в оглавление
назад
Летающие формы
Полетом обычно считают движение в воздухе, при котором преодолевается расстояние, превышающее возможности прыжка, совершаемого за счет собственных мышечных усилий. Не вызывает сомнений, что летающие млекопитающие возникли из древесных форм путем развития их способности к прыжкам, затем — к пассивному полету (планированию) и только в конечном итоге — к активному полету с помощью крыльев. Этот ряд можно наблюдать и при обзоре современных видов.
Перепрыгивание с ветки на ветку и преодоление таким способом довольно значительных расстояний не является у лазающих животных чем-то необычным. Так, как говорилось выше, обыкновенная белка (Sciurus vulgaris) и соня-полчок (Glis glis) при прыжке широко расставляют лапы, увеличивая тем самым поддерживаемую воздухом площадь тела. У колобусов (Colobus) для этого служат очень длинные волосы по бокам туловища, развевающиеся во время прыжков. Длинный пушистый хвост всех этих зверьков играет роль парашюта.

Прыжок самки восточного колобуса (Colobus guereza) вместе с детенышем.
К полувоздушным формам могут быть отнесены млекопитающие с летательной перепонкой — кожистой складкой, которая тянется по бокам туловища между передними и задними конечностями, предоставляющей возможность полноценного планирования. Планирующие формы возникали неоднократно в самых разных филетических линиях. Древнейший известный их пример — позднеюрский волатикотерий (Volaticotherium antiquum) из докодонтов (Docodonta). Среди двурезцовых сумчатых (Diprotodontia) подобный тип передвижения появлялся не менее трех раз (сумчатые летяги (Petauridae), кольцехвостые кускусы (Pseudocheiridae), перьехвостые кускусы (Acrobatidae)), дважды или трижды среди грызунов (летяги (Petauristinae), большинство шипохвостов (Anomaluridae)), а также у шерстокрылов (Dermoptera).
У сумчатых летяг (Petauridae) кожные складки натянуты между туловищем и конечностями, от кистей до стоп. Более сложное строение имеет складка настоящих летяг (Petauristinae), дополнительно поддерживаемая с каждой стороны особой тонкой длинной косточкой, отходящей от запястья назад и в сторону, а у африканских чешуйчатохвостых летяг (Anomalurus) из Anomaluridae — хрящевым выступом локтевого сгиба. Наибольшего развития кожная складка достигает у шерстокрылов (Cynocephalidae), у которых она распространяется по бокам тела почти от ушей и до кончика хвоста, прикрепляясь на конечностях ко вторым фалангам пальцев. При прыжках передние конечности планирующих форм вытягиваются далеко вперед и в стороны, а задние — назад, при этом кожа растягивается, увеличивая несущую поверхность.
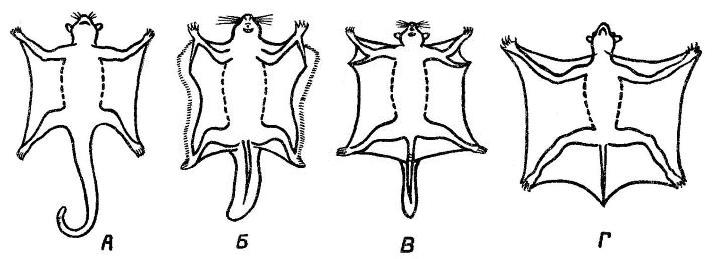
Представители различных групп млекопитающих, обладающие приспособлениями к планирующему полету: А — большая сумчатая летяга (Petaurus australis); Б — обыкновенная летяга (Pteromys volans); В — гигантский шипохвост (Anomalurus pelii); Г — филиппниский шерстокрыл (Cynocephalus volans).
Полувоздушные млекопитающие пользуются планированием не только для спуска по наклонной линии, но и для поворотов в воздухе и даже для незначительных подъемов. Так, обыкновенная летяга (Pteromys volans) обычно заканчивает свой прыжок-полет несколько взмывая кверху, что смягчает ее толчок о ствол дерева. Направление и угол полета меняется в результате поднятия, опускания или раздвигания конечностей и движений пушистого хвоста. Посредством планирования эти животные переносятся на десятки метров; например, сумчатые летяги рода Petaurus без труда преодолевает по воздуху расстояние до 40 м, настоящие летяги (Petauristinae) — до 60 м, гигантский летучий кускус (Petauroides volans) — до 100 м, а филиппинский (Cynocephalus volans) и малайский (Galeopterus variegates) шерстокрылы — и до 140 м. При этом последние способны не только планировать при спуске по отлогой линии со скоростью порядка 100 м/мин, но и совершать повороты в воздухе. За счет таких локомоций эти млекопитающие уходят от хищников, находят половых партнеров и осваивают новые кормовые угодья, не спускаясь на землю. При перемещении по древесным стволам кожные перепонки складываются во избежание порезов и проколов сучками.
Примечательно, что планирующие млекопитающие известны только в Старом Свете (включая Австралию). В Америке подобные жизненные формы не возникли — вероятно, потому что среди тамошних сумчатых и автохтонных грызунов почти нет типично древесных форм, а широконосые обезьяны к этому способу передвижения не прибегли.

Стадии планирующего полета южной американской летяги (Glaucomys volans).
В отличие от архозавров, у которых активный полет возникал как минимум дважды (птерозавры и птицы), в истории млекопитающих переход в воздушную среду произошел, по-видимому, только один раз — у представителей монофилетического отряда рукокрылых (Chiroptera). Приспособленность к полету позволяет им длительное время, быстро и с большой маневренностью передвигаться в воздухе, добывать здесь пищу и даже спариваться. Характерная особенность этой большой группы млекопитающих — превращение передних конечностей в длинные и гибкие крылья. Они устроены совсем не так, как крылья птиц, у которых маховые перья крепятся к костям предплечья. Основную площадь крыла рукокрылых составляет летательная перепонка (patagium), образованная эластичной кожной складкой, растянутой в области пясти и на четырех сильно удлиненных пальцах, кроме I-го, который остается свободным и вооружен крупным когтем для лазания (у крыланов (Pteropodidae) признаки самостоятельности сохраняет и II-й палец). Помимо передних конечностей, летательная перепонка охватывает бока туловища, а ее задний край крепится в области голеностопного сустава или же захватывает хвост. В качестве тормозного органа служит межбедренная часть летательной перепонки, поддерживаемая особой костной или хрящевой шпорой (calcar). Задние конечности необычным образом вывернуты, так что колени обращены назад и вверх — в связи с адаптацией к способу подвешивания на отдыхе вниз головой. Короткие пальцы стопы почти одинаковы по длине и вооружены острыми когтями.
У присосконогих летучих мышей — американских (Thyropteridae) и мадагаскарских (Myzopodidae) — у основания I-го пальца передних конечностей и на подошве задних конечностей имеются своеобразные округлые присасывательные диски. С их помощью эти зверьки прикрепляются к гладкой поверхности листьев во время дневного отдыха. Присасывательная сила присосок настолько велика, что животное может удерживаться всего на одной присоске.

Присасывательный диск на передней конечности трехцветного присосконога (Thyroptera tricolor).
У рукокрылых хорошо развиты грудные (летательные) мышцы, прикрепляющиеся у большинства видов к небольшому килю грудины. В отличие от птиц, которые пользуются для полета двумя парами мощных мышц, рукокрылым для этой цели служат четыре пары больших и несколько пар мелких мышц. У многих видов плечевая кость несет большой бугор, образующий дополнительный сустав с лопаткой, что предохраняет крыло от вывиха при резких его движениях во время полета. Кроме того, с воздушным образом жизни связано раннее срастание костей черепа, быстрое окостенение скелета, слияние некоторых элементов грудной клетки, мощное развитие ключиц и своеобразный механизм дыхания. Дыхательные движения грудной клетки во время полета производятся с помощью особых вспомогательных мускулов (а не межреберных мышц, как у других млекопитающих); движения грудной клетки также связаны с работой крыльев. Полет требует большой затраты энергии, с чем связаны крупные относительные размеры сердца. Сердечный индекс летучих мышей достигает 10-14,4 %, т. е. мало уступает сердечному индексу быстро летающих птиц (у ласточек, например, величина сердечного индекса равняется 14,7 %).

Основные типы полета рукокрылых: I — машущий; II — порхающий; III — планирующий.
Несмотря на различия в строении крыльев, биомеханика движений рукокрылых в воздушной среде принципиально не отличается от таковой птиц. Полет чаще всего машущий гребного типа, когда оба крыла опускаются сначала вперед и вниз, а потом поднимаются назад и вверх. Крылья при этом двигаются относительно медленнее и с меньшим размахом, чем у птиц сходных размеров. Возможны также зависающий (трепещущий) и планирующий (но не парящий) виды полета. Первый из них используется при необходимости оставаться на одном месте, как бы подвешенными в воздухе, и связан с особенно большим расходом энергии; второй, наоборот, является наиболее экономным. Быстрота и изворотливость полета зависят от формы крыльев. Лучшими летунами являются виды, имеющие узкие и длинные крылья. Таковы бульдоговые летучие мыши (Molossidae), в частности, бразильский складчатогуб (Tadarida brasiliensis): его полет — легкий и стремительный, причем средняя скорость полета составляет 40 км/ч, а максимальная — 105 км/ч. Обладатели широких и тупых крыльев летают сравнительно медленно, со средней скоростью около 15 км/ч; полет их неровный, порхающий. Крупные крыланы значительно уступают в легкости и подвижности мелким летучим мышам, но способны к более продолжительному непрерывному полету.

Разнообразие экологии рукокрылых. Типичный представитель летучих мышей — мексиканский большеух (Corynorhinus mexicanus) (1) питается насекомыми. Большой зайцегуб (Noctilio leporinus) (2) ловит рыбу с поверхности воды. Обыкновенный вампир (Desmodus rotundus) (3) прорезает кожу тапира и слизывает кровь. Землеройковидный листонос (Glossofaga soricina) (4) питается нектаром и пыльцой, а складчатомордый листонос (Centurio senex) (5) предпочитает сочные фрукты.
В случае необходимости многие рукокрылые могут довольно быстро передвигаться по земле, действуя обеими парами конечностей, лазать по отвесным поверхностям деревьев и пещер и даже плавать; способностью совершать вертикальный взлет с земли они превосходят птиц. Активны преимущественно ночью или в сумерки, ориентируются с помощью зрения и обоняния (фруктоядные крыланы) или активной эхолокации (насекомоядные летучие мыши). Нектароядные летучие мыши опыляют ночные цветки точно так же, как колибри — дневные. Малая предельная масса тела рукокрылых (1,6 кг, что на порядок меньшая, чем у летающих птиц), видимо, определяется не пороками их морфологии и физиологии, а необходимостью сохранения огромного резерва мощности для обеспечения полноценных летных качеств беременной самки.
вперед
в оглавление