в оглавление
назад
Скелетная система
Подобно другим тетраподам, млекопитающие обладают прочным внутренним скелетом. Он выполняет опорную функцию, защищает многие внутренние органы и служит субстратом для прикрепления большинства поперечнополосатых мышц, совершая с ними движения тела. В состав скелета входит череп, позвоночный столб, ребра, пояса конечностей и свободные конечности. В соответствии с широким спектром используемых способов передвижения и добывания пищи одни и те же части скелета у различных видов отличаются большим разнообразием строения. По сравнению с рептилиями общее количество костных элементов в скелете млекопитающих сокращено.

Скелеты эдафозавра (Edaphosaurus sp.) (I), тупайи (Tupaia glis) (II), кролика (Oryctolagus cuniculus) (III) и человека (Homo sapiens) (IV), врисованные в контуры тела: 1 — череп; 2 — позвоночник; 3 — ребра; 4 — грудина; 5 — лопатка; 6 — коракоид; 7 — ключица; 8 — плечевая кость; 9 — лучевая кость; 10 — локтевая кость; 11 — запястье; 12 — пясть; 13 — фаланги пальцев; 14 — таз; 15 — бедренная кость; 16 — большая берцовая кость; 17 — малая берцовая кость; 18 — предплюсна; 19 — плюсна.
С физиологической точки зрения скелет может показаться довольно инертной системой. Однако подобное впечатление обманчиво, поскольку не только клетки скелетных тканей, но и окружающее их «мертвое» межклеточное вещество является ареной беспрестанной биохимической активности. В течение всей жизни скелетные элементы постоянно преобразуются — как в отношении внутренней структуры, так и по внешней форме — в соответствии с механическими нагрузками натяжения и сжатия.
Ткани скелета
Скелет млекопитающих преимущественно костный, доля хрящевой ткани в нем сравнительно мала. Хотя хрящ и кость являются специализированными производными соединительных тканей и возникают из мезенхимы, они существенно различаются по природе, по способу образования и по положению.
Хрящ представляет собой полупрозрачный стекловидный материал, гибкий и довольно эластичный. Его основное вещество, или матрикс — это главным образом сульфатированный полисахарид (хондромукопротеин), который образует плотный гель, содержащий сеть соединительнотканных волокон. В разбросанных полостях, лакунах, помещаются хрящевые клетки — хондроциты. Обычно они имеют округлую форму и лишены ветвящихся отростков, характерных для костных клеток. Хондроциты изолированы внутри матрикса, который сами же секретировали. Кровеносные сосуды в хряще практически отсутствуют; вследствие этого питательные вещества, необходимые клеткам, должны достигать их путем диффузии через основное вещество. Наружная поверхность хряща покрыта слоем плотной, содержащей клетки соединительной ткани — надхрящницей (perichondrium).
Хрящевой ткани особенно много у зародышей млекопитающих. У детенышей она присутствует в зонах костей, продолжающих свой рост. У взрослых особей хрящ сохраняется лишь там, где требуется гибкий и податливый материал — на суставных головках, в местах крепления связок и сухожилий; им образованы грудные отделы ребер, межпозвоночные диски и ушные раковины.

Схема строения гиалинового хряща, покрытого надхрящницей: 1 — волокнистый слой надхрящницы; 2 — клеточный слой надхрящницы; 3 — молодые хондроциты; 4 — хондроцит в лакуне; 5 — межклеточное вещество (хрящевой матрикс); 6 — интерстициальный рост; 7 — изогенные группы хондроцитов (зрелые хрящевые клетки).
Во взрослом состоянии у млекопитающих безоговорочно доминирует кость. Вслед за более древними группами позвоночных продолжается совершенствование ее гистологической структуры.
Основной объем кости сложен из межклеточного вещества, или костного матрикса, — твердого непрозрачного обызвествленого материала, пронизанного сетью коллагеновых волокон. Между ними откладывается гидрооксиапатит (разновидность фосфата кальция) и, в меньшей степени, сульфат и карбонат кальция. От доли минеральных веществ, пропитывающих костную ткань, зависит ее прочность. Наиболее высока она в костях, выдерживающих максимальную механическую нагрузку. Степень минерализации костной ткани различается у разных групп: у китообразных из-за относительной невесомости тела в воде скелет содержит значительно меньше минеральных солей, чем скелет наземных млекопитающих. Особенно мало минерализованы кости молодых животных, отличающиеся гибкостью и мягкостью.
Внутри межклеточного вещества заключены костные клетки — остеоциты, которые его и секретировали. От хрящевых клеток остеоциты заметно отличаются тем, что они (и лакуны в кости, в которых они располагаются) имеют неправильную звездчатую форму. На ранних стадиях развития для остеоцитов характерны длинные ветвящиеся отростки, которые тянутся от лакун во всех направлениях по крошечным канальцам (canaliculi), достигающим соседних лакун. Позже протоплазматические отростки могут втягиваться, но канальцы остаются. Плотный костный матрикс непроницаем для питательных веществ и кислорода, поэтому они доставляются к костным клеткам по кровеносным сосудам.

Схема строения костных клеток: А — строение остеобласта: 1 — ядро; 2 — цитоплазма; 3 — остеоид; 4 — развитая зернистая эндоплазматическая сеть; Б — строение остеоцита: 1 — отростки остеоцита; 2 — эндоплазматическая сеть; 3 — ядро; 4 — внутренний сетчатый аппарат; 5 — митохондрия; 6 — остеоидное (необызвествленное) вещество кости по краям лакуны, в которой расположен остеоцит.
Наружная поверхность кости покрыта тонкой, но плотной волокнистой соединительнотканной оболочкой — надкостницей (periosteum). При необходимости ее глубокая часть предоставляет клетки для восстановления поврежденной костной поверхности, формируя параллельные костные пластинки. В зрелой кости надкостница связана с ней пучками шарпеевых волокон, которые внедряются в поверхность кости. Кровеносные сосуды и нервы проходят в надкостнице по узким фолькмановым каналам и ветвятся внутри кости по гаверсовым каналам расположенным под самыми различными углами. На растущих участках костей, слагаемых хрящом, а также на концах трубчатых костей надкостница отсутствует.
Если хрящ обычно выглядит на срезе довольно однородно, то кость имеет сложное внутреннее строение. Многие участки скелетных элементов, особенно их наружные слои, состоят из компактной костной ткани (compacta). Обычно она представляет собой совокупность плотно налегающих друг на друга костных пластинок (lamellae). Наиболее отчетливо такое строение выражено у трубчатых костей конечностей, несущих наибольшую нагрузку. Их стенки построены из остеонов — вложенных одна в другую костных трубочек, скрепленных плотными пучками коллагеновых фибрилл. Между ними располагаются костные клетки, а во внутренней полости такого остеона, в гаверсовом канале, проходят нерв и кровеносный сосуд. Промежутки между остеонами заполнены вставочными костными пластинками. Благодаря подобному строению кость отличается относительно небольшим весом и большой прочностью.
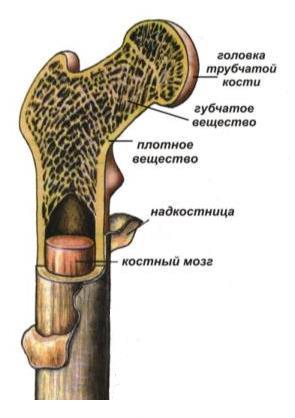
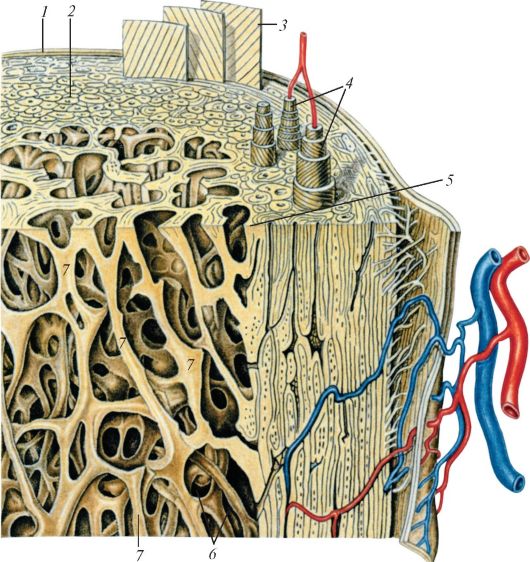
Слева — схема строения трубчатой кости, справа — то же в увеличении: 1 — надкостница; 2 — компактное вещество кости; 3 — слой наружных окружающих пластинок; 4 — остеоны; 5 — слой внутренних окружающих пластинок; 6 — костномозговая полость; 7 — костные балочки губчатой кости.
Сердцевина коротких и плоских костей, а также суставные концы трубчатых костей построены из губчатой костной ткани (spongiosa). Она содержит каркас из костных балочек, или трабекул. Балочки ориентированы вдоль линий напряжения в кости, что обеспечивает оптимальную прочность при имеющемся количестве материала. После повреждения, например перелома кости, с последующим неправильным срастанием вся система костных перемычек в пострадавшей области постепенно переориентируется так, чтобы наилучшим образом служить при нагрузках. Таким образом, с помощью губчатой костной ткани формируется крепкая и притом легкая и экономичная конструкция, выдерживающая значительные нагрузки.

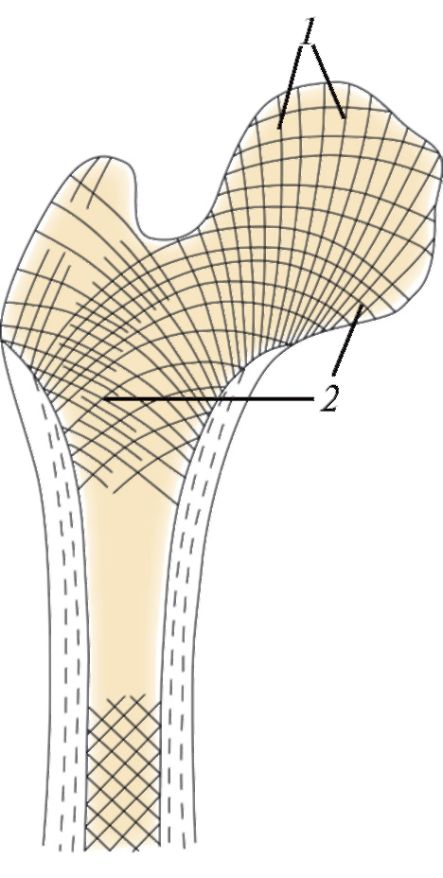
Слева — схема строения остеона: 1 — центральный канал остеона; 2 — пластинки остеона; 3 — костная клетка (остеоцит).
Справа — расположение костных балочек в губчатом веществе вдоль линий напряжения: 1 — линии сжатия (давления); 2 — линии растяжения.
Основной объем губчатой кости занят красным костным мозгом, вырабатывающим кровяные клетки. В полостях, расположенных в средних частях трубчатых костей, содержится желтый костный мозг. Он представляет собой депо жира. После больших кровопотерь на месте желтого мозга может воссоздаваться красный костный мозг.
О кости привычно думать как о статичном, неактивном материале. Это представление далеко от истины. На протяжении жизни кости непрерывно перестраиваются путем резорбции старого и отложения нового костного материала. В местах резорбции находятся большие многоядерные клетки — остеокласты. Они возникают из моноцитов и участвуют в рассасывании кости. После разрушение костного материала в данном месте формируются гаверсовы каналы, и кость заново откладывается концентрическими слоями в толще их стенок. Остатки ранее сформированных костных слоев, не разрушенных при образовании ныне существующих гаверсовых систем, образуют промежуточную систему. Кроме того, кости служат главными депо кальция и фосфора (в виде фосфата) — элементов, играющих важнейшую роль в метаболических процессах организма.

Тонкий поперечный срез через пястный элемент млекопитающего.
Костная ткань возникает благодаря активности мезенхимных клеток, которые становятся остеобластами и затем формируют вокруг самих себя толстый слой матрикса, содержащий множество волокон. Быстрое отложение солей кальция в этом межклеточном материале завершает формирование кости, и остеобласты, выполнив свою главную функцию, становятся остеоцитами. Кости млекопитающих в основном замещающие, или эндохондральные. В гораздо меньшей степени представлены покровные и мембранные кости.
Формирование эндохондральной кости выражается в постепенном замещении зародышевого хряща характерным для взрослого животного костным органом. При этом важно учитывать, что значительная часть костного материала в действительности с самого начала откладывается снаружи от зародышевого хряща. В длинных костях конечностей хрящ принимает окончательную форму взрослой кости на ранней стадии развития, когда он имеет еще чрезвычайно малые размеры. Вскоре хрящ начинает претерпевать изменения и перерождение в области середины своей длины: хондроциты набухают и выстраиваются в продольные столбики, материал между этими столбиками обызвествляется, а хрящевые клетки погибают. Кровеносные сосуды внедряются в хрящ с поверхности, и происходит разрушение хряща в этой центральной области. Остеобласты, проникающие с кровеносными сосудами, откладывают кость на месте разрушенного хряща. Окостенение распространяется от центра в направлении обоих концов элемента.

Срез элемента метаподия зародыша млекопитающего, в котором происходит окостенение стержня (кость показана черным). На поверхности происходит перихондральное окостенение. На обоих концах находится нормальный хрящ; ближе к середине стержня хрящ становится «гипертрофированным» (клетки раздуваются и выстраиваются в ряды), происходит обызвествление, за которым следует замещение хряща эндохондральной костью.
Если бы хрящ не рос, его замещение было бы завершено в короткое время, и получился бы полностью окостеневший элемент крошечного размера. Но этого не происходит, потому что хрящ растет с обоих концов примерно с той же скоростью, что и разрушается в центре. Хрящ, так сказать, долго ведет за собой «гонящиеся по пятам» остеобласты; в некоторых случаях процесс окостенения так никогда и не нагоняет хрящ на обоих концах элемента. При полном окостенении рост прекращается, потому что своими концами внутренние скелетные элементы обычно сочленяются с соседними, а добавить кость непосредственно на сочленовные поверхности без вреда для суставов нельзя.
У низших позвоночных внутренние элементы обычно окостеневают из одного центра — например, стержневого отдела длинной кости, или диафиза (diaphysis), а ее сочленовные концы, или эпифизы (epiphyses), часто даже во взрослом состоянии в значительной степени остаются хрящевыми. Но у млекопитающих и птиц (и значительно реже у рептилий) имеются добавочные центры окостенения. Обычно они развиваются на эпифизах, а также на выступающих отростках для прикрепления мышц. Здесь появляются центры окостенения, подобные центру диафиза, но меньшего размера. Эти центры могут превратить эпифизы в кость задолго до того, как рост диафиза закончился и, таким образом, несмотря на неполное окостенение, позволяют костному элементу полноценно функционировать. Между эпифизом и диафизом долго сохраняется прослойка хряща. Это зона роста; здесь хрящ непрерывно растет и замещается костью со стороны и диафиза, и эпифиза. Таким образом, удлинение кости может совершаться, не затрагивая ее сочленений. Стоит окостенению уничтожить срединную прослойку хряща, как диафиз и эпифиз объединяются — рост кости заканчивается и ею достигается окончательный взрослый размер. В этом отношении млекопитающие представляют контраст подавляющему большинству рептилий, у которых предельной величины взрослого животного нет, и рост, по-видимому, может продолжаться всю жизнь.

Окрашенные и просветленные препараты человеческих эмбрионов 14-16-недельного возраста; хорошо заметны темно-красные центры окостенения.
Исчезновение ростовых хрящей происходит у животных одного вида приблизительно в одно и то же время, и это используется человеком на практике. Например, охотоведы узнают возраст зайцев по исчезновению зон роста в костях запястья, что происходит, как правило, в возрасте 1 года.
Хотя значительная часть прироста внутренней — «хрящевой» — кости происходит путем замещения хряща, этим дело не ограничивается. Когда закладывается центр окостенения стержня длинного элемента, последний имеет маленький диаметр. По мере того как хрящ растет с обоих концов, они постепенно утолщаются, приближаясь к окончательному размеру, так что если бы кость формировалась исключительно путем внутреннего замещения, то она напоминала бы по форме песочные часы с суженной средней частью. Для исправления этого дефекта, помимо внутреннего замещения, в «хрящевых» костях происходит значительное приращение за счет непосредственного формирования на поверхности хряща перихондральной кости, образующей следующие друг за другом концентрические слои. Эти слои толще всего в первоначально тонкой средней части стержня. Процесс поверхностного добавления костного материала может продолжаться и после того, как лежащий глубже стержень окостенел; на этой стадии термин периостальная кость подходит лучше, чем перихондральная.

Окостенение и рост длинной кости млекопитающего. Горизонтальной штриховкой обозначены эндохондральные окостенения, вертикальной — перихондральные окостенения, черным — компактная костная ткань, белым по концам кости — хрящ, внутри — полость костного мозга. 1 — хрящевая стадия. 2 — закладка в хряще диафиза губчатой эндохондральной и компактной перихондральной кости. 3 — диафиз окостенел изнутри (эндохондрально), снаружи (перихондрально), эпифизы остаются еще хрящевыми. 4 — первая закладка эндохондрального окостенения в верхнем эпифизе. 5 — эндохондральное окостенение в обоих эпифизах, образование костномозговой полости вследствие резорбции эндохондральной кости и появление компактной костных масс в диафизе. 6 — в связи с дальнейшим окостенением продольный рост происходит только в тонких прослойках активно растущего хряща между диафизом и эпифизами. 7 — в связи с исчезновением хряща между диафизом и эпифизами продольный рост кости прекратился, хрящи остались только на сочленовных поверхностях, костномозговая полость увеличилась в результате резорбции центральной части при продолжении отложения кости на поверхности.
Важно отметить, что скелетные элементы, у которых отсутствует хрящевая стадия, обычно имеют простую пластинчатую форму и могут свободно нарастать с любой стороны, пока не достигнут окончательного размера. В то же время большинство глубоких элементов, которым предшествует хрящ, имеет сложные сочленения с соседними элементами, и к ним прикрепляются важные мышцы. Скелет млекопитающего уже на очень ранних стадиях развития является почти точной, хотя и крошечной, копией скелета взрослого организма, а ранняя прочность суставов и мест мускульных прикреплений дает возможность двигаться уже молодым животным. Если бы внутренние элементы с их сложными суставными связями сразу формировались как кость, то их рост до взрослого размера был бы невозможен, потому что кость может нарастать только снаружи, а поверхностный рост разрушил бы ее связи с окружающими структурами.
Намного проще процесс формирования покровных костей, имеющих кожное происхождение. В этом случае группа мезенхимных клеток в дерме приобретает свойства остеобластов и откладывает тонкие, неправильной формы пленки или пластинки плотного волокнистого межклеточного вещества, в котором вскоре откладываются характерные для кости минеральные соли. Такие пластинки постепенно нарастают с краев и утолщаются путем добавления последующих слоев кости на внутренней и наружной поверхностях. У взрослых особей пластинчатые покровные кости состоят из внутреннего и наружного слоев компактной кости с промежуточным слоем губчатой кости, возникшим в результате перестройки. У млекопитающих покровные кости имеются в черепе, нижней челюсти и плечевом поясе. За редкими исключениями вторичного характера (например, челюстной сустав), они совершенно не содержат хрящевой ткани.

Ранняя стадия развития покровной кости в черепе кошки (Felis silvestris): 1 — коллагеновое межклеточное вещество; 2 — однородные утолщенные коллагеновые волокна, которые становятся межклеточным костным веществом; 3 — клетки соединительной ткани с отростками (эти клетки становятся остеобластами и позже остеоцитами); 4 — фибробласты.
Сходный способ окостенения наблюдается в мембранных костях — более глубоких скелетных элементах, которые являются целиком костными на всех стадиях своего развития. Таковы, к примеру, гетеротопные кости.
Кости и хрящи соединяются друг с другом разными способами. Так, в черепе, где кости непосредственно примыкают друг к другу, движения нежелательны. Такое соединение между элементами, называемое синартрозом (synarthrosis), образовано тонкими промежуточными прослойками хряща или соединительной ткани, а видимая линия соединения называется швом (sutura). Свободно подвижное сочленение между соседними элементами называется диартрозом (diarthrosis). В таком случае контактирующие поверхности костных элементов покрыты слоем хряща, а синовиальная оболочка и плотная соединительная ткань образуют вокруг сустава суставную сумку (капсулу). Сложные утолщения этой капсулы образуют связки. В некоторых случаях суставная полость подразделяется на две части тонкой пластинкой волокнистого хряща — мениском. Суставные головки и ямки построены таким образом, что делают возможным движение только в определенных пределах и направлениях. Наряду с простыми полушаровидными головками в плечевом и тазобедренном суставах можно встретить и цилиндрические суставные поверхности, часто дополненные направляющими килями. Комбинированное движение в двух направлениях возможно при наличии сложных суставных седлообразных соединений, как, например, в запястьях приматов. Густая и эластичная синовиальная жидкость, содержащаяся в суставной сумке, снижает трение до минимума, так же как и пришлифованные друг к другу суставные поверхности.

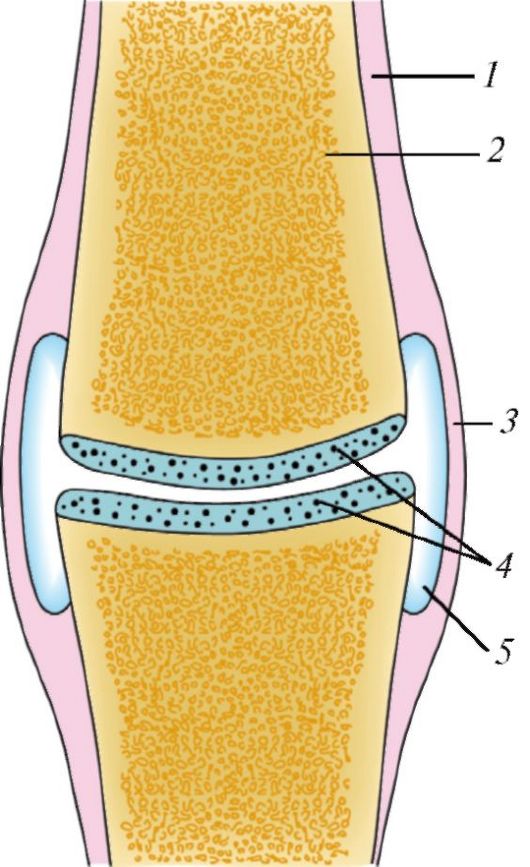
Слева — различные виды непрерывного соединения костей: А — синдесмоз: 1 — надкостница; 2 — кость; 3 — волокнистая соединительная ткань; Б — синхондроз: 1 — надкостница; 2 — кость; 3 — хрящ; В — симфиз (гемиартроз): 1 — надкостница; 2 — кость; 3 — межлобковый диск; 4 — щель в межлобковом диске.
Справа — схема строения диартрозного сустава: 1 — надкостница; 2 — кость; 3 — суставная капсула; 4 — суставной хрящ; 5 — суставная полость.
Скелет млекопитающих включает множество элементов, имеющих различную форму, строение, функцию, положение и эмбриональное происхождение. Остатки покровного скелета, характерного для большинства низших позвоночных, у млекопитающих сливаются с расположенным глубже внутренним скелетом, или эндоскелетом. В последнем можно провести разграничение на две категории неравного объема — соматические и висцеральные элементы. К висцеральному скелету принадлежат слуховые косточки и частично нижняя челюсть — производные скелетных элементов низших позвоночных, располагавшихся между жаберными отверстиями и управлявших движениями стенок глотки. Как будет показано далее, висцеральная область обладает особой мускулатурой и иннервацией. Соматический скелет включает в себя все остальные элементы. При этом большая часть мозговой коробки, позвоночник, ребра и связанные с ними элементы туловища составляют осевой скелет. Скелет конечностей, состоящий из поясов и свободных частей, можно считать производным осевого скелета.
Некоторые структурные единицы или даже одиночные кости скелета взрослых млекопитающих содержат элементы, происходящие из двух или большего числа различных источников. Так, например, плечевой пояс содержит и покровные, и эндоскелетные компоненты; нижняя челюсть и висцеральные, и покровные элементы. Сложнее всего устроен череп, который образован покровными, соматическими и висцеральными элементами.
вперед
в оглавление