в оглавление
назад
Кожная, сухожильно-мышечная и внутриорганная чувствительность
Кожная чувствительность делает возможным восприятие внешних воздействий различного характера. Ее рецепторами служат как свободные нервные окончания, так и специфические чувствительные структуры. Последние сильно различаются по строению; одни из них представляют собой группы видоизмененных эпителиальных клеток, оплетенные сетью нервных волокон, другие являются разветвленными окончаниями нервов, упакованными в глиальные колбы разной формы. Данные образования могут быть либо некапсулированными, либо заключенными в соединительнотканные капсулы. Иногда они бывают настолько велики, что их можно рассмотреть невооруженным глазом. Чувствительный аппарат особенно хорошо развит на пальцах лазающих млекопитающих и человека, на подушечках лап хищных, конце хобота слона, рыльце крота и т. д.
Многочисленные свободные нервные окончания располагаются в эпидермисе и, реже, в дерме; они выступают в качестве терморецепторов, механорецепторов и ноцицепторов, отвечающих соответственно за восприятие температуры, механических воздействий и болевых ощущений. Некапсулированные чувствительные структуры представлены дисками Меркеля, относящимися к рецепторам давления; в отличие от птиц, у которых они локализуются в дерме, у млекопитающих эти тельца лежат в глубоких слоях эпидермиса. Особенно разнообразны структуры, заключенные в капсулы. Тельца Мейснера, находящиеся в сосочковом слое дермы, воспринимают тактильные ощущения и поверхностное давление. Тельца Гольджи-Маццони, залегающие в дерме, и тельца Фатера-Пачини, расположенные преимущественно в подкожной жировой клетчатке, отвечают за восприятие глубокого давления и вибрации. Тельца Руффини, совмещающие функции рецепторов растяжения и тепла, находятся глубоко в дерме и в подкожной клетчатке. Колбы Краузе, реагирующие на холод, располагаются в сосочковом слое дермы, причем их в несколько раз больше, чем тепловых рецепторов. Кроме того, колбы Краузе залегают в соединительнотканной основе слизистых оболочек и среди мышечных волокон языка. По некоторым данным, одни и те же рецепторы в некоторых случаях могут воспринимать различные виды раздражений.

Некоторые кожные рецепторы млекопитающих: а — свободные нервные окончания; б — тельце Мейсснера; в — колба Краузе; г — тельце Фатера-Пачини; д — тельце Руффини.
Некоторые кожные рецепторы отличаются быстрой сенсорной адаптацией. В ответ на ступенчатое изменение стимуляции частота нервных импульсов быстро повышается, а затем снижается до уровня покоя. Это значит, что рецептор служит хорошим индикатором изменений в силе стимуляции, но плохим индикатором ее абсолютного уровня. Это дает преимущества в тех случаях, когда от кожных рецепторов требуется быстрая информация об изменениях среды, которые могут подействовать на организм, например об изменениях температуры. Плотность расположения кожных рецепторных структур неравномерна. Например, к температурным изменениям и давлению более чувствительны конечности и морда животного и менее чувствительно туловище.
Нервные волокна от кожных рецепторов уходят через верхние корешки спинного мозга в серое вещество его верхних рогов. Отсюда через переднюю комиссуру по спиноталамическому восходящему пути афферентные потоки поступают в таламические ядра противоположной стороны. В таламусе пути потоков от различных рецепторов расходятся. Здесь же осуществляется фильтрация сенсорной информации. Ее дальнейшая судьба связана с гипоталамическими ядрами, а также корой больших полушарий. Болевая афферентация также попадает в продолговатый мозг, на ретикулярную формацию, в структуры среднего мозга и лимбической системы.
Важную роль в восприятии прикосновений и отчасти положения тела в пространстве играют волосы и особенно длинные жесткие вибриссы. Их основания окружены сложно извитыми нервными окончаниями, на которые волоски и вибриссы действуют подобно рычагам для нажатия, передавая информацию о малейшем отклонении от исходного положения; при этом они реагируют не только на прикосновения, но и на колебания воздуха. Вибриссы располагаются на участках тела, чаще приходящих в соприкосновение с окружающими предметами. Наибольшего развития они обычно достигают на морде, где располагаются пучками в строго определенных местах — на верхней губе, на щеках, в составе бровей над глазами, на подбородке. У норников, обладающих широкой поверхностью соприкосновения со стенками норы, вибриссы помимо головы разбросаны по всему туловищу. У лазающих форм (например, у белок и лемуров) они также расположены на брюшной поверхности и на частях конечностей, контактирующих с субстратом при передвижении по деревьям, у кошек — на передних лапах и локтях. Вибриссы помогают животному ориентироваться на местности и находить пищу в условиях плохой видимости.

Схема расположения вибрисс на морде кролика (Oryctolagus cuniculus) с очерченной пунктиром «осязательной зоной».
У летучих мышей нижняя сторона крыльев усеяна крошечными осязательными волосками, помогающими им улавливать воздушные завихрения. Удалив волоски с помощью депиляционного крема, исследователи наблюдали за поведением животных в воздухе. Когда представителям одного из видов этих зверьков нужно было сделать поворот на 90 °, чтобы не наткнуться на находящееся на пути препятствие, вертикально взмыть вверх они не смогли. Некоторые зверьки во время полета регулярно натыкались на потолок. Совершать безопасные полеты летучие мыши смогли только тогда, когда на их крыльях снова отросли волоски.
У однопроходных вибриссы отсутствуют, однако имеются преобразованные из кожных желез рецепторы, чувствительные к электрическому полю. Из-за способности воды проводить электричество соответствующие рецепторы особенно хорошо развиты у кормящегося в водоемах утконоса (Ornithorhynchus anatinus). Порядка 40 000 их располагается на клюве и воспринимает электрические импульсы, порождаемые работающими мышцами рачков, мелкой рыбы, головастиков и другой добычи. Помимо электрочувствительных, на клюве утконоса имеются и тактильные рецепторы, улавливающие движение воды. Когда утконос охотится, он поводит головой из стороны в сторону, чтобы определить направление, в котором нужно искать добычу, и расстояние до нее. Проехидна (Zaglossus bruijni), живущая во влажном тропическом лесу, имеет на рыльце более 2000 электрорецепторов, которые она использует для выслеживания земляных червей и другой обитающей в почве добычи. Австралийская ехидна (Tachyglossus aculeatus) населяет более сухие места, и имеет на кончике рыльца всего около 400 электрорецепторов. Они служат для обнаружения добычи в сырую погоду, например когда идет дождь.
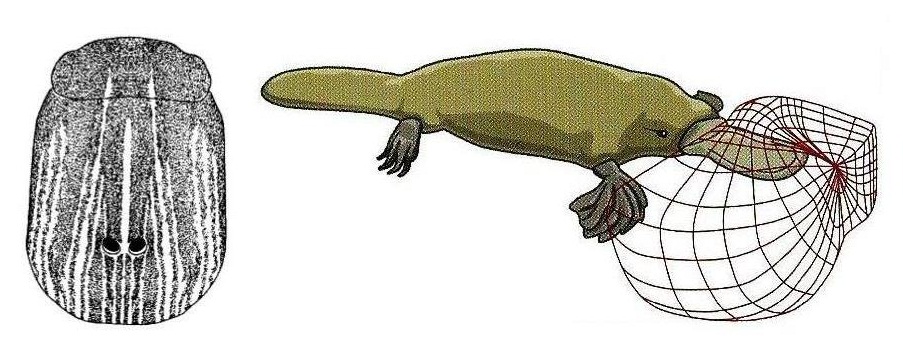
Слева — электрорецепторы (показаны в виде черных точек) на клюве утконоса (Ornithorhynchus anatinus).
Справа — чувствительная зона вокруг клюва охотящегося утконоса (Ornithorhynchus anatinus).
Среди других млекопитающих электрорецепторы выявлены у гвианского дельфина (Sotalia guianensis). Они располагаются в небольших ямках на рыле, в которых некогда находились вибриссы и, по-видимому, являются производными их луковиц. Эти органы фиксируют электросигналы, которые исходят от мелких морских обитателей, которыми обычно питается гвианский дельфин. Это животное обитает в прибрежных водах Южной Америки, где большое количество взвешенных частиц делает воду мутной и не позволяет обнаружить добычу с помощью зрения, а эхолокация на небольшом расстоянии не очень эффективна. Вполне вероятно, чувствительность к электрическим сигналам в той или иной степени свойственна всем дельфинам, хотя специфичные органы пока обнаружены только у указанного вида. Во всяком случае, этим можно объяснить, почему некоторые дельфины любят рыхлить придонный грунт в поисках пищи.

Электрорецепторы (отмечены стрелкой) на рыле гвианского дельфина (Sotalia guianensis).
Проприоцептивная чувствительность обеспечивается сенсорными структурами, расположенными в поперечнополосатых мышцах, сухожилиях и суставах — соответственно мышечными веретенами, сухожильными рецепторными органами Гольджи и суставными рецепторами. Чувствительные структуры первых двух типов являются основой «мышечного чувства». Они поставляют центральной нервной системе сведения о степени сокращения, напряжения или растяжения мышц. Эти данные необходимо передать прежде, чем по двигательным нервам мышце будет отдана следующая правильная команда (принцип обратной связи). Вместе с тем мышечные веретена участвуют в простом рефлексе, который противодействует удлинению мышцы, а сухожильные органы Гольджи — в простом рефлексе, противодействующем повышению мышечного напряжения. Кроме того, мышечные веретена являются для животного источником информации о положении в пространстве различных частей его тела. Суставные рецепторы дают информацию об угловом положении каждого сустава. Вся эта информация может быть получена без помощи других сенсорных структур и исходит (в отсутствие контактов с другими объектами) от частей тела, в которых имеются суставы, поперечнополосатые мышцы и их сухожилия.
Мышечные веретена состоят из тонких специализированных мышечных волокон, вокруг средней части которых спирально закручено чувствительное нервное окончание, называемое первичным, или аннулоспиральным. При удлинении мышцы веретено растягивается и посылает быстрые сигналы в спинной мозг. Встречаются также вторичные гроздевидные окончания, посылающие более медленные сигналы. Во многих веретенах млекопитающих имеются и первичные, и вторичные окончания, а в других — только первичные. Снаружи мышечные веретена заключены в фузиформную соединительнотканную оболочку, и их мышечные волокна называются интрафузальными в противоположность обычным — экстрафузальным. Последние иннервируются альфа-мотонейронами, тела которых находятся в спинном мозге. Интрафузальные волокна иннервируются более мелкими гамма-мотонейронами, поддерживающими веретено в тоническом состоянии активности, благодаря чему для его активации требуется меньшее мышечное растяжение. Поскольку мышечные веретена расположены параллельно экстрафузальным волокнам, при сокращении мышцы они склонны к расслаблению. Гамма-нейроны могут приказать интрафузальным волокнам напрячься, в результате чего веретено сохранит состояние готовности.
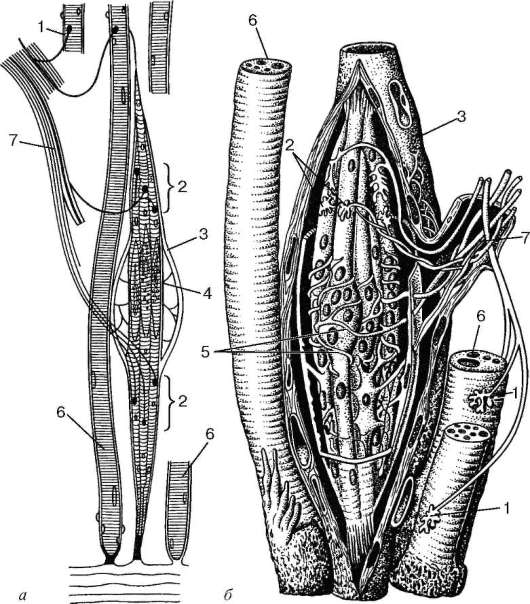
Схема строения мышечного веретена: а — моторная иннервация интрафузальных и экстрафузальных мышечных волокон; б — спиральные афферентные нервные окончания вокруг интрафузальных мышечных волокон в области ядерных сумок; 1 — нейро-мышечные эффекторные окончания экстрафузальных мышечных волокон; 2 — моторные бляшки интрафузальных мышечных волокон; 3 — соединительнотканная капсула; 4 — ядерная сумка; 5 — чувствительные кольцеспиральные нервные окончания вокруг ядерных сумок; 6 — скелетные мышечные волокна; 7 — нерв.
Интероцепторные тельца нескольких типов располагаются во внутренних органах и в промежутках между ними. Так, в брыжейке и соединительнотканных оболочках некоторых органов расположены уже упомянутые тельца Фатера-Пачини, воспринимающие глубокое давление и вибрации. Похожие рецепторы сигнализируют о наполнении желудка и мочевого пузыря. Рецепторы сосудов, принадлежащих к системе артериальных дуг (в частности, сонных артерий), регистрируют такие параметры, как давление крови и ее химический состав (содержание двуокиси углерода, солей, гормонов). В глубине тела (например, в венах) имеются рецепторы, фиксирующие температуры внутренней среды и способные вызывать дрожь, даже если температура кожных и мозговых рецепторов не изменяется. Терморецепторы спинного мозга влияют на дрожь, одышку и кровоток, причем эти же функции дублируются терморецепторами гипоталамуса. Повреждающие действия активируют различные виды интероцепторов, в том числе, вероятно, и специальные болевые (примечательно, что в самом мозге они отсутствуют). В организме имеются также рецепторы, реагирующие на изменение осмотического давления.
Информация, передаваемая большинством интероцепторов, не осознается и не вызывает ощущений. Иногда действия интероцепторов комбинируются, давая ощущения голода, жажды или тошноты, но это происходит вследствие сложных процессов в мозгу, которые не всегда связывают ощущение с определенными частями тела. Вероятно, это объясняется тем, что действие, которое должно быть произведено в ответ на голод и жажду, является гораздо менее непосредственным, чем ответ на прикосновение или поверхностное изменение температуры.
вперед
в оглавление