в оглавление
назад
Череп
Голова (caput) является вместилищем головного мозга и важнейших органов чувств, местом первоначальной обработки пищи и начала дыхательных путей. Все эти функции накладывают отпечаток на скелет головы — череп (cranium), строение которого сильно варьирует в зависимости от образа жизни и филогенетических связей животного. Как и у других позвоночных, в черепе млекопитающих различают 2 отдела — невральный, или мозговой (neurocranium), вмещающий мозг и главные органы чувств, и висцеральный, или лицевой (splanchnocranium), охватывающий переднюю часть пищеварительного тракта.
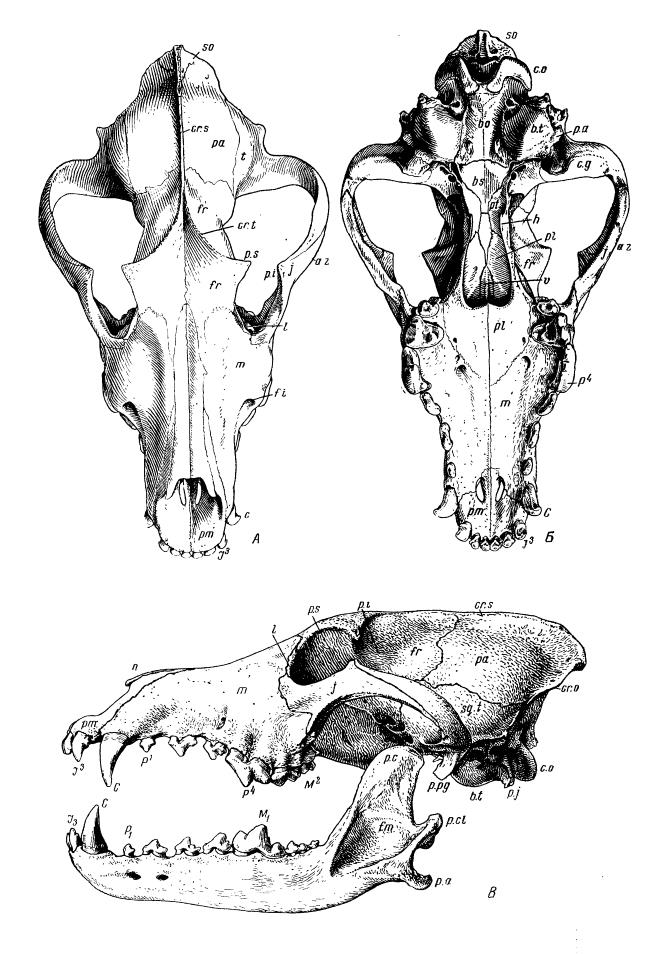
Череп с нижней челюстью волка (Canis lupus), вид сверху (А), снизу (Б) и сбоку (В): so — верхняя затылочная кость (supraoccipitale); bo — основная затылочная кость (basioccipitale); с. o — затылочные мыщелки (condyles occipitales); pa — теменная кость (parietale); сr. s — стреловидный, или сагиттальный, гребень (crista sagittalis); сr. o — затылочный гребень (crista occipitalls); b. t — слуховой пузырь (bulla tympani); р. а — слуховое отверстие (porus acusticus); bs — основная часть клиновидной кости (basisphenoideum); c. g — суставная впадина (cavitas glenoidalis); р. рg — засуставный отросток (proс. postgleonoideus); p. j — яремный отросток (proс. jugularis); t — височная кость (temporale); sq. t — чешуя височной кости (squama temporalis); fr — лобная кость (frontale); сr. t — височные гребешки или линии (crista temporalis); а. z — скуловая дуга (arcus zygomaticus); j — скуловая кость (jugale); p. s — верхний заглазничный отросток (proс. postorbitalis superior); p. i — нижний заглазничный отросток (proс. postorbitalis inferior); l — слезная кость (lacrimale); pl — небная кость (palatinum); pt — крыловидная кость (pterygoideum); h — крючочки (hamuli); m — верхнечелюстная кость (maxlllare); n — носовая кость (nasale); f. i — подглазничное отверстие (foramen infraorbitale); pm — межчелюстная кость (praemaxillare); p. с — венечный отросток (proс. coronoideus); p. сl — суставный отросток (рrос. condyloideus); p. a — угловой отросток (proс. angularis); f. m — массетерная ямка (fossa masseterica); I — резцы (incisivi); C — клык (caninus); P — предкоренные зубы (praemolares); М — коренные зубы (molares).
Череп млекопитающих относительно велик и тяжел, как и голова в целом. По завершении роста он полностью окостеневает, а многие кости сливаются между собой в комплексы. В ходе процесса упрощения черепа млекопитающих с каждой его стороны возникла лишь одна височная впадина, и образовалась единственная скуловая дуга, в связи с чем его относят к синапсидному типу. Конструкция висцеральных компонентов черепа глубоко своеобразна, уникальна среди позвоночных, в чем проявилось смешанное влияние адаптации к восприятию звуков в воздушной среде (эволюция среднего уха) и к механической обработке корма.
Мозговой отдел, или просто мозговой череп, относится у млекопитающих к тропибазальному типу (с узким основанием и укороченной в продольном направлении межглазничной перегородкой). По этому признаку он сходен с черепом рептилий, однако данная особенность может сильно стушевываться. Мозговая полость и орган обоняния у млекопитающих сильно разрастаются, вследствие чего передняя черепная стенка сближается с задней стенкой носовых капсул, а глазницы отходят от средней линии, оттесняясь в бока. В результате описанных процессов тропибазальный череп млекопитающих принимает облик ложного платибазального (с широким основанием).
По сравнению с другими позвоночными число отдельных костей в мозговом черепе млекопитающих значительно уменьшено, что связано с их редукцией и образованием из них более или менее сложных комплексов. В таких комплексах кости обычно срастаются между собой поздно, что обусловливает возможность беспрепятственного длительного роста крупного головного мозга, а в дальнейшем обеспечивает прочность черепа. Во многих случаях отдельные кости настолько утрачивают свою индивидуальность, что сохраняются лишь в виде отдельных центров окостенения сложной кости. На наружной поверхности костей развиваются шероховатости и гребни, служащие для прикрепления различных мышц.
Слиянием отдельных костей в комплексы, высоким относительным объемом и прочностью мозговой коробки, ставшей надежной конструктивной основой черепа в условиях значительных механических нагрузок, сопровождающих силовую обработку пищи челюстями, млекопитающие подобны птицам. Однако, в отличие от птиц, швы между костями мозговой коробки сохраняются у млекопитающих на протяжении всей жизни. У некоторых грызунов и во взрослом состоянии имеются лобно-теменные фонтанели — неокостеневшие промежутки в плоских костях или в области межкостных швов, затянутые соединительной тканью. Расположение, размеры и форма фонтанелей могут варьировать у разных особей внутри популяции или вида и в зависимости от возраста. В отличие от родничков новорожденных человека и других приматов, подвижности костей черепа относительно друг друга они не обеспечивают. Точная функция фонтанелей у грызунов пока не ясна; осторожно предполагается их связь с работой эпифиза головного мозга.
Теменное отверстие, характерное для многих зверообразных, у млекопитающих отсутствует. Предлобные (praefrontalia) и заднелобные (postfrontalia) кости среди современных представителей класса имеются только у однопроходных. Септомаксиллярные (septomaxillare), заглазничные, надвисочные и квадратноскуловые кости также редуцируются и у вполне сформированных млекопитающих утрачены. Основная (basioccipitale), пара боковых (exoccipitalia) и верхняя затылочная (suptaoccipitale) кости у ряда мезозойских групп и однопроходных сохраняются в виде отдельных затылочных окостенений. У сумчатых и плацентарных все эти элементы сливаются в одну крупную затылочную кость (occipitale), хотя у их зародышей различимы центры окостенения всех четырех первичных элементов. Исходно парные заднетеменные кости образуют у зверообразных небольшой срединный элемент; он сохраняется у млекопитающих, но у большинства взрослых животных также срастается с затылочной костью (как, по-видимому, и табличные кости).
Затылочная кость несет на себе 2 мыщелка (condyli occipitales), сформированных за счет боковых затылочных костей и обеспечивающих подвижное сочленение черепа с I-м шейным позвонком. В этом отношении млекопитающие отличаются от рептилий и птиц, у которых имеется только 1 мыщелок. Между мыщелками находится большое затылочное отверстие (foramen occipitale magnum), соединяющее полость черепа со спинномозговым каналом. У примитивных млекопитающих оно расположено почти под прямым углом к продольной оси черепа; у приматов с сильно развитым мозгом это отверстие перемещается книзу и находится почти в одной плоскости с основанием черепа (например, у человека). Помимо большого затылочного отверстия, в основании черепа имеется много мелких отверстий для прохождения нервов и кровеносных сосудов.
Крышу лобной области составляют парные лобные кости (frontalia), иногда срастающиеся между собой в одну цельную кость (у летучих мышей, приматов). Боковые края лобных костей образуют нависающие над глазницами заглазничные отростки. Крыша теменной области образована парными теменными костями (parietalia), которые продолжаются книзу на бока мозговой коробки и нередко тоже срастаются между собой. У основания теменных костей, между ними и затылочной костью часто располагается непарная межтеменная кость (interparietale), представляющая собой остаток дополнительного ряда покровных костей черепа палеозойских амфибий. Иногда она сохраняет свою независимость (например, у многих грызунов — мышей (Muridae), тушканчиков (Dipodidae)), но чаще срастается с теменными костями (у жвачных) или с верхнезатылочной костью (хищные, приматы).
Височная впадина (fossa temporalis) — отверстие в щечном отделе крыши черепа — имеется уже у пеликозавров. Исходно она располагается довольно низко, так что над ней смыкаются заглазничная и чешуйчатая кости. Функция височного отверстия неясна. Похоже, что кость исчезает вблизи центра зоны прикрепления мышцы и сохраняется по ее краям. Тогда центральные мышечные волокна прикрепляются к листку плотной соединительной ткани, закрывающему впадину. Впадины, конечно, облегчают череп, но это может иметь значение, по-видимому, только для крупных форм, у которых они хорошо развиты. У прогрессивных зверообразных височная впадина обычно сильно расширяется в направлении верхней части черепа. Отверстия обеих сторон в некоторых случаях почти встречаются наверху, так что между ними остается лишь узкая полоска прежней теменной поверхности.
На общую форму черепа значительное влияние оказывает наличие дополнительных костных гребней. Так, вдоль поверхности лобных и теменных костей тянутся 2 гребешка, или височных линии (linea temporales), начинающиеся на вершинах верхних или лобных заглазничных отростков. В некоторых случаях они выражены очень слабо. Нередко (у многих насекомоядных, грызунов, приматов, парнопалых) лобно-теменные гребешки раздельно и почти параллельно достигают заднего края теменной поверхности, где примыкают к затылочному, или ламбдовидному, гребню (crista occipitalis), идущему поперек верхнего края затылка и служащему для прикрепления шейных мускулов. Порой височные линии на большем или меньшем расстоянии от надглазничных отростков сближаются и сливаются в срединный продольный стреловидный, или сагиттальный, гребень (crista sagittalis), часто достигающий значительной высоты; позади он раздваивается и сливается с соответственно повышенным поперечным затылочным гребнем (crista occipitalis).
Сверху, сзади и отчасти спереди височные линии ограничивают височную впадину, на поверхности которой начинается очень сильная жевательная мышца, поднимающая нижнюю челюсть и прижимающая ее к верхней челюсти, — височная (musculus temporalis). Гребни развиваются в результате тяги к местам прикрепления фасций, покрывающих данную мышцу; эта тяга усиливается при ее сокращении и утолщении. Передвижение гребешков наверх, к средней линии черепа, и слияние их в стреловидный гребень увеличивают площадь начала мышцы и тем самым делают возможным более мощное ее развитие. Отсюда понятно, что высокие черепные гребни чаще встречаются у хищных форм, у которых сильное сжатие челюстей происходит как при овладении добычей, так и при разрезании ее мышц, сухожилий и дроблении костей. Кроме того, при одинаковом характере пищи гребни должны быть выше у млекопитающих с относительно небольшой мозговой коробкой. Поэтому самые высокие гребни находят у древних представителей класса с маленьким головным мозгом: у диноцерат (Dinocerata), крупных гиенодонтов (Hyaenodontidae), эндрюсарха (Andrewsarchus mongoliensis), энтелодонтов (Entelodontidae) и др. Обратное влияние оказывают небольшие размеры животных, у которых поверхность относительно большой мозговой коробки обеспечивает значительную мощность височной мыщцы без дополнительного увеличения площади ее начала. По заднему краю черепной крыши млекопитающих может формироваться поперечный выйный гребень (crista nuchalis) для мышц и связок, поддерживающих голову.
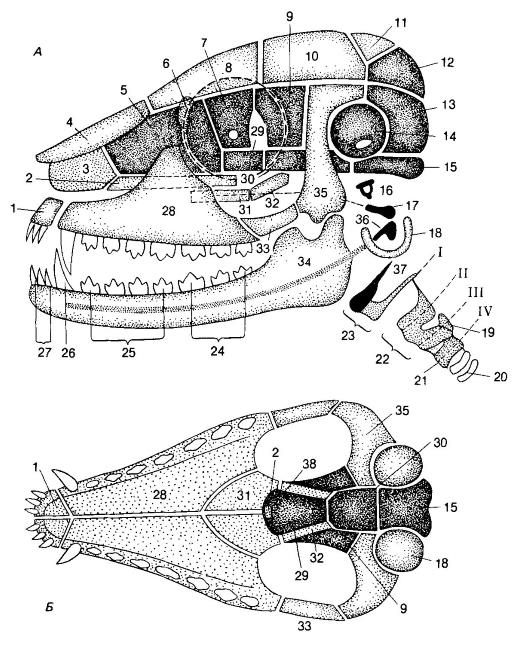
Схема строения черепа с нижней челюстью млекопитающего: А — вид сбоку; Б — вид снизу. Замещающие кости и остатки хрящевого зачаточного черепа показаны темным, покровные кости — светлым. Замещающие кости нейрокраниума: решетчатая (5), глазоклиновидная (7), крылоклиновидная (9), переднеклиновидная (29), основная клиновидная (30), каменистая (14), верхнезатылочная (12), боковая затылочная (13), основная затылочная (15). Покровные кости нейрокраниума: носовая (4), слезная (6), лобная (8), теменная (10), межтеменная (11), чешуйчатая (35), сошник (2). Висцеральный череп: небноквадратный хрящ полностью редуцируется, и на его месте (показано штриховой линией) находятся покровные небная (31) и крыловидная (32) кости; квадратная кость = наковальня (17). Меккелев хрящ также редуцируется; сочленовная кость = молоточек (36); зубная (34) образует вторичный челюстной сустав с чешуйчатой. Межчелюстная (1) и верхнечелюстная (28) образуют верхнюю челюсть, которая скуловой костью (33) связана с чешуйчатой. Из элементов подъязычной дуги образуются стремечко = столбик = гиомандибулярная кость (16) и передние рожки подъязычной кости (37). I-я пара жаберных дуг (I) образует задние рожки подъязычной кости, прирастающие вместе с передними рожками к телу этой кости (23). II-я пара жаберных дуг (II) образует переднюю часть щитовидного хряща (22) гортани, III-я (III) — заднюю часть этого хряща и черпаловидный хрящ (19) гортани, а IV-я (IV) — перстневидный хрящ (21) гортани; 20 — хрящевые кольца трахеи; 38 — вторичные хоаны. Барабанная кость (18) защищает наружный слуховой проход, образуя также раму, на которой натянута барабанная перепонка; 24 — коренные; 25 — предкоренные; 26 — клыки; 27 — резцы.
У основания мозгового черепа кпереди от затылочной кости располагается широко распространенная у позвоночных основная клиновидная кость (basisphenoideum). Впереди от нее находится узкая переднеклиновидная кость (praesphenoideum), которая, вероятно, является лишь обособившейся частью того же первоначального элемента (у рептилий ее гомолога нет). К бокам этих непарных костей прирастают парные основные крылоклиновидные (alisphenoideum) и глазоклиновидные (orbitosphenoideum) кости. Крылоклиновидная кость имеет вид узкой вертикальной пластинки, которая продолжается кверху в виде крыла, огибая сбоку мозговую коробку, и наверху встречается с кроющими костями. Данный элемент, очевидно, тождествен с эпиптеригоидом (epipterygoideum) зверообразных. Первоначально это была пластинка, подымавшаяся с крыловидной кости сбоку от основной клиновидной кости, но лежащая вне мозговой коробки. С разрастанием головного мозга она слилась с черепной коробкой, но все еще соединяется с крыловидной костью. Впереди алисфеноида в стенке глазницы находится глазоклиновидная кость (orbitosphenoideum). Это окостенение сравнительно недавнего происхождения, по-видимому, уже существовавшее у некоторых зверообразных.
Крылоклиновидные и глазоклиновидные кости образуют межглазничную перегородку и передненижнюю часть мозговой коробки, а также формируют крылья клиновидных костей. От основной клиновидной кости отходят большие, или височные, крылья (alae alisphenoidea), а от переднеклиновидной — малые, или глазничные, крылья (alae orbitosphenoidea); в совокупности они образуют нижнюю часть стенки глазницы. На внутренней поверхности основной клиновидной кости присутствует характерное углубление — турецкое седло (sella turcica), вмещающее гипофиз. Во многих случаях слияние основной клиновидной кости с соседними крылоклиновидной, крыловидной, переднеклиновидной и глазоклиновидной костями дает сложный составный (хотя и некрупный) элемент, называемый клиновидной костью (sphenoideum).
У некоторых млекопитающих, включая различные примитивные формы и оба главных отряда копытных, переднеклиновидная кость является самой передней частью мозговой коробки, однако в других отрядах, в частности у приматов, грызунов и хищных, перед ней находится новое окостенение — срединная решетчатая кость (ethmoidale). Когда присутствуют обе кости, переднеклиновидная занимает лишь переднюю часть дна мозговой коробки, входя в состав крыши хоан и (в качестве своей глазоклиновидной части) стенки глазницы, спереди от крылоклиновидной. Ниже переднего конца мозговой коробки впереди от переднеклиновидной кости лежит вторично непарный сошник (vomer) — длинная тонкая срединная кость кожного происхождения, принимающая участие в образовании носовой перегородки и прикрытая снизу вторичным костным небом. Предсошник (praevomer), представляющий собой важный элемент неба рептилий, среди современных млекопитающих имеется лишь у однопроходных. Парасфеноид (parasphenoideum), прикрывавший дно мозговой коробки у нижестоящих форм, у млекопитающих как отдельное окостенение исчез (зачаток наблюдается лишь у зародышей); представлен он или нет в составе клиновидной кости — вопрос спорный.
В зависимости от степени развития зрения глазница имеет большие или меньшие размеры. Большей частью отверстия глазниц направлены в стороны; иногда, при бинокулярном зрении (у приматов и ряда хищных), они повернуты вперед. Перемычка, отделяющая глазницу от височной впадины, утрачивается у некоторых высокоразвитых зверообразных и отсутствует у примитивных млекопитающих; ее верхний конец иногда сохраняется в виде отростка лобной кости. У некоторых групп млекопитающих перемычка по заднему краю глазницы в дальнейшем восстанавливается. Иногда она образована спускающимся вниз заглазничным отростком лобной кости (вepxним) и поднимающимся ему навстречу отростком скуловой кости (нижним заглазничным), причем между их вершинами натянута крепкая связка; в других случаях оба отростка удлиняются и срастаются в костную заглазничную дугу. Эта дуга предохраняет глаз от давления со стороны лежащих позади жевательных мышц. Наиболее совершенна указанная изоляция у высших приматов, у которых глазницу сзади замыкает почти сплошная, глубоко заходящая костная стенка (остается лишь небольшая щель), образованная разрастанием глазничных крыльев передней клиновидной кости.
У переднего угла каждой глазницы помещается небольшая слезная кость (lacrimale). Ее уменьшение произошло еще у зверообразных и было вызвано разрастанием вверх верхнечелюстной кости, несущей сильные клыки. Передняя часть слезной кости выходит на лицевую поверхность, задняя — в полость глазницы. На глазничной поверхности кости, вблизи края глазницы, находятся 1-2 слезных отверстия — протоки, ведущие в слезно-носовой канал (canalis nasolacrimalis), который служит для отведения слез из глазницы в носовую полость. В зависимости от ряда причин, многие из которых неясны, размеры и форма обеих частей слезных костей, а также положение и число слезных протоков у разных форм весьма различается. Иногда слезная кость совсем отсутствует (у однопроходных, ластоногих и некоторых других), порой на ней нет слезных отверстий (у китообразных, сирен, хоботных). В некоторых случаях (у парнопалых, лошадей (Equidae)) она имеет на лицевой поверхности глубокую впадину (т. н. предглазничную) для помещения железы; иногда у ее верхнего края имеется этмоидальная щель, ведущая в носовую полость (олени (Cervidae), жирафы (Giraffidae) и др.). Благодаря такой вариативности строения слезная кость имеет довольно существенное значение для систематики.
Значительные и разнообразные изменения претерпевает в процессе эволюции костная основа слухового аппарата. Изначально она представляет собой хрящевые капсулы, расположенные с двух сторон затылочной области. Отдельные окостенения здесь рано срастаются в парную (правую и левую) каменистую кость (petrosum) — вместилище лабиринта внутреннего уха и внутреннюю стенку барабанной полости среднего уха. Увеличенный головной мозг млекопитающих оттесняет каменистую кость на нижнюю поверхность черепа. Заднее и наружное окостенение стенки слуховой капсулы образует сосцевидная кость (mastoideum), лежащая позади каменистой. В дальнейшем эти кости тесно срастаются в одну каменисто-сосцевидную, или околоушную, кость (perioticum). Гомология околоушного блока млекопитающих ушным костям других позвоночных отрицается.
У разных форм сосцевидный отдел околоушной кости развит различно: иногда он выходит на наружную поверхность черепа узкой полоской между яремным (processus jugularis) и заслуховым (processus posttympanicus) отростками; в других случаях он почти или совсем не виден снаружи, и указанные отростки плотно прилегают друг к другу, или же, напротив, вздувается в большой пузырь, выходя иногда даже на верхнюю поверхность черепа (у некоторых мокроносых приматов, тушканчиков (Dipodidae), шиншилл (Chinchillidae), долгоногов (Pedetidae) и др.). У многих хищных и у человека данный отдел дифференцируется в сосцевидный отросток (processus mastoideus); к нему сзади и медиально часто прилегает околозатылочный отросток (processus paroccipitalis), образованный латерально вытянутой боковой затылочной костью. Иногда околоушная кость слабо связана с окружающими костями черепа — чаще у древних или примитивных форм, как, например, насекомоядных, рукокрылых, ксенартр и некоторых других. У китообразных она соединяется с черепом только связками. В других случаях — у большинства хищных, парнопалых и ряда других околоушная кость тесно сращена с соседними костями — основной затылочной (basioccipitale), боковыми затылочными (exoccipitalia), основной клиновидной (basisphenoideum) и ее крыльями (alisphenoidea).
С каменистой частью околоушной кости соприкасается расположенная книзу от нее барабанная кость (tympanicum), которая ограничивает среднее ухо снаружи. Эта кость свойственна исключительно млекопитающим и, по-видимому, является производным покровной угловой кости (angulare) нижней челюсти. Первоначально барабанная кость имеет форму полукольца или кольца, окружающего наружное слуховое отверстие; на ней натянута барабанная перепонка, воспринимающая звуковые волны. Такой она и остается у более примитивных форм (однопроходных, многих сумчатых и насекомоядных, рукокрылых, некоторых ксенартр, креодонтов и др.). Зачастую снаружи от перепонки барабанная кость вытягивается в слуховой канал, который может удлиняться срастающимися между собой отростками черепа — заслуховым и засуставным (processus postglenoideus). Наряду с барабанной, боковую часть слухового отдела образует другая покровная кость — чешуйчатая (squamosum). Внутри барабанная и чешуйчатая кости нередко плотно срастаются с околоушной костью. Вместе с выступами соседних костей основания черепа они окружают барабанную полость, заключающую в себе 3 крошечные слуховые косточки.
У примитивных современных млекопитающих (например, у большинства сумчатых) барабанная полость не имеет костной защиты от внешних повреждений и прикрыта только лежащей снизу перепонкой; у многих древних форм она также открыта. В процессе эволюции низ данной полости прикрывается костной пластинкой — выростом барабанной кости. Эта пластинка нередко вздувается, образуя костный слуховой пузырь, или барабан (bulla auditiva, или bulla tympani), иногда весьма крупный. В ряде случаев нижняя стенка слухового пузыря образована барабанной костью лишь в своей передней части, а в ее задней части развивается особая внутренняя барабанная кость (entotympanicum) (лептиктиды, ксенартры, некоторые насекомоядные, прыгунчики, даманы, сирены, тупайи, рукокрылые, панголины, хищные, тапиры и носороги). Помимо нее, слуховой пузырь может быть образован находящейся снаружи внешней барабанной костью (ectotympanicum) (шерстокрылы, грызуны, зайцеобразные, различные копытные). Каменистый отдел околоушной кости является основным элементом слухового пузыря относительно редко (приматы, некоторые ежи (Erinaceidae), тенреки (Tenrecidae) и прыгунчики (Macroscelididae); у других ежей, тенреков и прыгунчиков встречаются иные варианты строения, в том числе уникальные). Таким образом, пузырь может состоять из нескольких отдельных, но сливающихся у взрослых особей элементов.
Воздушные полости слуховых пузырей и соединенные с ними полости, образующиеся в соседних костях (включая порой даже небные и крыловидные), служат усиливающими звук резонаторами. Сильнее всего вздуты слуховые пузыри и развиты воздухоносные полости у пустынных животных — лисиц, тушканчиков и др. Околоушная кость и слуховой пузырь у некоторых форм остаются раздельными, а у других (например, у человека) сливаются друг с другом и прилежащей чешуйчатой костью крыши черепа в одну очень сложную височную кость (temporale).
Таким образом, развитие костного слухового аппарата млекопитающих идет в направлении усиления связанности, компактности составляющих его элементов, прочности их срастания с черепом, развития костного среднего уха, предохраняющего нежные слуховые косточки от повреждения, и резонирующих приспособлений. Указанное общее направление эволюции видоизменяется в связи с образом жизни животных, той или иной степенью развития их слуха и по другим причинам; при этом все элементы костного слухового аппарата приобретают разную форму, величину и связь между собой и с другими костями. В частности, очень различно у разных форм строение слуховых пузырей — их величина, форма, состав и степень вздутия. Иногда развитие приостанавливается и сохраняется примитивный характер всей области. Разнообразие в строении костного слухового аппарата у млекопитающих делает его важным систематическим признаком.
Слуховой пузырь прободен наружным слуховым проходом (mеаtus acusticus externus), ведущим снаружи к барабанной перепонке, а также отверстием на своем переднем конце для слуховой, или евстахиевой, трубы, соединяющей среднее ухо с глоткой. Внутренняя сонная артерия, снабжающая кровью головной мозг, первоначально проходила от заднего конца черепа вперед по его нижней стороне мимо среднего уха. У большинства млекопитающих эта область перекрыта барабаном, и артерия тянется вперед под ним по сонному каналу (canalis caroticus) почти до того места, где она входит в мозговую коробку. Рядом с этим местом может находиться алисфеноидный, или крыловой, канал (canalis alaris), пропускающий ветвь данной артерии вперед к небу.
Зрительный нерв обычно входит в полость черепа через зрительное отверстие (foramen opticum) в глазоклиновидной кости. Перед крылоклиновидной костью внутри глазничной полости находится крупное переднее рваное отверстие (foramen lacerum anterius) — клиновидная, или глазничная, щель. Круглое (foramen rotundum) и овальное (foramen ovale) отверстия обычно пронизывают саму крылоклиновидную кость, а у ее заднего края лежит среднее рваное отверстие (foramen lacerum medium), с которым может сливаться овальное. Обычно нервы глазных мышц и самая передняя, глазная ветвь тройничного нерва проходят через переднее рваное отверстие, верхнечелюстная и нижнечелюстная ветви тройничного нерва — через круглое и овальное отверстия. Среднее рваное отверстие не содержит главных нервов, но именно через него или рядом с ним в мозговую коробку входит внутренняя сонная артерия.
VII-й и VIII-й нервы проходят в слуховую капсулу из полости черепа через внутренний слуховой проход (meatus acusticus internus) в каменистой части височной кости. Первый из них первоначально выходил на поверхность у передней оконечности слуховой капсулы, однако у млекопитающих в связи со сложным строением среднего уха он выходит позади слухового пузыря через шиловидно-сосцевидное отверстие (foramen stylomastoideum). Здесь же в промежутке между слуховой капсулой и основной затылочной костью находится заднее рваное (foramen lacerum posterius), или яремное (foramen jugulare), отверстие, через которое проходят IX-й, Х-й и XI-й головные нервы и внутренняя яремная вена, собирающая кровь от головы. XII-й нерв обычно проходит через отдельное, иногда множественное, подъязычное отверстие (foramen hypoglossi) в дне затылка. Первоначально он выходил через переднее мыщелковое отверстие (foramen condyloideum anterius) в боковой затылочной кости (exoccipitale).
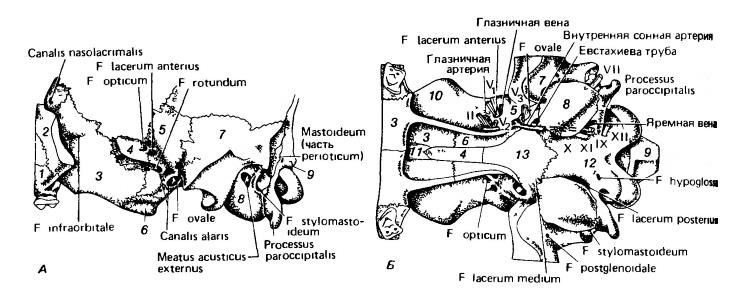
Мозговая коробка волка (Canis lupus), вид сбоку (А) и снизу (Б). Показаны отверстия в основании мозговой коробки. На Б изображены главные нервы, прохождение внутренней сонной артерии, ее небной ветви и яремная вена. 1 — maxillare; 2 — jugale; 3 — palatinum; 4 — praesphenoideum; 5 — alisphenoideum; 6 — pterygoideum; 7 — squamosum; 8 — слуховой барабан; 9 — occipitale; 10 — frontale; 11 — vomer; 12 — basioccipitale; 13 — basisphenoideum.
Сбоку и часто несколько спереди от слуховой области с каждой стороны черепа начинаются уже упомянутые чешуйчатые кости (squamosa). Единственные у млекопитающих из набора щечных костей, они образуют у них значительную часть боковой стенки мозговой коробки. Каждая чешуйчатая кость расположена либо на границе затылочной и теменной части черепа, либо, при прогрессивном развитии мозговой полости и оттеснении слуховой капсулы книзу, составляет нижнебоковые стенки теменной области. Чешуйчатая кость обладает гленоидной суставной ямкой для причленения нижней челюсти. Следует особенно подчеркнуть, что сочленение нижней челюсти непосредственно с чешуйчатой костью составляет одну из наиболее характерных особенностей черепа млекопитающих. Также эта кость несет направленный вперед мощный скуловой отросток (processus zygomaticus ossis squamosi), который вместе со скуловой костью (jugale, или zygomaticum) и скуловым отростком на заднем крае верхнечелюстной кости образует единственную с каждой стороны скуловую дугу (arcus zygomaticus), ограничивающую глазницу и височную область черепа снаружи и снизу.
Скуловая дуга имеет разную форму и величину. На ее нижней и внутренней поверхности начинается вторая мощная мышца — собственно жевательная, или массетерная (musculus masseter), поднимающая нижнюю челюсть и прикрепляющаяся к нижней челюсти на наружной поверхности ее углового отдела. Подобно височной, массетерная мышца сильно развита у хищников, у которых особенно сильное сжатие челюстей имеет место в их заднем отделе, вблизи прикрепления указанной мышцы, где находятся их плотоядные (для разрезания мясной пищи) и задние бугорчатые (для раздавливания плодов и пр. при смешанном режиме) зубы.
Массетерная мышца хорошо развита и у некоторых других групп, например у свиней (Suidae) (работа коренных при дроблении плодов, корней и пр.). В этом случае скуловая дуга не только широкая (в вертикальном направлении), но и выпуклая вверх; это, во-первых, предохраняет ее от излома при сокращении мышцы, во-вторых, усиливает ее роль как контрфорса, воспринимающего давление наверх со стороны задних зубов при жевании. При этом скуловые дуги обычно не только выпуклы вверх, но и широко расходятся в стороны: внутри от них должно быть достаточно места для утолщающихся при сокращении мощных височной и массетерной мышц.
В тех случаях, когда размельчение пищи происходит путем не разрезания или раздавливания, а посредством перетирания, — переднезадними движениями (слоны (Elephantidae), многие грызуны) или боковыми (жвачные), или же размельчение пищи не требует большого усилия и сильного сдавливания челюстей (насекомоядные, рукокрылые), или, наконец, пища направляется в желудок почти без предварительного размельчения (ластоногие, китообразные, многие ксенартры), — скуловые дуги становятся тонкими, прямыми, прижатыми к черепу, неполными, а иногда и совсем исчезают (ехидны (Tachyglossidae), землеройки (Soricidae), муравьеды (Myrmecophagidae)). При этом скуловая дуга теряет свое значение как места начала задних порций массетерной мышцы, так и контрфорса: давление нижней челюсти на верхнюю уже не концентрируется в их задних отделах, а распределяется по ряду щечных зубов равномерно; кроме того, зубные ряды в этих случаях обычно перемещены к средней линии черепа, основание которого и принимает на себя давление при жевании.
Характер жевательных движений вызывают видоизменение и других черепных элементов. При переднезадних движениях тяга вперед совершается преимущественно некоторыми частями массетерной мышцы. У грызунов, у которых преобладает именно такой способ размельчения пищи, поверхностный слой массетерной мышцы перемещается на лицевую сторону верхнечелюстной кости, впереди глазницы, где для его прикрепления развиваются специальные площадки и выступы. Кроме того, иногда передняя порция внутреннего слоя той же мышцы проходит через сильно увеличенное подглазничное отверстие (foramen infraorbitale) на лицевую поверхность, где прикрепляется на верхнечелюстной и даже на межчелюстной костях. Движение челюсти назад производится двубрюшной мышцей (musculus digastricus) от яремного отростка черепа к нижней и внутренней стороне нижней челюсти. При боковых перетирающих движениях попеременно работают крыловидные мышцы (наружная и внутренняя) от крыловидного отростка основной и от крыловидной костей к внутренней поверхности нижней челюсти. В этих случаях крыловидные выступы вырастают в высокие, иногда раздвоенные пластины.
В свете вышеизложенного интересно проследить путь формирования черепной коробки высших млекопитающих. У их далеких предков теменные и лобные кости формировали над мозговым отделом широкую крышу. С развитием височных впадин, их поверхность сильно уменьшилась. Однако эти кости имели тенденцию к образованию выступов, направленных вниз вдоль стенок полости черепа под челюстными мышцами. У млекопитающих при сильном увеличении мозга эти выступы сильно разрослись, распространившись над вздувшейся поверхностью мозга в стороны и образовав значительную часть крыши и боковых стенок мозговой коробки. Боковая кость покровной крыши — чешуйчатая — тоже участвует в образовании вторичного покрытия мозга.
Все эти новые продолжения элементов покровной крыши выглядят как части первичной поверхности черепа, однако на самом деле ими не являются. Они лежат глубже височных мышц, смыкающих челюсти, а первичная крыша черепа находилась от челюстных мышц снаружи. Первичную поверхность черепа представляет только та часть лобных и теменных костей, которая лежит между височными линиями или которая образует стреловидный гребень; остальная их поверхность — новое, глубже залегающее разрастание. Если не вдумываться, можно счесть скуловые дуги внешним атрибутом черепа как такового, вроде ручек у кувшина. Однако все обстоит как раз наоборот. Дуги — последние остатки прежних боковых стенок черепа, а цельная мозговая коробка — это в основном новообразование, лежащее глубже челюстных мышц.
Чтобы заполнить «брешь» в стенке разрастающейся мозговой коробки, добавился еще один элемент: в виде крылоклиновидной кости в нее была включена верхнекрыловидная кость. У рептилий мозговая коробка между глазницами не окостеневает, а верхнекрыловидная кость напоминает вертикальный стержень, который как часть верхней челюсти лежит снаружи от этой неокостеневшей зоны. У зверообразных данный стержень расширяется до пластинки, по-прежнему находящейся снаружи от мозговой коробки, а у млекопитающих эта кость включилась в ее стенку, заполнив брешь над основной клиновидной костью, и так прочно срослась с последней, что (как подразумевает новое название) внешне выглядит крылом составного клиновидного элемента.
В результате костная капсула, окружающая мозг млекопитающих, представляет собой весьма сложное образование. Исходная мозговая коробка формирует в основном ее дно, а покровные элементы и крылоклиновидная кость участвуют в построении стенок и крыши этой разросшейся конструкции.
Изображения черепов зверообразных (пеликозавр диметродон (Dimetrodon), горгонопс сцимногнат (Scymnognathus), тероцефал баурия (Bauria)) и млекопитающего (плацентарное волк (Canis)); вид сверху, сбоку и снизу.
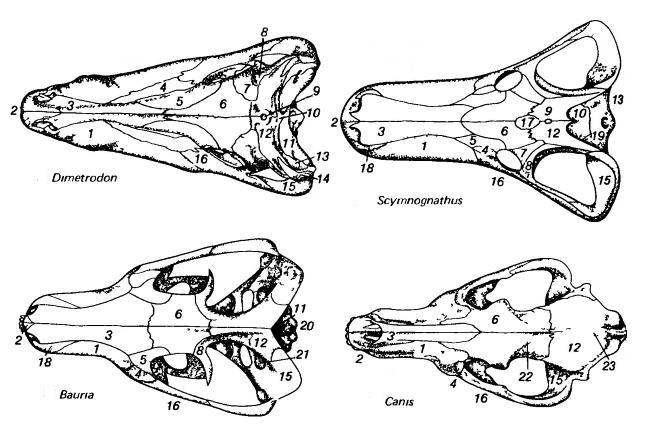
1 — maxillare; 2 — premaxillare; 3 — nasale; 4 — lacrimale; 5 — prefrontale; 6 — frontale; 7 — postfrontale; 8 — postorbitale; 9 — теменное отверстие; 10 — posparietale; 11 — supraoccipitale; 12 — parietale; 13 — tabulare; 14 — supratemporale; 15 — squamosum; 16 — jugale; 17 — preparietale; 18 — septomaxillare; 19 — occipitale; 20 — exoccipitale; 21 — prooticum; 22 — височная линия; 23 — сагиттальный гребень.
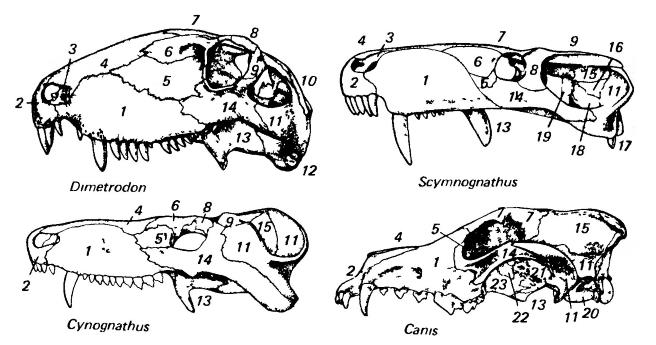
1 — maxillare; 2 — premaxillare; 3 — septomaxillare; 4 — nasale; 5 — lacrimale; 6 — prefrontale; 7 — frontale; 8 — postfrontale; 9 — postorbitale; 10 — supratemporale; 11 — squamosum; 12 — quadratum; 13 — pterygoideum; 14 — jugale; 15 — parietale; 16 — posparietale; 17 — quadratojugale; 18 — supraoccipitale; 19 — epipterygoideum; 20 — слуховой барабан; 21 — alisphenoideum; 22 — presphenoideum; 23 — palatinum.
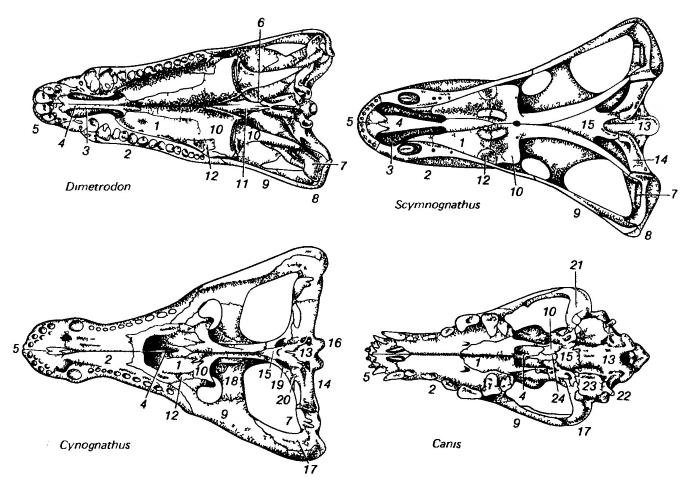
1 — palatinum; 2 — maxillare; 3 — внутренняя ноздря; 4 — vomer; 5 — premaxillare; 6 — базальное сочленение неба и мозговой коробки; 7 — quadratum; 8 — quadratojugale; 9 — jugale; 10 — pterygoideum; 11 — parasphenoideum; 12 — ectopterygoideum; 13 — basioccipitale; 14 — opisthoticum; 15 — basisphenoideum; 16 — exoccipitale; 17 — squamosum; 18 — postorbitale; 19 — epipterygoideum; 20 — prooticum; 21 — сочленовная ямка; 22 — околозатылочный отросток; 23 — слуховой барабан; 24 — presphenoideum.