в оглавление
назад
Череп (продолжение)
Общая форма черепа нередко значительно изменяется вследствие развития внутри его костей воздушных камер или синусов. При этом губчатая костная ткань, прослойка которой разделяет у молодого животного наружный и внутренний слои плотного костного вещества, исчезает, и оба плотных слоя раздвигаются, оставляя между собой полость. В образовавшейся камере, при достаточной ее величине, развиваются тонкие костные поддерживающие перегородки, делящие камеру на ячеи и расположенные в разных направлениях. Чаще всего полости развиваются в крыше черепа — в лобных, теменных и затылочных костях; но иногда они распространяются в боковые кости мозговой коробки в чешуйчатые, сосцевидные, боковые затылочные, а также в кости основания черепа — решетчатую, основную клиновидную и крыловидные. Нередко синусы связаны с полостями черепа — носовой, барабанной; слизистая оболочка последних при этом врастает в синусы и покрывает их стенки изнутри.
Пневматизация костей черепа свойственна только млекопитающим. О ее значении существуют разные теории. По-видимому, все они частично правы, и развитие костных полостей определяется не одной причиной. Прежде всего, они возникают везде, где для сохранения нужных пропорций, прочности и компактности черепа необходимо утолщение костей; в таких случаях костные полости устраняют значительное утяжеление костей, поэтому у водных животных по понятной причине костных черепных пазух не бывает. У крупных форм при прочих равных условиях костные полости развиты сильнее, чем у близких к ним мелких, что также объясняется стремлением к облегчению костей. Далее, в ходе эволюции отдельных ветвей млекопитающих развитие лобных пазух часто вызывается увеличением полушарий большого мозга, верхнепередняя граница которых, находящаяся у примитивных форм на одном уровне с поверхностью обонятельных лопастей, начинает все более круто спадать к последним, и с поверхности черепа образуется впадина, нарушающая плавность eгo очертаний и eгo крепость. В этом месте плотные слои лобных костей раздвигаются и восстанавливается правильность очертаний черепа, а внутри кости возникает полость или синус.
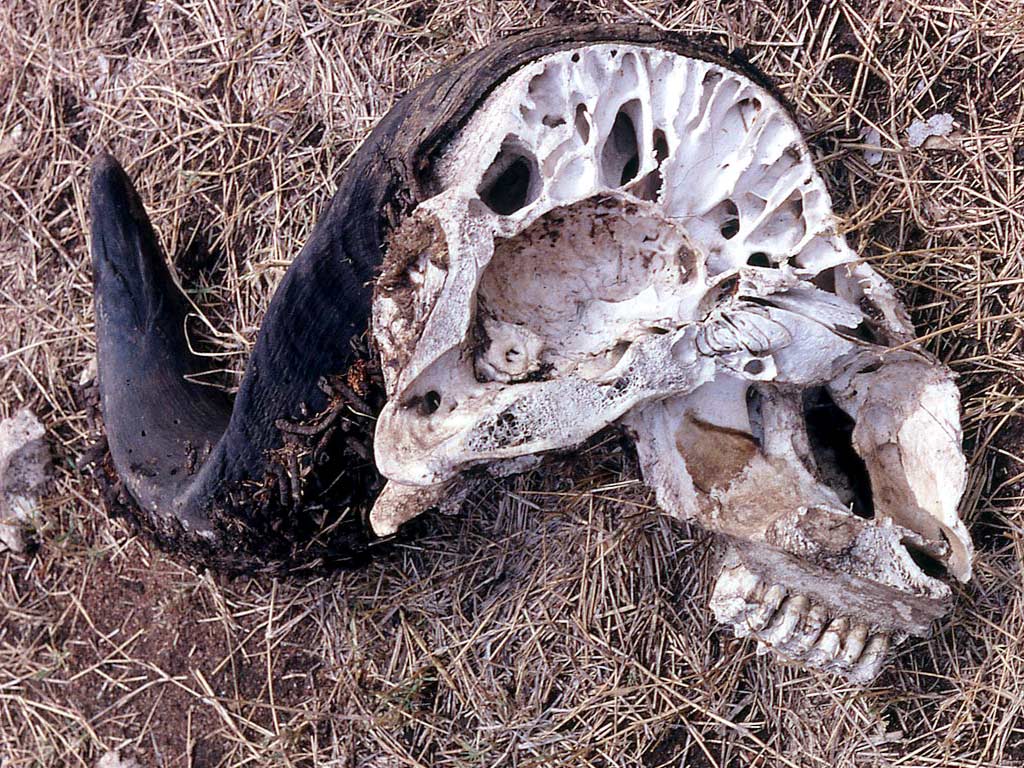
Лобные пазухи африканского буйвола (Syncerus caffer).
По указанным причинам лобные пазухи почти никогда не встречаются у древних форм, например, эоценовых и олигоценовых лошадей (Equidae), имеющих маленькие полушария. Как правило, эти костные полости развиваются у лошадей лишь с миоцена. Механическое значение пазух в крыше черепа очевидно у животных, несущих рога, например, у полорогих (Bovidae) и носорогов (Rhinocerotidae), где многочисленные перегородки в полостях ослабляют сотрясение при ударах рогами. В этих случаях костные пазухи имеются во всех костях черепной крыши, как у крупных носорогов, или заходят в костные пеньки рогов, а часто — и в их роговые отростки, как у полорогих. Однако в некоторых случаях в этом отношении имеются исключения: так, практически не пневматизирован череп бронтотериев (Brontotheriidae), а у некоторых оленей (Cervidae), несмотря на наличие крупных рогов, в крыше черепа кости лишены синусов. Максимального развития достигают костные полости в черепе слонов (Elephantidae), у которых вследствие этого достигается без заметного утяжеления значительное увеличение затылочной поверхности, на которой укрепляются мощные шейные связки и мышцы, поддерживающие тяжелую голову с бивнями.
Соотношение между мозговым и лицевым отделами черепа различно и зависит от разных причин. Лицевая часть заметно укорачивается в тех случаях, когда требуется сжатие челюстей значительной силы на их переднем конце (укорочение плеча сопротивления рычага), что имеет место, например, у хищников (схватывание и удержание добычи), грызунов (грызение), аминодонтов (Amynodontidae) (вытягивание из земли корневищ). Укороченный лицевой отдел высших приматов, так сказать, втянут под разросшуюся мозговую коробку. Напротив, лицевая часть удлиняется, когда, в связи с необходимостью перетирания жесткой растительной пищи, увеличивается общая жевательная поверхность щечных зубов — коренные удлиняются, а предкоренные в той или иной степени моляризуются, как, например, у жвачных и поздних лошадей. Удлинение лицевого отдела имеет место также в тех случаях, когда животное вскапывает мордой землю (свиньи), или когда требуется захватить одновременно большое количество пищи (китообразные), или при некоторых своеобразных способах питания (трубкозубы, муравьеды и др.). У мелких представителей любой группы головной мозг относительно велик, и в результате дорсальная поверхность черепа выглядит вздутой, а морда — сравнительно небольшой. Та же закономерность проявляется и в ходе онтогенеза млекопитающих. У молодых особей мозговая коробка в сравнении с лицевой частью обычно развита сильнее, однако лицевая часть растет быстрее, и у взрослых животных относительно длиннее.

Различия в пропорциях черепа (мозг заштрихован) у льва (Panthera leo) (вверху) и кошки (Felis silvestris) (внизу); вертикальная линия означает приблизительную границу между лицевым и мозговым отделами черепа.
Носовые кости (nasalia), прикрывающие носочелюстную область сверху, в большинстве случаев довольно большие и длинные, свободно выступающие над носовым отверстием. Иногда они доходят впереди до уровня передних краев предчелюстных костей, но чаще не достигают вершинами уровня последних, так что носовое отверстие на черепе направлено не прямо вперед, а вперед и несколько вверх. Значительное укорочение носовых костей и отодвигание носового отверстия назад, на верхнюю поверхность черепа, обычно связано с развитием гибкого рыла или хобота, когда костная в основании носовая трубка спереди переходит в подвижную хрящевую. Это имеет место, например, у тапиров (Tapiridae), астрапотериев (Astrapotheria), пиротериев (Pyrotheria), но особенно — у хоботных (Proboscidea). У млекопитающих, обладающих хоботообразным или подвижным носом (у кротов (Talpidae), свиней (Suidae) и др.), в результате окостенения носовой перегородки дополнительно развивается особая косточка — praenasale. Другой случай редукции носовых костей и отодвигания носового отверстия назад, на верхнюю поверхность черепа, имеет место у водных животных, что облегчает вдыхание воздуха при подъеме над поверхностью воды. Максимально это выражено у китообразных (Cetacea). Их «дыхало» находится на вершине черепа; носовая область сместилась назад, а затылочные кости вперед, так что прежняя крыша с поверхности черепа практически исчезла. Кроме того, у некоторых китов череп еще больше усложнен в связи с асимметрией.
У переднего края мозговой коробки, выше сошника и впереди от клиновидных костей развивается непарная решетчатая кость (ethmoidale). Ее нижняя сторона скрыта под твердым небом, а средняя часть образует носовую перегородку (lamina perpendicularis). Боковые отделы решетчатой кости обращены вовнутрь поперечными костными перегородками — продырявленными пластинками (lamina cribrosa), которые пронизаны многочисленными отверстиями для прохождения пучков волокон обонятельного нерва. Среди современных млекопитающих только решетчатые кости утконоса (Ornithorhynchus anatinus) несут каждая по одиночному отверстию. Обонятельные мешки у представителей рассматриваемого класса распространены назад и отделены от увеличенной полости черепа упомянутыми продырявленными пластинками. Срединная обонятельная кость (когда она присутствует) формируется в этой области и простирается вперед в качестве срединной перегородки между двумя носовыми полостями. Две наружные ноздри млекопитающих сближены у средней линии черепа, образуя в кости единое отверстие (как и у черепах).
В глубине носовых полостей на костях их стенок находятся обонятельные раковины (turbinalia) — сложно закрученные парные ячеистые образования из тонких костных пластинок, покрытых сверху слизистой оболочкой. Наиболее глубоко помещаются решетчатые pаковины (ethmoturbinalia), принадлежащие упомянутым выше боковым частям решетчатой кости. Выше и ближе к переднему носовому отверстию располагаются принадлежащие носовым костям и отходящие от их нижних стенок носовые раковины (nasoturbinalia). Наконец, впереди и ниже раковин решетчатой кости расположены отходящие от верхнечелюстных костей верхнечелюстные раковины (maxilloturbinalia). Последние могут иметь разнообразную форму: они образуют то одиночные завитки, то двойные, несущие иногда древовидные разветвления. Величина, сложность строения и соотношение размеров разных раковин варьирует в зависимости от степени развития органа обоняния.

Схематическое изображение расположения носовых раковин на поперечном разрезе через носовую полость: I-V — endoturbinalia; 1-6 — ectoturbinalia.
Вторичное твердое небо образуется широкими, направленными внутрь пластинами-отростками верхнечелюстных костей и собственно небными костями (palatinum); в переднем отделе к ним также присоединяются небные отростки предчелюстных костей. Все эти элементы сходятся по средней линии, или шву неба. Вторичное твердое небо отделяет носовую полость от ротовой и придает рылу трубчатую конструкцию, а вместе с ней — высокую прочность. Ширина и форма неба тесно связаны с расположением щечных зубов, т. е., в конечном счете, с характером жевательной функции. Протяженность твердого неба очень различна. Чаще всего оно тянется в виде непрерывной пластинки от переднего конца черепа до заднего края зубного ряда. Иногда по заднему краю небо сокращается в длину, оканчиваясь значительно ближе впереди заднего края зубов; порой от него остается всего лишь узкий (спереди назад) мостик между альвеолярными частями челюстей (у зайцеобразных). В других случаях небо, напротив, сильно удлиняется назад; максимально — у некоторых гиенодонтов и китообразных. Сзади вторичное костное небо продолжает слизистая складка — мягкое небо.
В передней части твердого неба, а именно на протяжении небных отростков предчелюстных костей, у многих млекопитающих заметны 2 удлиненных резцовых отверстия (foramina incisiva). При их помощи посредством парных небно-носовых каналов (ductus nasopalatinus) с ротовой полостью сообщается вомероназальный, или якобсонов, орган. В отличие от многих млекопитающих, у человека резцовое отверстие непарное, но следы былой парности все же заметны по его форме. Вомероназальный орган у человеческого зародыша в норме редуцируется на 9-м месяце внутриутробного развития. В задней части твердого неба находятся небные отверстия (foramina palatina), пропускающие вперед нервы и мелкие кровеносные сосуды.
К задним выступам небных костей, образующим желоб, прирастают относительно небольшие крыловидные кости (pterygoideum). У большинства млекопитающих эти кости вполне самостоятельны; между их крыловидными выступами (hamuli pterygoidei) часто находится заметная впадина (fossa mesopterygoidea), особенно бросающаяся в глаза, если смотреть на череп снизу. Обычно крыловидные кости срастаются с расположенным над ними клиновидным отделом мозговой коробки, образуя с каждой стороны пластинку нисходящего крыловидного отростка (processus pterygoideus). Небные и крыловидные кости занимают вертикальное положение и ограничивают с боков задние носовые ходы, или вторичные хоаны (choanae). Такое строение неба предотвращает закупорку носовых ходов (т. е. перерыв дыхания), в то время как пищевой комок задерживается в ротовой полости для его пережевывания. У других современных наземных позвоночных (кроме крокодилов и черепах) хоаны открываются не кзади от небных костей, а между верхнечелюстными костями. Эктоптеригоиды (ectopterygoideum) у териевых млекопитающих исчезли.
По сравнению с представленной картиной, строение неба пеликозавров является весьма примитивным. У них сохраняется четыре покровных элемента — сужающиеся между хоанами сошники, небные кости, маленькие остатки наружных крыловидных костей и крыловидные кости, простирающиеся вдоль большей части длины неба. На каждой крыловидной кости имеется крупный боковой выступ с дополнительными зубами. Сохраняются две замещающие кости небного комплекса. Верхнекрыловидная кость по-прежнему образует подвижное базальное сочленение с мозговым черепом. Эта кость образует углубление для базиптеригоидного отростка мозговой коробки и несет вертикальный отросток, восходящий сбоку от последней к крыше черепа. Квадратная кость пеликозавров остается крупной и не только образует сочленение с нижней челюстью, но и простирается далеко вперед вдоль внутренней поверхности крыловидной кости.
Ряд преобразований неба у более продвинутых зверообразных уже приближает его строение к типичному для млекопитающих. В передней части главным изменением является постепенное развитие вторичного неба. Предчелюстные, верхнечелюстные и далее назад небные кости загибаются вентральным краем к средней линии и смыкаются, образуя длинную «полку», расположенную ниже первичной крыши ротовой полости. Так возникают ходы, по которым воздух перед тем как попасть в «рот» проходит далеко назад от исходного местонахождения внутренних ноздрей. Парные сошники сливаются в единый элемет (они сохраняют свое положение в первичной крыше ротовой полости и поэтому почти не видны, если смотреть снизу). Сошники участвуют в формировании дна под клинообонятельной областью и в дальнейшем образуют перегородку между двумя носовыми полостями. Сама трубчатая клинообонятельная кость, служащая для прохождения передних нервов, образует со временем переднеклиновидную кость. Квадратные кости постепенно уменьшаются и теряют жесткую связь с другими элементами.
Как и у многих низших тетрапод, подвижное сочленение неба и мозговой коробки в линии млекопитающих утрачивается, а крыловидная и верхнекрыловидная кости прирастают к клиновидному отделу черепа. Стержневидный отросток верхнекрыловидной кости врастает в стенку мозговой коробки в виде крылоклиновидной кости. Кроме того, ранее крупные крыловидные отростки сильно уменьшаются. Часто в их основании расположены крыловидные каналы, через которые вперед проходят сосуды и нервы.
Боковой выступ на небе исчезает (и вместе с ним маленькая наружная крыловидная кость), а задняя ветвь крыловидной кости, когда-то вытянутая назад и наружу к квадратной кости, постепенно сокращается. Таким образом, исходные замещающие элементы неба у млекопитающих заметно изменяются и больше не участвуют в формировании небных структур. Судя по всему, появление вторичного неба было в значительной мере связано с обретением постоянной температуры тела и необходимостью более тщательно обрабатывать во рту относительно большее количество пищи.
Первичная верхняя челюсть, относящаяся к висцеральным компонентам черепа, состоит из четырех окостенений: покровных небных и крыловидных костей, замещающих крылоклиновидных костей и наковален. У млекопитающих от прямого участия в схватывании и механической обработке добычи все они отстранены. Данные функции выполняет вторичная верхняя челюсть, сформированная двумя парными покровными костями, несущими в своих ячейках зубы. Большая их часть помещается в массивных и высоких верхнечелюстных костях (maxillaria), а в расположенных кпереди от них предчелюстных, или межчелюстных, костях (ргаеmaxillaria, или intermaxillaria) находятся только резцы. Верхнечелюстные кости также образуют боковые части переднего отдела черепа и очерчивают наружные стенки носовой полости. В определенной мере в этом задействованы и предчелюстные кости, которые поднимаются и примыкают к латеральным краям носовых костей.
На передней поверхности каждой верхнечелюстной кости около ее подглазничного края расположено подглазничное отверстие (foramen infraorbitale). Инoгда оно вытягивается в подглазничный канал (canalis infraorbitalis), служащий для прохождения кровеносных сосудов и нервов из глазницы вперед к поверхности рыла. У грызунов сильно развиты жевательные мышцы, и сбоку лицевой части черепа может находиться глубокая выемка, в которой они начинаются, а подглазничный канал иногда превращается в крупное окно для их размещения.
Степень развития предчелюстных костей сильно различается и тесно связана с сидящими в них верхними резцами. При редукции резцов эти кости уменьшаются, иногда до полного исчезновения (многие рукокрылые); при их большом развитии они увеличиваются, особенно сильно — их носовые отростки, направляющиеся назад-наверх и окаймляющий снаружи переднее носовое отверстие. Эти отростки передают давление, испытываемое резцами при их работе (например, у грызунов при грызении) в лобную кость, к которой он в таких случаях примыкает, и дальше, в задние отделы черепа. Исключительно больших размеров достигает носовая часть предчелюстных костей у хоботных, у которых она заключает в себе огромные альвеолы гигантских бивней. Очень сильно разрастается носовая ветвь предчелюстных костей у китообразных, где это связано с отодвиганием назад-вверх носового отверстия. У приматов предчелюстные кости срастаются с верхнечелюстными костями. У однопроходных предчелюстные кости несут отросток, подобный таковому рептилий (processus ascendus).

Череп 3-месячного щенка песца (Аlорех lagopus) (А) и сагиттальный распил черепа взрослой особи (Б): 1 — носовая кость; 2 — верхнечелюстная кость; 3 — слезная кость; 4 — скуловая кость; 5 — лобная кость; 6 — крылоклиновидная кость; 7 — теменная кость; 8 — чешуйчатая кость; 9 — межтеменная кость; 10 — верхнезатылочная кость; 11 — боковая затылочная кость; 12 — затылочный мыщелок; 13 — околоушная кость (сосцевидный отдел); 14 — барабанная кость; 15 — место вторичного челюстного сустава; 16 — крыловидная кость; 17 — глазоклиновидная кость; 18 — небная кость; 19 — переднеклиновидная кость; 20 — предчелюстная кость; 21 — верхнечелюстная раковина; 22 — носовая раковина; 2З — обонятельные раковины; 24 — продырявленная пластинка срединной обонятельной (решетчатой) кости — передняя стенка мозговой капсулы; 25 — околоушная кость (каменистый отдел); 26 — основная клиновидная кость; 27 — носоглоточный ход (вторичная хоана); 28 — сошник; 29 — вторичное твердое небо.
Нижняя челюсть (mandibula) млекопитающих образована лишь парной зубной костью (dentale), формирующейся как покровная на основе первичного меккелева хряща. Часть челюсти, несущая зубы, именуется ее телом (corpus mandibulae). За последними коренными зубами располагается нижнечелюстная ветвь (ramus mandibulae), которая поворачивается под тупым углом в верхнезаднем направлении. Место перехода нижнего края тела в задний край ветви нижней челюсти называется ее углом (angulus mandibulae). Зачастую здесь имеется угловой отросток (processus angularis), нередко крупный, или шероховатое утолщение, служащие местами крепления челюстной мускулатуры. Ветвь разделяется на восходящие венечный и сочленовный отростки. К венечному отростку (processus coronoideus) прикрепляется височная мышца, которая закрывает челюсть. Вытянутый в поперечном направлении сочленовный отросток (processus condylaris) снабжен округлой в сечении суставной головкой для подвижного сочленения с гленоидной суставной ямкой в нижнем отделе чешуйчатой кости (fossa condyloidea). У многих млекопитающих нижняя челюсть широко подвижна в горизонтальной плоскости.
На внутренней стороне челюсти близ ее заднего конца имеется нижнечелюстное отверстие, пропускающее нервы и кровеносные сосуды к нижним зубам, а в передней части наружной стороны — подбородочное отверстие, через которое кровеносные сосуды и нервы проходят к подбородку и к области нижней губы.
Форма нижней челюсти почти полностью определяется способом питания. У хищников, которым для овладения живой добычей требуется схватывание и удержание при помощи клыков, а размельчается пища (наподобие работы ножниц) верхними и нижними щечными зубами, для чего необходимо сильное сжатие челюстей. В связи с этим их нижняя челюсть обычно короткая, и диастема (разрыв в зубном ряду) отсутствует или укорочена (укорочение плеча сопротивления рычага). Сама челюсть в горизонтальной части высокая (противодействие излому при сжатии); венечный отросток длинный по отношению к уровню суставной головки, отвесный (удлинение плеча силы височного мускула, сжимающего челюсти, который прикрепляется на вершине отростка) и широкий спереди назад (противодействие его излому при тяге височного мускула и увеличение поверхности eгo прикрепления).
Обе половины челюсти действуют как одно целое, и их передние отделы неподвижно сращены между собой по поверхности сращения или симфизу. При этом у хищников суставная головка челюсти обычно расположена очень низко — часто на уровне коренных зубов, чем достигается, во-первых, возможность значительного возвышения венечного отростка над уровнем сустава без чрезмерного eгo удлинения, во-вторых, возможность шире открывать пасть при схватывании добычи и, в-третьих, при таком положении оси вращения подъем челюсти на конечных стадиях смыкания происходит почти вертикально, что способствует разрезанию пищи. Ниже всего расположен сустав челюсти у саблезубых кошек (Machairodontinae), где это имеет особое значение. Сильно выражены места прикрепления мышц, сжимающих челюсти: широкий угловой отдел (pars angularis) с глубокой массетерной ямкой на наружной поверхности, с угловым отростком (processus angularis) и гребнями и бороздами для крыловидных мышц на внутренней поверхности. Все описанные приспособления сильнее развиты у хищников, питающихся исключительно мясной пищей (кошки (Felidae), гиены (Hyaenidae)), слабее — у хищников со смешанным пищевым режимом (барсуки (Melinae), медведи (Ursidae), отчасти собаки (Canidae)).
Много общего с нижними челюстями хищников имеют нижние челюсти древних приматов, которым приходится раздавливать зубами твердые плоды. Морфологическое отличие состоит в более высоком положении суставной головки, что и понятно: обезьянам не требуется так широко открывать пасть, как хищникам, а подъем оси вращения вызывает на конечных стадиях смыкания челюстей некоторое движение нижних зубов вперед по поверхности верхних, что способствует перетиранию пищи.
Совершенно иное строение имеет челюсть растительноядных, у которых размельчение пищи происходит не путем разрезания или раздавливания, а посредством перетирания, и захват пищи не требует сильного сжатия челюстей. Характернее всего этот тип челюсти выражен у жвачных. Челюсть здесь длинная и низкая, с длинной диастемой, венечный отросток невысоко поднят над уровнем суставной головки и загнут назад, суставная головка лежит высоко над уровнем зубного ряда, так как широкое открывание пасти не требуется, а смыкание челюстей при таком положении оси вращения сопровождается перетирающими заднепередними движениями нижних зубов по верхним. Массетерная ямка и угловой отросток не развиты. Так как при перетирающем размельчении пищи движение нижней челюсти происходит не только в одном направлении (у жвачных — внутрь и наружу), но имеет комбинированный характер — частично также вперед и назад, то каждая из половин челюсти действует до известной степени независимо от другой; поэтому они в переднем отделе соединены при помощи хряща и сохраняют некоторую взаимную подвижность.
Между двумя описанными типами строения челюстей наблюдаются многочисленные промежуточные — у животных с нерезко выpaженной пищевой специализацией: со смешанным пищевым режимом, с питанием более мягкими частями растений и пр. Описанные приспособления выражены у них в различной степени и в разных комбинациях. Таковы, например, насекомоядные, часть креодонтов, кондиляртры, непарнопалые, даманы и многие другие.

Череп зайца-беляка (Lepus timidus); вид снизу (А), сверху (В) и сбоку (С): 1 — предчелюстная кость; 2 — верхнечелюстная кость; 3 — переднее небное отверстие; 4 — небная кость; 5 — сошник; 6 — скуловая кость; 7 — надглазничный отросток лобной кости; 8 — крылья крыловидной кости; 9, 10 — передняя клиновидная кость; 11 — основная клиновидная кость; 12 — основная затылочная кость; 13 — переднее раваное отверстие, через которое из полости черепа в глазницу выходят нервы III-й, IV-й и VI-й пары и две первые ветви V-й пары; 14 — слуховой пузырь; 15 — затылочный мыщелок; 16 — большое затылочное отверстие; 17 — носовые кости; 18 — скуловой отросток челюстной кости; 19 — лобная кость; 20 — скуловой отросток височной кости; 21 — теменная кость; 22 — слуховое отверстие; 23 — верхняя затылочная кость; 24 — глазнично-основная кость; 25 — слезная кость; 26 — сочленовный отросток нижней челюсти; 27 — венечный отросток; 28 — угловой отросток.
Своеобразно строение нижней челюсти грызунов, у которых сочетается необходимость сильного сжимания челюстей в передней части (грызение) с перетирающими переднезадними жевательными движениями. Короткая и относительно высокая челюсть (однако с длинной диастемой для помещения огромных резцов) coчетается у них с высоко расположенной суставной головкой и с коротким венечным отростком. Суставная головка обычно вытянута в продольном направлении и, соответственно, суставная ямка имеет корытообразную форму. Грызение происходит при переднем положении нижней челюсти с нефиксированной осью челюстного рычага и при ее заднем положении, когда ось челюстного рычага фиксируется, упираясь суставной головкой в сочленовную поверхность. Разделение функций грызения и перетирания пищи приводит к образованию дополнительных сочленений нижней челюсти с черепом, способствующих фиксации оси челюстного рычага. Так, гоферы (Geomyidae) имеют три сочленения нижней челюсти с черепом. При сильной работе резцов ряды коренных могут совсем не соприкасаться, что предохраняет их от излишнего снашивания. И у других форм сильное развитие резцов, имеющее тот или иной биологический смысл, вносит значительные видоизменения в строение челюсти (у хоботных, некоторых носорогов, астрапотериев и др.).
В тех случаях, когда жевательная функция челюстей ослабевает и ротовая полость служит преимущественно для наполнения ее мягкой, легко усвояемой пищей, челюсти делаются слабыми, тонкими и низкими, с низким венечным отростком; обычно это связано с упрощением зубов (у ластоногих, зубатых китов, трубкозубов и др.). При полной утрате зубов и жевательной функции челюстей нижняя челюсть делается очень тонкой, палочковидной; обе ее половины на переднем конце очень слабо связаны друг с другом (у однопроходных, муравьедов, беззубых китов).
Разнообразное строение нижней челюсти млекопитающих, всегда связанное с xapaктeром зубной системы и в значительной степени отражающее биологию животных, делает эту часть скелета очень важной для систематики и понимания образа жизни ископаемых представителей класса.
Строго соответствует характеру жевательных движений и строению зубов также форма челюстного сустава. У хищников и насекомоядных, где движения челюсти исключительно вертикальные (связаны с режущими зубами), т. е. представляют собой вращение вокруг оси, поперечной к оси черепа, суставная головка ее имеет форму цилиндрического поперечного валика, вращающегося во впадине соответствующей формы. Высокие предсуставный и засуставный отростки так охватывают спереди и сзади суставную впадину, что исключаются скользящие движения головки (и челюсти) взад и вперед. У свинообразных, примитивных хоботных, отчасти приматов, т. е. у животных с бугорчатыми зубами, форма челюстного сустава напоминает форму его у хищных, однако некоторые боковые и переднезадние движения у них возможны.
В тех случаях, когда размельчение пищи происходит путем перетирания передне-задними движениями (связано с поперечногребенчатыми и поперечнопластинчатыми зубами), как у слонов и зайцеобразных, суставная головка нижней челюсти яйцевидная с большой осью, расположенной продольно; она движется в аналогично расположенной желобовидной суставной впадине. Наконец, при более сложных перетирающих движениях, в которых участвуют и боковой и передне-задний компоненты (связано с лунчатыми и гребенчатыми зубами, имеющими и продольные и поперечные гребни), как у жвачных, большинства непарнопалых и многих грызунов, сустав имеет слабый рельеф, допускающий разнообразные движения.

Левые половины нижних челюстей зверообразных (сверху вниз — пеликозавр диметродон (Dimetrodon), горгонопс цинароид (Cynaroides), цинодонт циногнат (Cynognathus)) и млекопитающего (плацентарное волк (Canis)), иллюстрирующие редукцию элементов челюсти: 1 — пластинчатая кость (spleniale); 2 — зубная кость (dentale); 3 — угловая кость (angulare); 4 — надугловая кость (supraangulare); 5 — articulare; 6 — венечная кость (coronare); 7 — prearticulare; 8 — венечный отросток; 9 — мыщелок; 10 — нижнечелюстное отверстие; 11 — угловой отросток; 12 — подбородочное отверстие.
Большинство костей, исходно принадлежавших нижней челюсти, утратились или резко изменились. У зверообразных значение зубной кости постепенно растет. Она вытягивается назад, приближаясь к первичному челюстному суставу, а несколько спереди, под височной областью, на ней образуется направленный вверх отросток для прикрепления височной мышцы. Попутно уменьшаются размеры других окостенений нижней челюсти, которые у млекопитающих вообще исчезают из нее, и их остатки следует искать в слуховом аппарате. Сегодня известно много продвинутых зверообразных, у которых одновременно развиты и функционируют и старый челюстной состав пеликозавров (сочленовная - квадратная кости), и новый сустав млекопитающих (зубная - чешуйчатая кости). Эта система будет работать, если оба сустава находятся на одной поперечной оси (дверь можно навесить на сколько угодно петель, если все они расположены на одной прямой). Формирование нового челюстного сустава — только одно из направлений глубокого преобразования всего жевательного аппарата в ходе эволюции млекопитающих; его мышцы также совершенно непохожи на присутствующие у более примитивных форм. Данные изменения в первую очередь обусловливались потребностью теплокровных организмов в большем количестве пищи и ее пережевывании.

Челюстной сустав продвинутого цинодонта пробайногната (Probainognathus), вид снизу (А) и изнутри (Б). У этой формы функционирует и старый сустав между сочленовной и квадратной костями и новый сустав между зубной и чешуйчатой костями. Следует обратить внимание на то, что сочленовная кость (которая вскоре станет молоточком), квадратная (наковальня) и стремечко образуют цепочку, ведущую от области барабанной перепонки (не показана, расположена сзади и снизу от челюстного сустава) к слуховой капсуле мозговой коробки. 1 — articulare; 2 — зубная кость (dentale); 3 — суставная поверхность на чешуйчатой кости, с которой сочленяется зубная кость; 4 — чешуйчатая кость (squamosum); 5 — квадратная кость (quadratum); 6 — стремечко (stapes); 7 — затылочный мыщелок; 8 — скуловая кость (jugale).
Иные исходные предпосылки к преобразованиям нижней челюсти связаны с адаптацией к восприятию звуков в воздушной среде, которая уже у ранних зверообразных пошла иным путем, чем у других древних тетрапод. Проксимальная часть нижнечелюстного меккелева хряща, филогенетически соответствующая замещающей сочленовной кости (goniale), дала начало III-й звукопередающей косточке — молоточку (malleus), непосредственно связанному с барабанной перепонкой. Задняя часть небно-квадратного хряща, соответствующая квадратной кости (quadratum) прочих позвоночных, сильно редуцировалась, перешла в ушную область и преобразовалась во II-ю звукопередающую косточку — наковальню (incus). Сочленение наковальни с молоточком соответствует первичному челюстному суставу рептилий. Верхний элемент подъязычной дуги, составляющий у рыб подвесок (hyomandibulare), у амфибий, рептилий и птиц вошел в слуховой аппарат под видом слуховой косточки столбика (columella), а у млекопитающих превратился в I-ю по счету звукопередающую косточку — стремечко (stapes), запирающее овальное окно. Наличие 3 связанных между собой слуховых элементов — морфологическая черта, присущая исключительно млекопитающим. С нижней челюстью они теряли связь дважды — независимо у предков однопроходных и у Theriiformes.

Нижняя челюсть зародыша кролика (Oryctolagus cuniculus): 1 — меккелев хрящ; 2 — молоточек (malleus); 3 — наковальня (incus); 4 — стремечко (stapes); 5 — зубная кость (dentale); 6 — goniale; 7 — барабанная кость (tympanicum).
Подъязычный аппарат млекопитающих состоит из единственной подъязычной кости (hyoideum). Она имеет массивную среднюю часть — тело (corpus), и две пары расположенных по его сторонам отростков — передних, или малых (cornu minus), и задних, или больших (cornu majus) рожков. Тело подъязычной кости поперечно вытянуто и обычно дуговидно изогнуто; оно расположено в дне глотки у основания языка вентральнее переднего конца трахеи. Рожки тянутся вверх и наружу с обеих сторон трахеи. В тонком отростке переднего рожка, дотягивающегося наверху до слухового отдела, могут встречаться дополнительные окостенения. У некоторых млекопитающих, в том числе и у человека, этот дорсальный конец рожка может прирастать к каменистой части височной кости в виде небольшого шиловидного отростка (processus styloideus). Согласно эмбриологическим данным, тело подъязычной кости образовалось из остатков копул, передние рожки — из гиоида, задние — из I-й жаберной дуги. II-я и III-я жаберные дуги соединились и образовали щитовидный хрящ гортани, а остатки IV-й и V-й дуг образуют скелет остальных гортанных хрящей.

Дериваты жаберных дуг у человека. 1, 2, 3, 4, 7 — I-я-V-я примордиальные дуги. Из I-й дуги, меккелева хряща, возникает слуховая косточка — молоточек (malleus, или articulare) (16). Из хряща, соответствующего квадратной кости, образуется наковальня (17). Обе эти косточки изображены в естественном положении после удаления барабанной перепонки; 15 — наружное ухо; 14 — стремечко, образующееся из верхней проксимальной части подъязычной дуги; из II-й подъязычной дуги возникают: в проксимальной части — processus styloideus (13), из дистальной — малые рожки подъязычной дуги и подъязычный хрящ; из III-й дуги (первой жаберной) возникает большая часть подъязычного хряща (обозначен черным) и большой рог подъязычной дуги. Cartilago triticea (11) и большие рожки щитовидного хряща составляют остатки первичного соединения гиоидного и тиреоидного аппаратов; из IV-й дуги (второй жаберной) возникает верхний отдел щитовидного хряща (5); из V-й дуги (III-й жаберной) — нижний отдел этого хряща (6). Вероятно, что из V-й дуги образуются и черпаловидные хрящи (10); 8 — трахея; 9 — перстневидный хрящ; 12 — ligamentum stylohyoideum, идущее от processus styloideus к малым дугам подъязычного хряща.
вперед
в оглавление