в оглавление
назад
Дыхательная система
Дыхательная система служит для газообмена организма с окружающей средой, обеспечивая поступление кислорода и выведение углекислого газа. У млекопитающих она представлена легкими и воздухопроводящими (дыхательными) путями: верхними и нижними. В систему верхних дыхательных путей входят носовая полость, носоглоточные ходы, а частично — и ротовая полость, поскольку она тоже может участвовать в процессе дыхания. Система нижних дыхательных путей состоит из гортани (иногда ее относят к верхним дыхательным путям), трахеи и бронхов.
Если у низших позвоночных носовая полость обычно занимает короткий участок на переднем конце головы, то у типичных млекопитающих (исключение составляют сухоносые обезьяны (Haplorhini)) она достигает предельных размеров, внедряясь далеко назад в область глазниц и иногда занимая более половины длины черепа. Ее размер увеличен благодаря тому, что у млекопитающих имеется вторичное небо. В результате первичный свод ротовой полости оказался заключенным в носовую полость; бывшая дорсальная часть ротовой полости спереди образовала вентральную часть основной носовой полости, а позади — длинный носоглоточный ход, несущий воздух назад к вторичной хоане. Эти новые дополнения к носовому аппарату служат преимущественно для дыхания, однако обоняние в большинстве случаев тоже сильно развито. Объемистая носовая полость млекопитающих никоим образом не является простым мешком. От ее боковых стенок ответвляются раковины чрезвычайно сложной структуры, разделяющие воздушный путь на ряд частично изолированных ходов. Костная основа раковин образует в поперечном сечении подобие сот. В типичном случае решетчатая кость образует от двух до четырех раковин, а носовая и верхнечелюстная — еще по одной.
Функционально носовая полость разделяется на преддверие, нижний дыхательный и задневерхний обонятельный отделы. Преддверие начинается от наружных ноздрей, его задача состоит в улавливании относительно крупных частиц (пыли), проникающих с воздухом. Этот отдел более развит у обитателей степных и пустынных районов, где запыленность воздуха особенно велика. Мелкие пылевые частицы улавливаются в дыхательном отделе, выстланном слизистой оболочкой из мерцательного эпителия, под которой залегает густая венозная сеть. Здесь происходит не только дальнейшая очистка воздуха от механических примесей, но и его обеззараживание с помощью слизи, выделяемой специальными железами и содержащей активные бактерицидные вещества (например, лизоцим). Согласованные движения ресничек мерцательных клеток выводят слизь с осевшими частицами наружу. Вместе с тем вдыхаемый воздух, проходя между складками носовых раковин, согревается и увлажняется протекающей по капиллярам кровью, а также отдает ей небольшую часть кислорода. На обратном пути воздух отдает тепло и воду остывшим при вдохе перегородкам. При перегреве описанный теплообменник используется только на вдохе, при выдохе же нагретый воздух выпускается через рот. Обонятельный отдел, или обонятельная капсула, представляет собой расширение с лабиринтом обонятельных раковин, выстланных чувствительным эпителием.
У большинства млекопитающих от носовой полости ответвляются воздушные пазухи (sinus pneumatici), служащие для облегчения черепа. Обычно они проникают в верхнечелюстную, лобную, обонятельную и клиновидную кости; достигают затылка в высоком куполообразном черепе слона и у некоторых других млекопитающих.
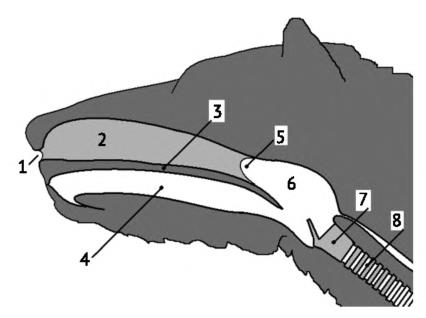
Верхние дыхательные пути млекопитающего: 1 — внешние ноздри; 2 — полость носа; 3 — вторичное небо; 4 — ротовая полость; 5 — хоаны; 6 — носоглотка; 7 — гортань; 8 — трахея.
Из полости носа воздух поступает в носоглотку, которая отделена от ротовой полости мягким небом и открывается в глотку (pharynx) отверстием хоан. У многих млекопитающих мягкое небо длиннее, чем у человека, вследствие чего задняя стенка глотки через пасть не видна. На дне глотки расположено срединное отверстие — гортанная щель (glottis). Сразу за ней дыхательный проход расширяется в камеру — гортань (larynx), лежащую под глоткой и несколько позади нее. По сравнению с другими позвоночными, строение гортани усложнено. Ее стенки поддерживаются гиалиновыми хрящами, к которым крепятся поперечнополосатые гортанные мышцы. Спереди и с боков находится крупный щитовидный хрящ (cartilago thyroidea), формирующий корпус гортани. Под ним расположен перстневидный хрящ (cartilago cricoidea), образующий кольцо на переходе в трахею. По бокам дорсальной поверхности гортани над перстеневидным хрящом имеются два черпаловидных хряща (сartilagines arytenoidea). От переднего края щитовидного хряща вверх и вперед выступает поперечно ориентированный лепесткообразный клапан — надгортанник (epiglottis) с пластинкой волокнистого хряща внутри. У многих млекопитающих он прилегает в положении покоя сзади к краю мягкого неба и устанавливает прямое соединение носоглотки с гортанью. При проглатывании корма надгортанник поворачиваться назад, закрывая поперечно вытянутый вход в гортань. Наличие щитовидного хряща и надгортанника — отличительный признак млекопитающих, у прочих позвоночных они отсутствуют. Поддерживающие гортань хрящи соединяются связками, мембранами, относительно подвижными суставами и снабжены мышцами.
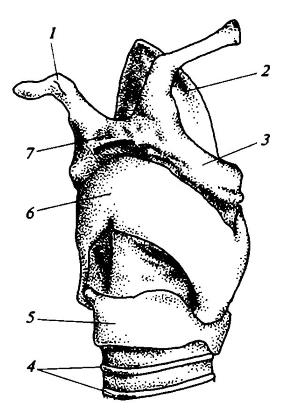
Гортань кошки (Felis silvestrus), вид вполоборота снизу и слева: 1 — малый рожок подъязычной кости; 2 — надгортанник; 3 — большой рожок подъязычной кости; 4 — хрящевые кольца трахеи; 5 — перстневидный хрящ; 6 — щитовидный хрящ; 7 — тело подъязычной кости.
Между щитовидным и черпаловидными хрящами на внутренней стороне боковых стенок гортани натянуты голосовые связки (ligament vocalia) — парные вертикальные складки слизистой оболочки, содержащие эластичную ткань и приводимые в движение особыми мышцами. Эти связки могут плотно прижиматься друг к другу и тогда начинают вибрировать при прохождении между ними потока выдыхаемого воздуха. Вибрация голосовых связок производит звуки (у ряда млекопитающих это происходит вследствие вибрации других частей гортани).
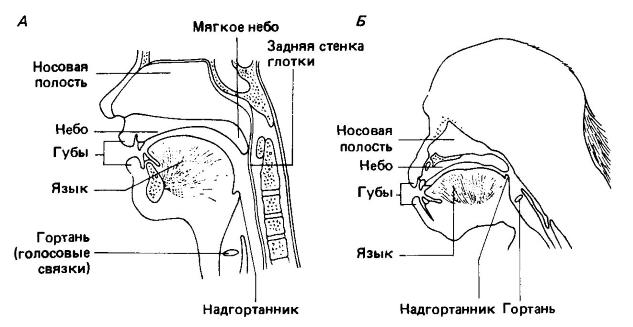
Схематическое изображение (сагиттальный разрез) головы и шеи взрослого человека (Homo sapiens) (А) и взрослого шимпанзе (Pan troglodytes) (Б). У шимпанзе и человеческого плода гортань расположена в верхней части голосового пути, тогда как у взрослых людей — в нижней ее части. Такое расположение гортани и дает возможность человеку изменять с помощью языка конфигурацию полости глотки и таким образом производить широкий спектр модулированных звуков.
Между перстневидным и щитовидным хрящами расположены особые мешковидные полости — желудочки гортани. У некоторых млекопитающих они сильно разрастаются и образуют гортанные мешки, играющие роль резонаторов. У обезьян-ревунов (Alouatta) такие мешки помещаются в пузыревидном вздутии подъязычной кости. Голоса большинства млекопитающих ограничены преимущественно низкочастотным диапазоном, что, однако, не уменьшает возможность передачи емкой и сложной информации. Часто сильно отличаются голоса не только различных видов, но и особей разного пола одного вида.

Горловой резонаторный мешок кричащего сиаманга (Symphalangus syndactylus). Сиаманги образуют долговременные пары, сообщая об этом громким дуэтом, в котором песни самца и самки несколько различаются. Самец поет, тогда как самка издает серии гудков и подобие лая. Детеныши также могут присоединиться к семейному хору. Некоторые исследователи считают, что пение дуэтом усиливает взаимные связи внутри пары размножающихся сиамангов. Однако дуэты чаще всего исполняются, когда пара чувствует присутствие другой семейной группы, которая может претендовать на ее участок. Заявляя таким образом о своих территориальных правах, сиаманги уменьшают риск столкновения и поддерживают друг друга.
В носоглотке зубатых китов (Odontoceti) имеются специальные образования — «звуковые губы», которые вибрируют, когда воздух проходит сквозь носовое отверстие. Большинство зубатых китов имеют две пары звуковых губ, что позволяет одновременно издавать два звука. Череп и мягкие ткани головы концентрируют звуковые колебания и играют роль звуковой линзы. У усатых китов (Mysticeti) звуковые губы отсутствуют, и образование звуков происходит в глотке, но поскольку у них нет голосовых связок, точный механизм остается неясным. Этот процесс не является полностью аналогичным таковому наземных млекопитающих, поскольку китам для создания звуков нет необходимости выдыхать воздух.
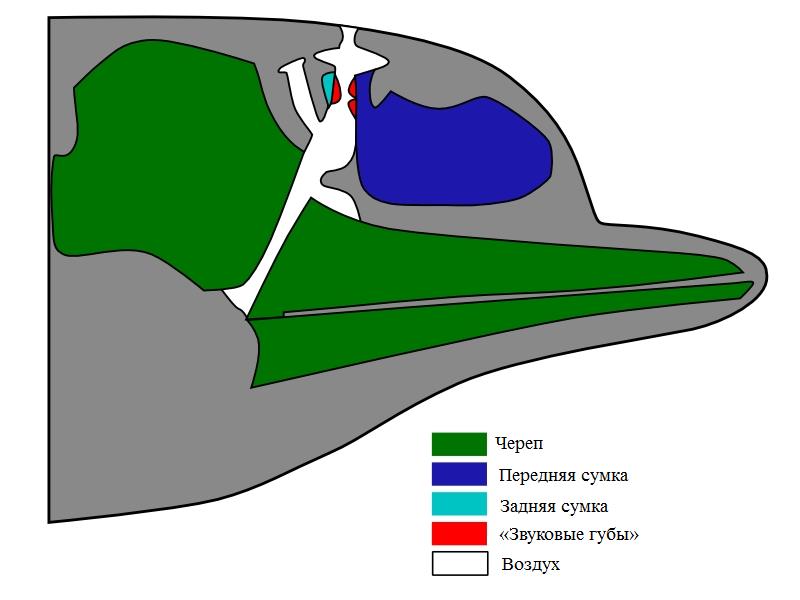
Органы зубатого кита, участвующие в формировании звуков.
Сзади гортань переходит в дыхательное горло, или трахею (trachea) — непарную трубку, прилегающую к пищеводу вентрально. Стенки трахеи поддерживаются многочисленными незамкнутыми сверху кольцами трахеальных хрящей (cartilagines tracheales), которые защищают ее при сокращении мышц шеи, а также при вдохе, когда давление в ней становится ниже атмосферного. Дорсальная стенка трахеи волокнистая, что позволяет пищеводу расширяться при глотании. Войдя в грудную полость, трахейная трубка разделяется на два главных бронха (bronchi principales). Они имеют меньший диаметр и идут каждый к основанию своего легкого.
Легкие (pulmones) представляют собой крупные, латерально расположенные губчатые образования, пронизанные многочисленными каналами. Они относительно коротки в переднезаднем направлении, но широки. Вместе с сердцем, лежащим между ними, легкие занимают большую часть грудной клетки. Они не имеют полостей, и избежать возникновения значительных внутренних напряжений в ходе изменения объема легких удается благодаря их разделению на доли и возможности взаимного скольжения этих долей. В результате того, что сердце несколько смещено влево, число долей в правом легком (до 6) всегда больше, чем в левом (до 3). Доли делятся на более мелкие сегменты, а те в свою очередь — на мельчайшие дольки. Соседние легочные элементы отделены друг от друга прослойками соединительной ткани. Помимо определенного количества последней, в стенках легких содержатся гладкие мышечные волокна и множество кровеносных сосудов. Наружная и внутренняя поверхности легких выстланы эпителием; над кровеносными капиллярами он может быть крайне тонким. Снаружи легкие покрыты плотной соединительнотканной оболочкой — легочной плеврой, так что каждое легкое оказывается внутри отдельной плевральной полости (cavum pleurale). В свою очередь, стенки грудной полости выстланы пристеночной плеврой. Пространство между легочной и пристеночной плеврой — плевральная щель — заполнено жидкостью, сходной по составу с лимфой и играющей роль смазки. Грудная полость отделяется от брюшной полости сухожильно-мышечной перегородкой — диафрагмой (diaphragma).
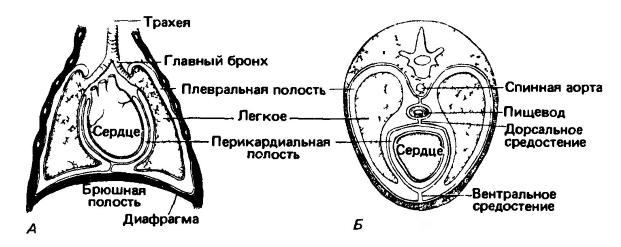
Схемы фронтального (А) и поперечного (Б) разрезов грудной клетки млекопитающего; показано расположение сердца, легких, плевральных и перикардиальной полостей.
При переходе в легкие главные бронхи сначала продолжаются в их толще, а затем делятся на все более мелкие бронхи — сегментарные (II-го порядка), субсегментарные (III-V-го порядков) и дольковые (VI-го порядка). Стенки бронхов образованы гиалиновыми хрящевыми кольцами, препятствующими их спадению, и гладкими мышцами; изнутри бронхи выстланы слизистой оболочкой. В свою очередь, дольковые бронхи разветвляются на бронхиолы (bronchioli), диаметр которых у человека не превышает 1 мм. Наиболее крупные, терминальные бронхиолы обладают лишенными хряща стенками и содержат гладкомышечные клетки, что позволяет регулировать воздушный поток и контролировать сопротивление ему. Терминальные бронхиолы делятся на респираторные (дыхательные) бронхиолы I-го порядка, которые дают респираторные ответвления II-го и III-го порядков. Стенки дыхательных бронхиол сформированы реснитчатыми эпителиальными клетками и альвеолоцитами, не содержат гладкомышечных клеток. Бронхиолы участвуют в проведении воздуха и обеспечении санации дыхательных путей.
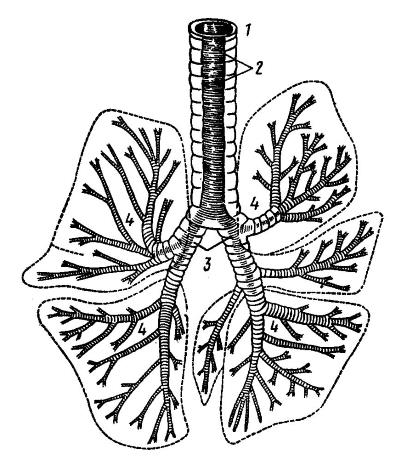
Бронхиальное древо бурого медведя (Ursus arctos): 1 — трахея; 2 — ее мембранная часть; 3 — главные бронхи; 4 — ветвление бронхов.
Мельчайшие респираторные бронхиолы образуют несколько генераций альвеолярных ходов (ductus alveolaris), несущих ячеистые расширения — альвеолярные мешочки и альвеолы (alveolae pulmonis). Последние представляют собой микроскопические пузырьки, собранные гроздьями на стенках бронхиол или альвеолярных мешочков наподобие виноградин в кисти. Диаметр альвеол составляет от 10 до 35 мкм, это — предельно малая величина, при которой поверхностное натяжение на влажных стенках альвеол не мешает расправлению легких. Чтобы диаметр альвеол не уменьшался ниже допустимого, на них имеется особая липидная пленка, снижающая поверхностное натяжение — сурфактант. Тонкие стенки альвеол выстланы дыхательным эпителием и оплетены густой сетью капилляров. Эти капилляры ответвляются от легочной артерии, идущей от сердца; в сердце кровь возвращается по легочной вене. Между кровью и полостью альвеол находятся лишь тонкие мембраны, сквозь которые легко происходит диффузия газов и обмен с воздухом кислородом и двуокисью углерода (наряду с альвеолами, в газообмене и синтезе сурфаканта участвуют респираторные бронхиолы). Высокая эффективность легких при таком устройстве определяется развитым внутренним бронхиальным древом, которое обеспечивает равномерное участие альвеол во взаимодействии с воздухом. Но сам принцип возвратно-поступательного движения воздуха в системе не допускает столь интенсивной степени его обновления в альвеолах, как в легких птиц, где он проходит по петлевидной траектории.
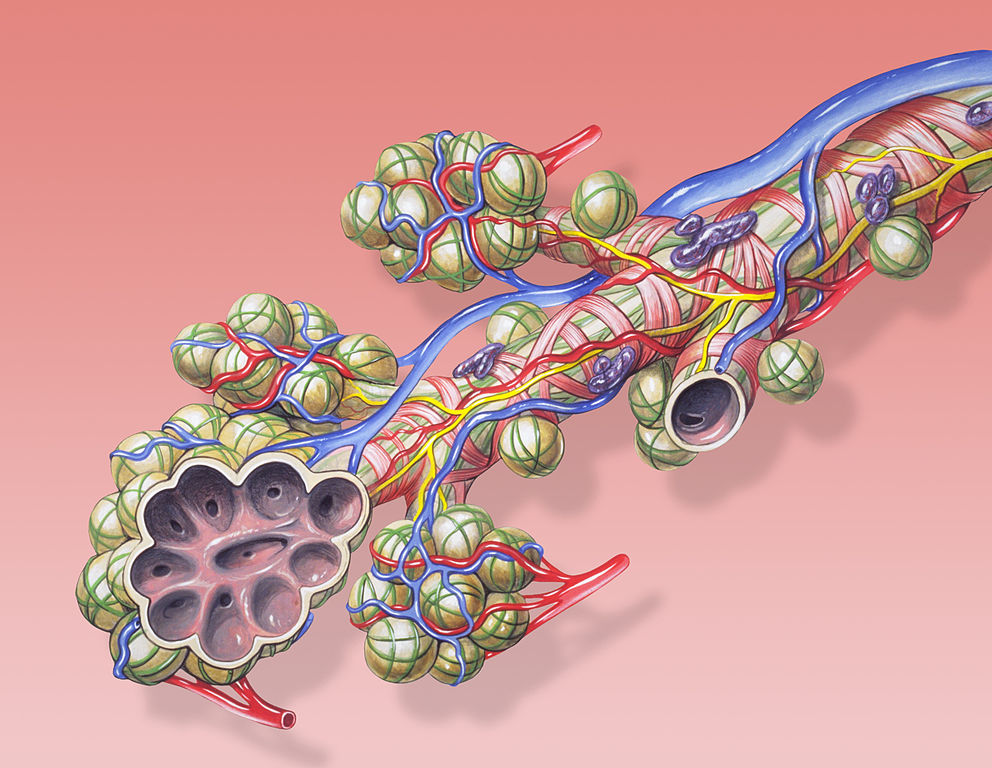
Схема оконечности бронхиолы и расположенных на ней альвеол. Красным показаны артериальные, синим — венозные капилляры, фиолетовым — железы, выделяющие слизь в просвет бронхов, желтым — ветви нервов.
Альвеолярная структура легких значительно увеличивает их дыхательную поверхность, которая, например, у человека составляет порядка 100 кв. м при общей поверхности тела около 1 кв. м. При этом относительная величина респираторной поверхности больше у активных и подвижных млекопитающих. Так, если у ленивца легкие заключают около 5 млн. альвеол, то у кошки их около 400 млн. Подобный разброс неудивителен — дыхание медлительного ленивца гораздо более вяло, чем у деятельной кошки. При этом дыхательная поверхность достигает у ленивца (Bradypus tridactylus) 6 кв. см на 1 г массы тела, у кошки (Felis silvestris) — 28 кв. см на 1 г, у мыши (Mus musculus) — 54 кв. см на 1 г, у подковоноса (Rhinolophus ferrumequinum) — 100 кв. см на 1 г. Площадь респираторной поверхности увеличивается не только за счет увеличения размера легких, но и за счет усложнения их внутренней дифференцировки.
Таким образом, форма и величина легких в значительной степени связаны с образом жизни животных: малоподвижные крупные формы имеют однодольчатые, относительно небольшие легкие, наоборот, высокоподвижные мелкие животные характеризуются относительно большими многодольчатыми легкими. Относительное увеличение легких наблюдается у высокогорных и водных млекопитающих. У китов существует кольцеобразная мускулатура, запирающая входы в альвеолы и позволяющая задерживать в них воздух даже на больших глубинах.
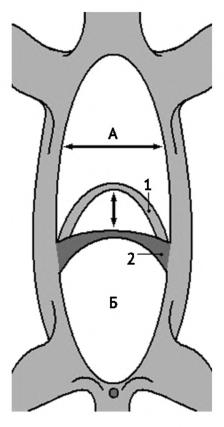
Брюшное дыхание млекопитающего: А — грудная полость; Б — брюшная полость; 1 — диафрагма расслабленна; 2 — диафрагма сокращена; горизонтальной стрелкой показаны движения ребер, вертикальной — движения диафрагмы.
Дыхание млекопитающих выполняется с помощью очень эффективного всасывающего механизма. Расширение грудной полости отчасти происходит за счет движения ребер вверх и наружу, осуществляемых лестничной мышцей (musculus scalenus). Однако большее значение имеет движение диафрагмы назад или вниз. В расслабленном состоянии диафрагма имеет форму купола, обращенного вершиной к голове; при сокращении она уплощается. Каждое легкое свободно лежит внутри своей плевральной полости, прикрепляясь только переднемедиальной стороной, через которую в него входят бронх и кровеносные сосуды. Дыхательные движения расширяют грудную полость и, следовательно, плевральные полости; при их расширении легкие втягивают воздух, чтобы заполнить появившийся дополнительный объем. При расслаблении диафрагмы собственная упругость и возросшее давление в брюшной полости заставляют ее занять исходное положение. Таким образом, давление в грудной полости изменяется благодаря работе двух механизмов: движениям ребер, изменяющим ширину грудной полости, и движениям диафрагмы, изменяющим ее глубину. В разном сочетании оба механизма функционируют у всех млекопитающих; у хищников преобладает первый, у копытных — второй. В любом случае, вентиляция легких млекопитающих очень интенсивна, а их внутренняя структура позволяет достигнуть уровень газообмена, по эффективности уступающего только птичьему. И все-таки этот уровень является вполне достаточным для поддержания высокой скорости обмена веществ, а за счет нее — постоянной температуры тела млекопитающих.

Бегущая импала (Aepyceros melampus). Ускорения и замедления при каждом прыжке копытных заставляют содержимое их брюшной полости двигаться внутри тела взад - вперед, дополнительно воздействуя на диафрагму и помогая дыханию.
Число дыхательных движений находится в зависимости от величины животного, определяющей ту или иную интенсивность обмена веществ; чем больше отношение поверхности тела к его массе, тем чаще дыхание. Так, у лошади (Equus ferus) число дыханий за 1 мин. равно 8-16, у человека (Homo sapiens) — 15-20, у черного медведя (Ursus americanus) — 15-25, у лисицы (Vulpes vulpes) — 25-40, у крысы (Rattus norvegicus) — 100-150, у мыши (Mus musculus) — около 200. Кроме того, частота дыхания зависит от физического состояния животного: в покое она уменьшается, а при движении возрастает. В указанной связи различна и интенсивность потребления кислорода.
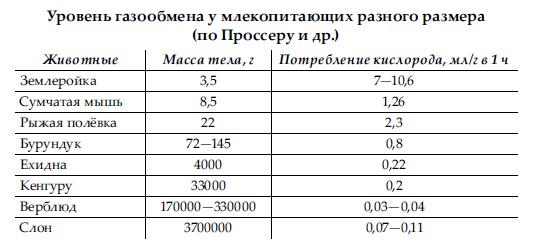
Приведенные в таблице данные также свидетельствуют о более низком уровне обмена у однопроходных и сумчатых по сравнению с плацентарными, что соответствует и примитивности их общей организации. Для сравнения укажем, что у чешуйчатых рептилий этот показатель равен всего 0,12-0,3, а у амфибий — 0,012-0,075.
Легкие, по существу, являются единственным органом дыхания млекопитающих. В газообмене также участвует слизистая поверхность воздухоносных путей и частично кожа, однако роль их ничтожна: лишь около 1 % потребляемого кислорода поступает через кожные кровеносные сосуды. Сказанное понятно, если учесть ороговение эпидермиса и незначительную общую поверхность кожи сравнительно с суммарной дыхательной поверхностью легких, которая превышает ее в 50-100 раз. Вентиляция легких не только обусловливает газообмен, но и имеет существенное значение для терморегуляции.
Управляет легкими дыхательный центр, расположенный в продолговатом мозге. Повышение в крови (а значит, и в спинномозговой жидкости) концентрации углекислого газа, сопровождающееся образованием угольной кислоты, влечет возрастание концентрации ионов водорода. Они раздражают нервные клетки, которые и дают сигнал межреберным мышцам усилить дыхательные движения. Наоборот, щелочная реакция крови дыхание тормозит.
вперед
в оглавление