. н. симметродонтам (Symmetrodonta). Судя по всему, именно от предка с подобными зубами произошли все териевые млекопитающие. Вполне вероятно, что трибосфенические коронки развились у однопроходных и териев независимо друг от друга, а не были унаследованы от общего предка. Однако данная точка зрения остается спорной, поскольку строение зубной системы у некоторых ранних однопроходных и дотрибосфенических териеобразных довольно сходно. Кроме того, для подобных выводов родственные связи мезозойских млекопитающих пока изучены явно недостаточно.
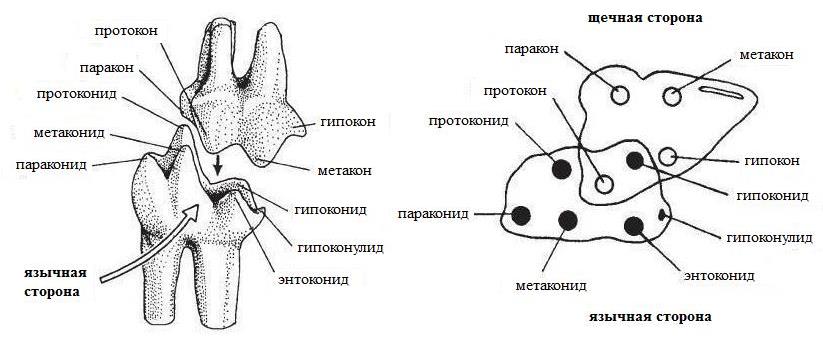
Схема смыкания примитивно устроенных верхнего и нижнего трибосфенических зубов.
Более существенное усложнение трибосфенического зуба проявляется в том, что позади основания коронки образуется особый округлый выступ — пятка. Первоначально пятка ниже и меньше по размерам, чем передняя часть зуба, но затем она увеличивается и на ней появляются в различной степени развитые вершины. На верхнечелюстных коренных пятка формируется позади протокона и именуется талоном (talon). На талоне развивается задняя внутренняя вершина — гипокон (hypoconus), который иногда вырастает из задневнутреннего воротничка, иногда же образуется путем отделения от протокона (в первом случае его называют ложным гипоконом, во втором — истинным). Нередко между протоконом и параконом с метаконом образуются маленькие промежуточные вершинки: передняя — протоконуль (protoconulus) и задняя — метаконуль (metaconulus). На нижнечелюстных коренных пятка обнаруживается раньше и обычно развивается в большей степени, чем на верхнечелюстных. Она находится позади тригонида и называется талонидом (talonid). С течением времени талонид расширяется, удлиняется и принимает чашевидную форму со впадиной (бассейном) в середине. По краям талонида появляются дополнительные вершинки: задняя наружная — гипоконид (hypoconid), задняя промежуточная — энтоконид (еntoconid) и задняя внутренняя — гипоконулид (hypoconulid).
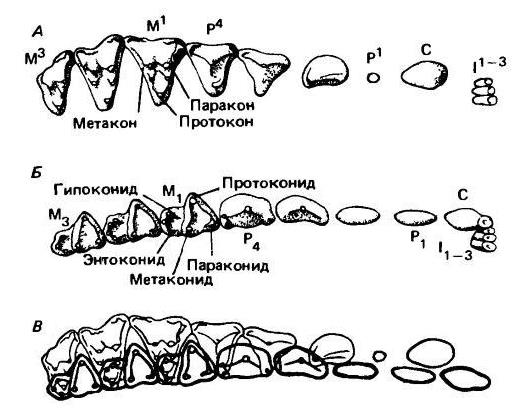
Схемы зубной системы мелового млекопитающего дидельфодона (Didelphodon sp.): А — правые верхние зубы со стороны коронок; Б — левые нижние зубы со стороны коронок; В — зубные ряды в положении окклюзии — контуры нижних зубов (показаны жирными линиями) наложены на контуры верхних. Каждый верхний зуб лежит кзади и кнаружи от соответствующего ему зуба нижней челюсти. Оба ряда частично заходят один за другой, производя разрезающее действие вдоль диагональной линии между каждым верхним зубом и лежащим за ним нижним. Прохождение верхнего зуба задерживается пяткой (талонидом) на соответствующем нижнем коренном; протокон входит во впадину талонида.
При смыкании челюстей талонидный бассейн обеспечивает более прочный упор для протокона верхнего зуба, гипокон которого упирается теперь в тригонидный бассейн последующего нижнего зуба, а энтоконид талонида — во впадину между протоконом и гипоконом верхнего, подобно пестикам в ступках. Таким образом, в коренных данного типа к режущему действию, осуществляемому, как и прежде, острыми гребнями сторон тригона и тригонида, присоединяется зачаточное давящее или дробящее действие в области талонида, позволяющее измельчать семена, мелкие плоды и пр. Благодаря развитию раздавливающей способности зубов появляется возможность перехода от сугубо хищного (главным образом, насекомоядного) образа жизни к частичной или полной всеядности. Такого рода зубы характерны для начальных стадий многих ветвей млекопитающих, прослеживающихся с позднего мезозоя: для простейших всеядных сумчатых, многих насекомоядных в широком понимании, примитивных хищных, архаичных приматов и некоторых других. Подобный тип зубов содержит в зачатке возможности развиваться как в сторону усиления хищного, режущего типа, так и в сторону усиления растительноядности, вплоть до исключительно растительного режима и питания очень жесткими частями растений.
Примечательный случай сходной эволюции щечных зубов зафиксирован у шуотеридий (Shuotheridia) — малочисленной группы мезозойских млекопитающих. В отличие от настоящего талонида, псевдоталонид их нижних коренных находится не позади тригонида, а впереди него. Подобное строение коронок было названо псевдотрибосфеническим. Указанная группа просуществовала недолго и исчезла, не оставив потомков.
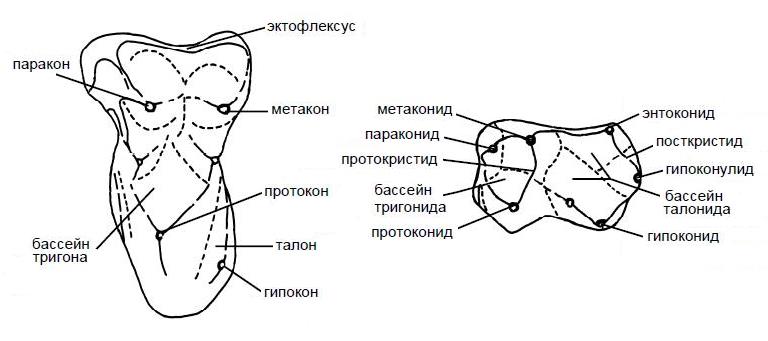
Схема строения и номенклатура полностью сформированных верхнего и нижнего трибосфенических коренных зубов.
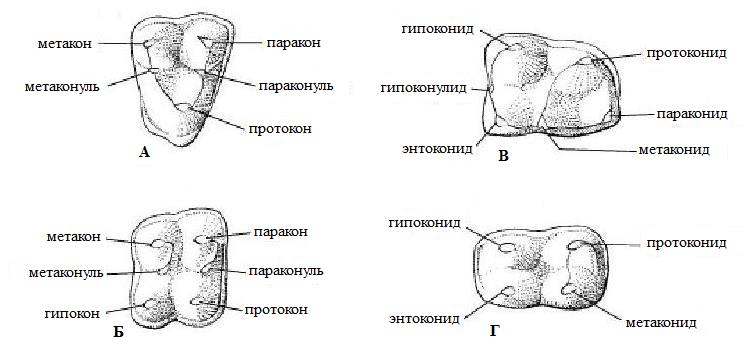
Схемы рельефа коренных зубов плацентарных млекопитающих. На всех изображениях наружный край зуба обращен вверх, а передний край — вправо. А — правый верхний коренной примитивной формы; Б — правый верхний коренной, приобретший прямоугольные очертания благодаря добавлению гипокона на задневнутреннем углу; В — левый нижний коренной, имеющий шесть вершин; Г — левый нижний коренной, приобретший прямоугольные очертания благодаря утрате параконида.
При переходе от всеядного режима к питанию крупной теплокровной добычей усиливается режущая и ослабевает давящая способность зубов. Как и прежде, режущая функция сосредотачивается в заднем отделе щечного ряда, однако режущие лезвия теперь не поперечные, как у древнейших форм, а продольные, что более эффективно. Щечной зуб, режущая способность которого выражена наиболее резко, называется хищническим (dens sectorius). В каждой половине верхней и нижней челюсти их по 2 у креодонтов (Creodonta) и по 1 у хищных (Carnivora). У креодонтов в роли хищнических зубов чаще всего выступают I-й и II-й верхние коренные и II-й и III-й нижние коренные, у хищных это всегда IV-й верхний предкоренной и лежащий позади него I-й нижний коренной. На верхних хищнических зубах креодонтов лезвие образуется параконом, метаконом и вытянутым в длину метастилем; паракон и метакон иногда сливаются в одну вершину. У хищных форму лезвия на верхнем плотоядном зубе принимает еще не разделенный амфикон и, кроме метастиля, вытягивается и принимает режущую форму и парастиль. Все вершины соединяются в одно вытянутое вдоль челюстной оси челюсти лезвие, на котором имеются вырезки, отмечающие границы исходных вершин. Одновременно уменьшается в размерах протокон, давящая функция которого ослабевает, а в высшем выражении хищного типа он, как и несущий его внутренний выступ зуба, совсем исчезает. На нижних хищнических зубах лезвие образуется параконидом, постепенно передвигающимся из исходного положения внутри коронки вперед и наружу, на один уровень с наружным конусом — протоконидом; как и на верхних, оба лезвия исходных конусов разделены на вершине выемкой. Пересекаясь при смыкании челюстей, верхние и нижние хищнические зубы работают наподобие ножниц, разделяя на куски плотные ткани — мясо, сухожилия, частично кости.
Пока вместе с мясной пищей употребляется и растительность (например, у собак (Canidae)), сохраняется низкий талонид, противостоящий уменьшенному верхнему I-му коренному. С переходом к исключительно мясоедению талонид, попутно с I-м верхним коренным, уменьшается и совсем исчезает (гиенодонты (Hyaenodontidae), кошки (Felidae), некоторые гиены (Hyaenydae) и др.). Кроме того, уменьшается и исчезает утративший свою функцию метаконид. Исчезают и лежащие позади хищнеческих зубов коренные, что происходит отчасти в связи с вытесняющими их удлиняющимися хищнеческими зубами, отчасти, когда они сохраняют исходное бугорчатое строение, — вследствие утраты ими своих функций (у настоящих хищных).
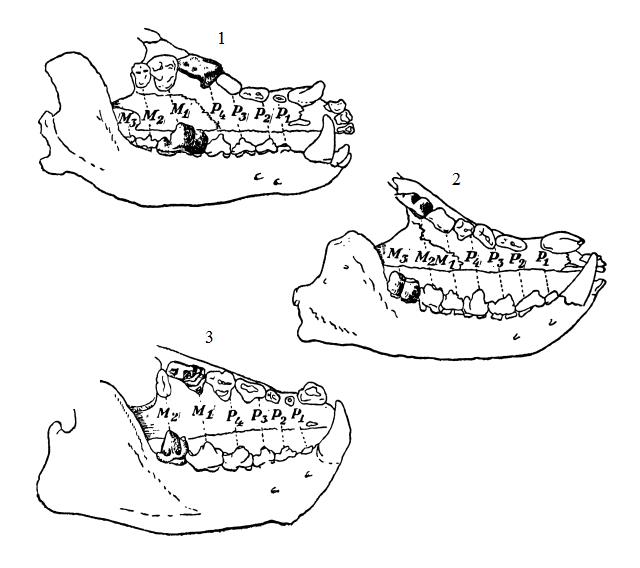
Три различных пути развития хищнических зубов у плотоядных млекопитающих: 1 — волк (Canis lupus); гиенодон (Hyaenodon sp.); оксиена (Oxeaena sp.).
Иначе идет развитие коренных при усилении растительного режима питания. Как правило, зубы в этом случае становятся четырехугольными за счет усиления их задних отделов — в первую очередь, пятки. Гипокон на талоне может достигнуть размеров 3 основных вершин, что обусловливает появление квадритуберкулярного, или четырехбугорчатого, зуба. Внизу четырехугольная форма может достигаться несколькими способами, например путем утраты параконида на переднем крае тригонида. Четырехбугорчатые коренные, в частности, характерны для ежей (Erinaceidae), енотов (Procyonidae) и многих приматов. В дальнейшем число основных вершин на зубах может увеличиваться, достигая 5 и даже 6.
У млекопитающих всеядных и с уклоном в растительноядность вершины коренных понижаются и становятся округлыми, а связывающие их режущие гребни исчезают. В этом случае говорят о тупобугорчатых, или бунодонтных, зубах. Зачастую они одновременно и четырехбугорчатые. Зубы такого рода хорошо выполняют функцию раздавливания относительно мягкой пищи — мяса, листьев, луковиц, плодов и т. д. Ярко выраженная бунодонтия свойственна всеядным формам, например большинству медведей (Ursidae), свиньям (Suidae), пекари (Tayassuidae), человекообразным обезьянам (Hominoidea). Близки по строению щечные зубы ежей (Erinaceidae) и хомяков (Cricetidae).
В бунодонтном зубе передние и задние вершины и возвышения могут сливаться, образуя продольно либо поперечно сориентированные гребни с эмалевыми режущими краями, которые иногда покрываются мелкими бугорками. Такой тип зубов именуется складчатым, или лофодонтным. На верхних коренных в этом случае имеется 3 основных гребня, или лофа, — задний, или металоф (metaloph), передний, или протолоф (protoloph), и наружный, или эктолоф (ectoloph). От них внутрь могут отходить добавочные гребни, или складки. От передней поверхности металофа отходит шпора, или кроше (crohet), от задней поверхности протолофа — противошпора, или антекроше (antecrohet), а от эктолофа — криста (crista); последних на коронке может быть несколько. От задней поверхности металофа назад может отходить задняя шпора, или посткроше (postcrochet). Поначалу эти добавочные выступы небольшие, но в дальнейшем получают тенденция разрастаться и соединяться друг с другом. На нижних коренных наличествует 2 основных гребня, или лофида, — передний, или металофид (metalophid), и задний, или гиполофид (hypolophid). Первый гребень дважды коленообразно изгибается, задний — дуговидный или коленообразно изогнутый. Между гребнями расположены углубления — долинки.
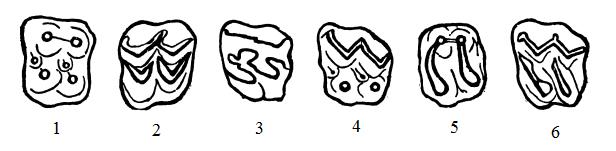
Различные типы жевательной поверхности коренных зубов копытных — бунодонтный (1), селенодонтный (2) и лофодонтный (3) типы, а также буноселенодонтный (4), лофобунодонтный (5) и лофоселенодонтный (6) смешанные типы.
Простые лофодонтные зубы обычны у млекопитающих, питающихся листьями и тонкими веточками деревьев и кустарников. В ходе приспособления к перетиранию более жесткой, богатой клетчаткой растительности (в первую очередь травы, поедание которой к тому же сопровождается захватом частиц почвы), резко усиливающей стирание коронок, сложность их строения возрастает. Зубы с 2 поперечными гребнями называются билофодонтными; они характерны для тапиров (Tapiridae), сирен (Sirenia), многих приматов (Primates), зайцеобразных (Lagomorpha) и многих грызунов (Rodentia). Простейшие лофодонтные зубы с низкими коронками имеются у белок (Sciuridae). У землекопов (Bathyergidae) гребни сливаются в проходящее по краю коронки кольцо. 2 поперечно посаженных гребня на коренном зубе носорогов (Rhinocerotidae) дополнительно соединяются гребнем, идущим вдоль наружной стенки. Жевательные поверхности лошадей (Equidae) представляют собой результат дальнейшего усложнения билофодонтных зубов и содержат целый комплекс продольных и поперечных гребней. Путем постепенного увеличения числа и протяженности поперечно направленных гребней вырабатываются полилофодонтные зубы, характерные для слонов (Еlephantidae), водосвинок (Hydrochoerinae) и болотных крыс (Otomyini). При этом наблюдается вытягивание гребней в пластинки и правильное чередование эмали, дентина и цемента.
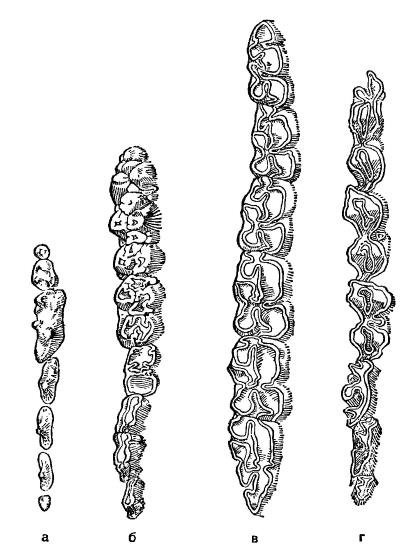
Разные типы жевательной поверхности коренных зубов млекопитающих: а — бугорчато-режущий у лисицы (Vulpes vulpes); б — тупобугорчатый у кабана (Sus scrofa); в — складчатый у кулана (Equus hemionus); г — лунчатый у зубра (Bison bonasus).
Еще одним производным бунодонтных зубов являются бугорчато-лунчатые, или бунолесодонтные, зубы. Они встречаются у копытных, питающихся листьями древесных пород, например, у некоторых парнопалых (антракотериев (Anthracotheriidae) и др). Наружные вершины на верхних и внутренние вершины на нижних коренных зубах в этом случае преобразуются в полулунные гребни. При этом иногда формируются различные ребра и стили, в частности, пара-, мезо- и метастиль. Как и бугорчатые, бугорчато-лунчатые зубы чаще всего имеют низкую коронку. По мере перехода к питанию травянистой растительностью подобные зубы превращаются в лунчатые, или селенодонтные. Лучше всего этот тип выражен у жвачных (Ruminantia), особенно у позднейших их представителей. Основные 4 вершины на их коренных зубах приняли полулунную форму. Эти 4 полулуния глубокими выемками (внутренними на верхних зубах, наружными — на нижних) делятся на переднюю и заднюю колонки, каждая из которых заключает по 2 полулуния — наружное и внутреннее. При этом нередко возникают дополнительные элементы, осложняющие строение коронки. Среди лунчатозубых парнопалых встречаются формы с наиболее высокой коронкой коренных. Параллельно усложнению коронок вместо исходного для млекопитающих чередования щечных зубов развивается их противостояние.
У некоторых грызунов и частью у других эвтериев передний и задний воротнички верхних коренных зубах разрастаются, становясь похожими на основные гребни и вместе с ними формируют т. н. тетралофодонтную зубную коронку. На нижних коренных этой конструкции соответствуют тригонид и расположенные позади от него талонид. При этом протоконид смещен к наружному краю коронки, пара- и метаконид — к внутреннему, гипоконид расположен в ее передне-губной части, а между ним и тригонидом заключен углубленный бассейн талонида.
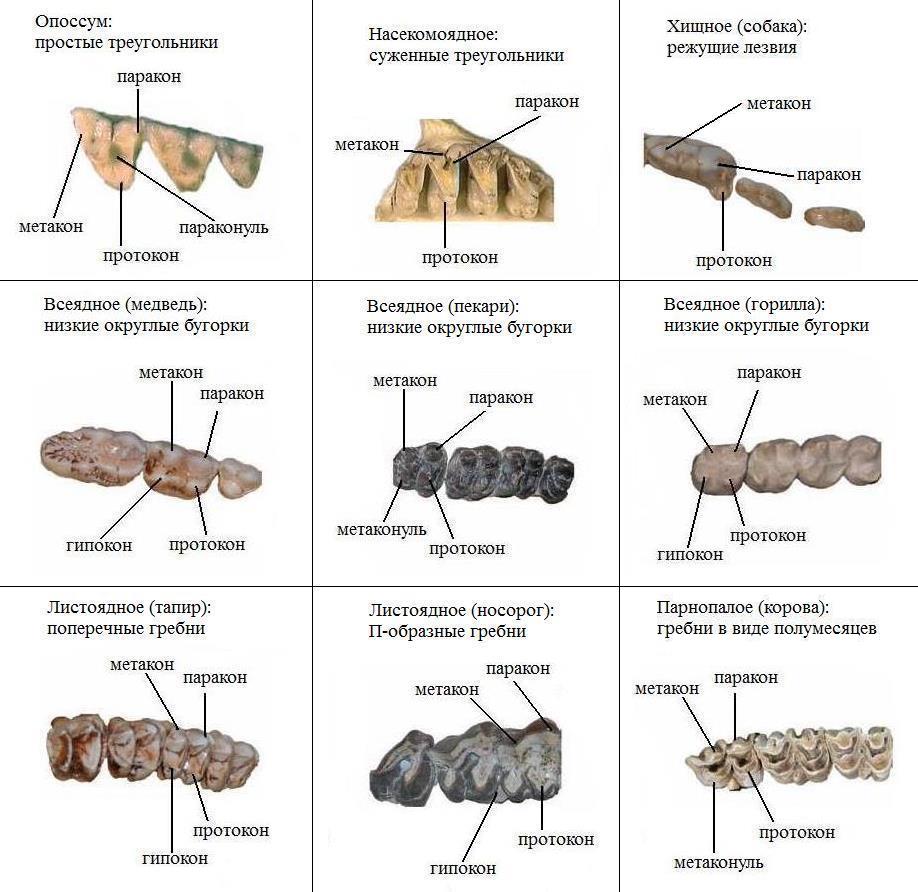
Таким образом, номенклатура основных элементов жевательной поверхности для коренных верхней челюсти такова:
протокон (рrotoсonus) — передняя внутренняя вершина;
паракон (рaraconus) — передняя наружная вершина;
гипокон (hypoconus) — задняя внутренняя вершина;
метакон (metaconus) — задняя наружная вершина;
протоконуль (рrotoconulus) — передняя промежуточная вершина;
метаконуль (metaconulus) — задняя промежуточная вершина;
тригон (trigon) — часть коронки, объединяющая протокон, паракон и метакон;
талон (talon) — часть коронки позади тригона, несущая гипокон, протоконуль и метаконуль;
протолоф (protoloph) — передний поперечный гребень, в состав которого входят протокон и протоконуль;
эктолоф (ectoloph) — наружный гребень, в состав которого входят парастиль, паракон, мезостиль, метакон и метастиль;
металоф (metaloph) — задний поперечный гребень, в состав которого входят гипокон и метаконуль;
главная, или средняя, долинка (medisinus, medifossette) — впадина между протолофом и металофом;
задняя долинка (postsinus, postfossette) — впадина позади металофа;
шпора, или кроше (crohet), — складка передней поверхности металофа в среднюю долинку;
противошпора, или антекроше (antecrohet), — складка задней поверхности протолофа в среднюю долинку;
задняя шпора, или посткроше (postcrochet), — складка задней поверхности металофа в заднюю долинку;
криста (crista) — складка внутренней поверхности эктолофа в среднюю долинку;
добавочная долинка — впадина, отшнурованная снаружи от средней долинки слиянием шпоры и кристы;
поясок, или воротничок (cingulum), — кайма, окружающая основание коронки;
Для коренных нижней челюсти номенклатура следующая:
параконид (рaraconid), или парастилид (parastylid), — передняя внутренняя вершина;
метаконид (metaconid) — передняя промежуточная, или средняя внутренняя, вершина;
протоконид (рrotoconid) — передняя наружная вершина;
гипоконулид (hypoconulid) — задняя внутренняя вершина;
энтоконид (еntoconid) — задняя промежуточная, или задняя внутренняя, вершина;
гипоконид (hypoconid) — задняя наружная вершина;
тригонид (trigonidum) — часть коронки, несущая протоконид, параконид и метаконид;
металофид (metalophid) — передний гребень, состоит из передней, наружной и задней ветвей (в форме буквы П);
гиполофид (hypolophid) — задний гребень, состоит из наружной и задней ветвей;
передняя внутренняя долинка (antesinus) — впадина между передней и задней поперечными ветвями металофида;
задняя внутренняя долинка (postsinus) — впадина между металофидом и гиполофидом;
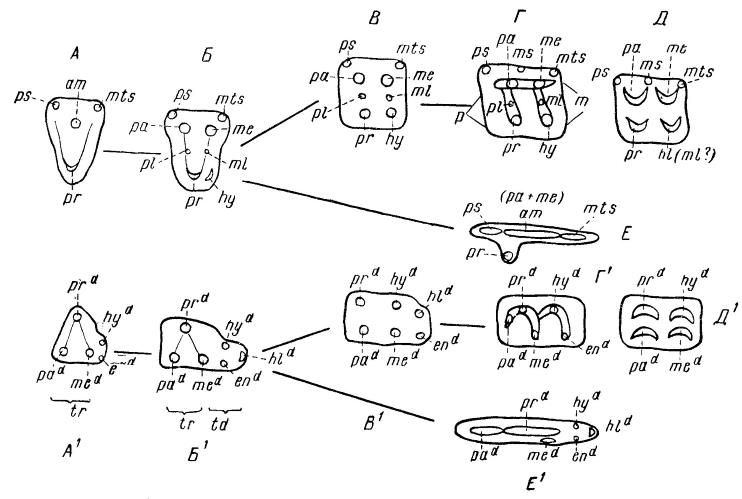
Схемы коренных (верхние A-E, нижние Al-El) зубов млекопитающих разных типов. Элементы: ра — паракон; me — метакон; рr — протокон; hу — гипокон; аm — амфикон; рs — парастиль; ms — мезостиль; mts — метастиль; рl — протоконуль; ml — метаконуль; р — протолоф; m — металоф; pad — параконид; med — метаконид; prd — протоконид; hуd — гипоконид; еnd — энтоконид; hld — гипоконулид; tr — тригонид; td — талонид. Верхний ряд — верхние зубы, нижний — нижние.
Зубы с короткой коронкой, высота которой уступает ее длине и ширине, хорошо развитыми корнями и ограниченным ростом называют брахиодонтными. Эмалевый колпачок покрывает их коронку и простирается вплоть до шейки, тогда как цемент распространен не выше границы десен. Подобное строение зубов является исходным и сохраняется у плотоядных, всеядных и примитивных растительноядных форм — насекомоядных, рукокрылых, хищных, неспециализированных групп копытных. Однако питание травой ставит перед растительноядными млекопитающими серьезную проблему, потому что она является жестким, грубым материалом, который оказывает на жевательную поверхность щечных зубов особенно высокую абразивную нагрузку. Таким образом, если бы низкокоронковые зубы сохранялись у травоядных форм, они бы в скором времени снашивались до самых корней. В связи с этим обстоятельством развиваются гипсодонтные зубы, коронка которых по высоте обычно превосходит длину и зачастую имеет призматическую форму. Усложнение, противостояние и повышение коронок делают работу зубов максимально продуктивной.
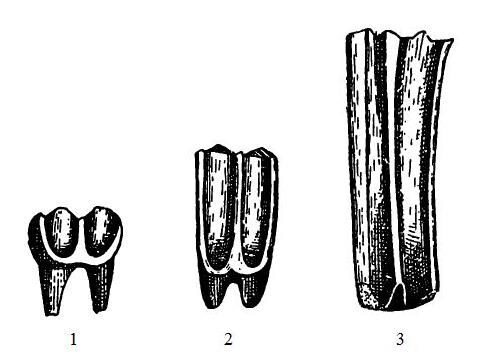
Стадии повышения коронки коренных зубов у лошадиных: 1 — брахиодонтный зуб анхитерия (Anchitherium); 2 — гипселодонтный зуб гиппотерия (Hippotherium); 3 — призматический зуб настоящей лошади (Equus).
Образование простейших гипсодонтных зубов происходит посредством удлинения наполненного дентином тела зуба с сохранением первоначальной формы вершин на жевательной поверхности. Впервые такие зубы сформировались у некоторых низших форм, но оказались неудачными. После того, как эмалевая поверхность стирается, основная нагрузка ложится на относительно мягкий и легко стирающийся дентин; прочная эмаль остается только в виде тонкого ободка. Преуспевающие формы имеют гипсодонтные зубы, построенные совсем по другой схеме. Высота их коронки увеличивается посредством роста каждой вершины или, чаще, гребня наподобие небоскреба; эти тонкие пики сращены воедино цементом, распространившимся перед прорезыванием по поверхности всего зуба. По мере изнашивания зуба происходит стирание всего комплекса его твердых слоев: эмали, дентина и цемента. Поскольку это стирание оказывается неравномерным, рабочая поверхность зуба получается ребристой, как терка. Кроме того, благодаря позднему или неполному развитию корней без шеек такие зубы могут обладать длительным ростом, что также призвано компенсировать их износ.
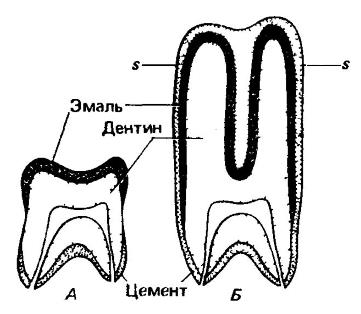
Схемы, показывающие способ образования гипселодонтного зуба. А — обычный брахиодонтный зуб; Б — гипселодонтный зуб, имеющий очень высокие вершины и гребни и целиком покрытый цементом. При любой степени стирания коронки такого зуба, например до уровня s-s, на жевательной поверхности будет представлено не менее девяти следующих друг за другом слоев цемента, эмали и дентина.
Высококоронковость свойственна многим растительноядным млекопитающим. Так, зайцы (Leporidae), населяющие пустынные районы и питающиеся жесткими растениями, к которым постоянно примешивается песок, имеют более высокие зубы по сравнению со своими сородичами из других местностей. Повышение и более или менее продолжительный рост коронок также наблюдается у части современных сумчатых, грызунов и всех без исключения зайцеобразных, а среди вымерших групп — у некоторых тиллодонтов (Tillodonta), нотоунгулят (Notoungulata) и т. д. Постоянным ростом характеризуются щечные зубы ряда ныне живущих копытных, например лошадей (Equidae) и полорогих (Bovidae). Из плотоядных форм продолжительный рост характерен для верхних клыков моржей (Odobenidae) и ископаемых тилакосмилов (Thylacosmilidae). Однако самыми известными примерами гипселодонтии, несомненно, являются постоянно растущие резцы хоботных, грызунов и зайцеобразных.
вперед
в оглавление