назад
Гигантизм
Одной из замечательных черт эволюции фактически всех основных групп позвоночных была тенденция к прогрессивному увеличению размеров тела вплоть до гигантизма, каждый раз оказывавшегося эволюционным тупиком. Эта закономерность в полной мере проявилась и у млекопитающих. Чуть ли не на протяжении всего мезозоя млекопитающие были мелкими и даже очень мелкими животными, размером от землеройки до кошки. Переход ранних млекопитающих из мелкоразмерного класса в крупноразмерный начался в конце мела - начале палеогена и был связан, очевидно, с вымиранием гигантских рептилий. В разных группах эта общая тенденция реализовалась несколько различным образом.
Так, в истории южноамериканских сумчатых (Ameridelphia) весьма примечательно формирование группы крупных и даже очень крупных (размером до медведя) плотоядных, которых в настоящее время выделяют в отдельный отряд спарассодонтов (Sparassodonta). Они просуществовали на протяжении большей части кайнозоя и дали значительное разнообразие форм (3-5 семейств, более 30 родов), в том числе и саблезубых (в семействе боргиен (Borhyaenidae)). Поскольку крупные травоядные сумчатые в Новом Свете не известны, очевидно, спарассодонты хищничали на плацентарных — неполнозубых и эндемичных копытных.
В Австралии, напротив, переход сумчатых в крупный размерный класс был связан именно с растительноядной специализацией: в отряде двурезцовых (Diprotodontia) сформировались пастбищные животные размером от овцы до носорога (в семействе дипротодонтов (Diprotodontidae)). Крупных автохтонных хищников почти не было, если не считать волкоподобных тилацинов (Thylacinus); своего рода курьез представляют «сумчатые львы» (Thylacoleonidae): они принадлежат к двурезцовым сумчатым, но строение их щечных зубов позволяет предполагать хищничество.
Среди плацентарных, пожалуй, лишь анагалиды (Anagalida), липотифли (Lipotyphla) и архонты (Archonta) остались почти целиком мелкоразмерными. Среди них гигантизм свойствен только отдельным растительноядным представителям: некоторым приматам (мегаладаписам (Megaladapis), гигантопитекам (Gigantopithecus), гориллам (Gorilla) и некоторым другим), а также немногим вымершим грызунам (из числа диномисов (Dinomyidae), водосвинок (Hydrochoeridae) и бобров (Castoridae)).
Во многих других филетических линиях эвтериев тенденция к гигантизации преобладала или хотя бы была отчетливо выражена. В первую очередь это было связано с развитием растительноядности; но вслед за появлением крупных жертв не заставляли себя ждать и крупные хищники.
Одну из таких линий, исторически, видимо, первую, представляют ранне-среднепалеогеновые цимолесты (Cimolesta). Относящиеся сюда подотряды тэниодонтов (Taeniodonta), тиллодонтов (Tillodontia) и пантодонтов (Pantodonta) стартовали из мелкого размерного класса всеядных млекопитающих и в каждом из них возникали гигантские животные размером с носорога, причем пантодонты достигали значительного разнообразия (более десятка семейств). Еще одна группа эвтериев, в разных ветвях которой нередко появлялись крупные растительноядные формы (все они вымерли), — американские ксенартры (Xenarthra).
Однако более всего примечательны копытные в широком смысле — унгуляты (Ungulatа), которым названная тенденция особенно свойственна — и по выраженности соответствующих адаптаций, и по частоте их появления. В самых разных филетических линиях они дали очень крупные растительноядные формы: это были исходно азиатские диноцераты (Dinocerata), непарнопалые (Perissodactyla) и парнопалые (Artiodactyla), африканские даманы (Hyracoidea), эмбритоподы (Embrithopoda) и хоботные (Proboscidea), а также вышеперечисленные южноамериканские эндемичные копытные. Парацератерии (Paraceratherium) из семейства гиракодонтов (Hyracodontidae) достигали 8,5 м в длину, почти 6 м в высоту, а весили, судя по косвенным признакам, до 20 тонн — это были самые крупные известные наземные млекопитающие (вес наибольших современных слонов примерно втрое меньше).
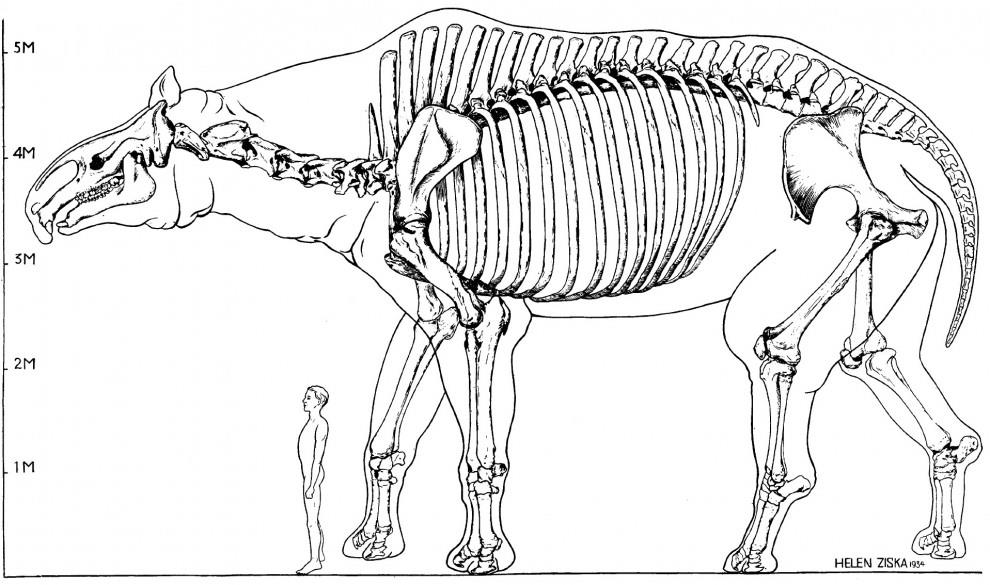
Сравнительные размеры парацератерия (Paraceratherium transouralicum) и человека.
Особенно крупные плотоядные эвтерии также возникали неоднократно, но в составе немногих отрядов. В двух из них — гиенодонтах (Hyaenodonta) и хищных (Carnivora) — это были типичные хищники. Гигантскими были и многие плотоядные мезонихии (Mesonychia).
Разумеется, особого упоминания заслуживают морские млекопитающие, среди которых гигантизм — не исключение, а правило. Таковы, в первую очередь, китообразные (Cetacea), а также сирены (Sirenia), десмостилии (Desmostylia) и ластоногие (Pinnipedia): масса их представителей составляет от нескольких тонн у самых крупных ластоногих и десмостилий до сотни и более тонн у некоторых усатых китов (Mysticeti).
При увеличении веса в конструкции опорной системы наземного животного происходят различные изменения. С возрастанием массы тела, например, выпрямляются углы в отдельных суставах конечностей, и тогда сила веса приближается к направлению оси соответствующих частей конечности, разгружая, таким образом, мускулатуру и нагружая скелет конечностей. Поэтому у животных с большим весом таз прикрепляется к крестцу более отвесно, причем положение тазобедренного сустава приближается к вертикали, опущенной из крестцового сочленения (ослабление тенденции к повороту); седалищная часть таза «преломляется» назад; происходит ряд других изменений, приводящих к уменьшению нагрузки наиболее напряженных костей скелета.
Увеличение телесных размеров влечет за собой утяжеление скелета, грузоподъемность которого возрастает примерно на один порядок медленнее, чем масса мускулатуры и остальных органов. Результатом этого и бывает абсолютное и относительное утолщение костей у крупных животных, ведущее к увеличению доли скелета в общей массе тела. У мелких млекопитающих (например, у обыкновенной бурозубки (Sorex araneus)) доля скелета от общей массы тела составляет около 8 %, а у крупных зверей (таких, как гиппопотам (Hippopotamus amphibius), саванный слон (Loxodonta africana)) достигает примерно 20 %. Несмотря на утолщение костей, скелет многих крупных млекопитающих более или менее облегчен за счет пневматизации.
Путем компромисса между увеличением размеров тела и торможением нарастания массы отдаляется достижение предельных величин утяжеления. В другом случае может развалиться вся опорная система животного. Утолщение костей в свою очередь вызывает необходимость развития мускулатуры, а так как прочность кости определяется ее диаметром и рост ее происходит медленнее, чем наращивание мышечной массы, то упрочение опорной системы в конце концов должно иметь определенные пределы из-за чрезмерного утяжеления. Критический порог увеличения массы у сухопутных животных находится при весе около 25 тонн. Специальные исследования костей уже упомянутых парацератериев свидетельствуют о том, что они выдерживали силу сжатия в 250 тонн. Таким образом, для 25-тонного животного запас прочности составлял 10-кратную величину. Подобный запас прочности характерен и для животных с меньшей массой тела. Однако при больших физических нагрузках не выдерживают в первую очередь сухожилия, связки и суставы, а не кости. Это и есть главная причина, почему наиболее тяжелые позвоночные — киты — живут в воде. Здесь их вес облегчается силой, приблизительно равной их массе, чем значительно уменьшаются требования к опорной системе. Когда киты выбрасываются на берег, они погибают прежде всего из-за того, что их раздавливает собственный вес.

Наиболее крупное современное млекопитающее — синий кит (Balaenoptera musculus) (до 33 м и более 150 т) рядом с самым мелким — этрусской землеройкой (Suncus etruscus) (от 6 см и 1,2 г).
Большие размеры обеспечивают их носителям целый ряд выгод. Крупное животное, следовательно, физически сильное, имеет меньше врагов и находится ближе к вершине пищевой пирамиды. Поэтому оно может позволить себе роскошь одиночества. Одиночный образ жизни предполагает отсутствие пищевых конкурентов в ближайшем окружении, что облегчает поиски больших количеств пищи. Большие размеры тела позволяют животным совершать длительные миграции с минимальными затратами энергии в поисках мест скопления пищевых ресурсов. Крупные животные живут дольше, чем мелкие. Наконец, большие размеры тела формируют определенные метаболические особенности организма, что, в свою очередь, создает предпосылки для жизни как в условиях высоких температур, так и в полярных районах с коротким летом и низкими среднегодовыми температурами.
С другой стороны, у мелких животных также имеется ряд преимуществ. Небольшому зверьку проще отыскать корм в ближайшем окружении и насытится его малым количеством. Высокая напряженность метаболизма мелких животных обусловливает их способность в короткий промежуток времени совершить полный жизненный цикл от образования гамет до формирования половозрелой особи. За 3-летний период жизни 30-граммовая мышь способна произвести на свет в 100 раз больше потомков, чем 5-тонный слон за всю его продолжительную жизнь. При такой большой численности у мелких животных гораздо меньший риск сокращения популяции с появлением неблагоприятных факторов. Уже поэтому виды мелких животных легче переживают различные природные катастрофы. Кроме того, их высокая численность обеспечивает большую вероятность появления благоприятных в данных условиях наследственных отклонений — мутаций.
В свете сказанного можно считать, что увеличение размеров тела является одной из форм узкой специализации. Новые виды чаще возникают от сравнительно примитивных форм, малоспециализированных, обладающих большим числом самых общих признаков, которые могут обеспечить более быстрое приспособление к изменяющимся условиям среды, чем от видов, высокая специализация которых приводит в тупик одностороннего приспособления.
вперед
в оглавление