в оглавление
назад
Вкус и обоняние
Описанные выше простые чувства — это главным образом ответы на физические раздражители. Вкус и обоняние представляет собой ответы на химические раздражители, т. е. речь идет о хеморецепции. Ее механизмы включают распознавание специфических молекул рецепторными участками на клеточных мембранах.
Рецепторами вкуса служат небольшие вкусовые луковицы (caliculi gustatorii), которые имеют вид компактных скоплений продолговатых клеток эктодермального происхождения, погруженных в эпителий ротовой полости. В луковицах различают 2 типа клеток — чувствительные и окружающие их опорные. Кроме того, в клеточном слое присутствуют секреторные клетки, обеспечивающие луковицу слизью. Апикальная мембрана чувствительных клеток имеет несколько ресничек, выступающих из слизистого слоя. Реснички (волоски) обладают высокой чувствительностью к химическим веществам. Для того чтобы произошло раздражение чувствительной клетки, необходимо, чтобы вкусовое вещество вошло в контакт с вкусовой луковицей в составе водного раствора. У водных позвоночных в качестве растворителя выступает окружающая водная среда. У наземных животных функцию растворителя выполняет паротидная слюна — серозный секрет околоушных слюнных желез.
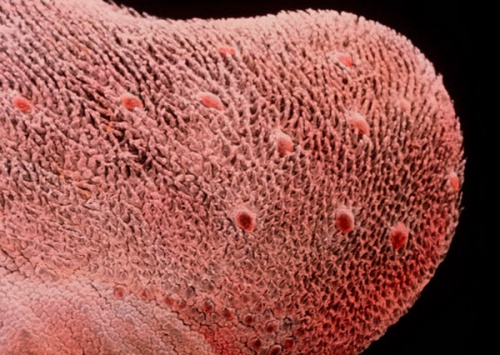
Микрофотография человеческого языка.
У млекопитающих вкусовые луковицы сконцентрированы в основном на языке, располагаясь на боковых поверхностях языковых сосочков (papillae linguales). Помимо этого, редкие луковицы имеются в слизистой оболочке губ, мягкого неба, глотки и надгортанника. Общее число луковиц у большинства млекопитающих невелико — десятки или сотни. Больше всего их у видов, долго удерживающих при жевании пищу в ротовой полости (например, у копытных и приматов). У кошки примерно 500 вкусовых луковиц, у насекомоядной летучей мыши — 800, у собаки — 1600, у человека — 9000, у свиньи — около 15000.
Внешне все вкусовые луковицы выглядят одинаково, но у тех животных, у которых они исследовались экспериментально, обнаружено несколько их типов, различающихся функционально: каждый из них реагирует на присутствие в пище определенных молекул или ионов. Большинство млекопитающих различают четыре основных вкуса — горький, соленый, кислый и сладкий. Хищные звери и человек также выделяют мясной вкус (т. н. «умами»). Первичный нейрон каждой чувствительной клетки имеет белок-рецептор, который избирательно соединяется со «своим» вкусовым веществом по принципу стереохимического сродства, в результате чего возникает специфическая афферентация. Вследствие ее координации в высших нервных центрах получается широчайшая палитра вкусовых ощущений.
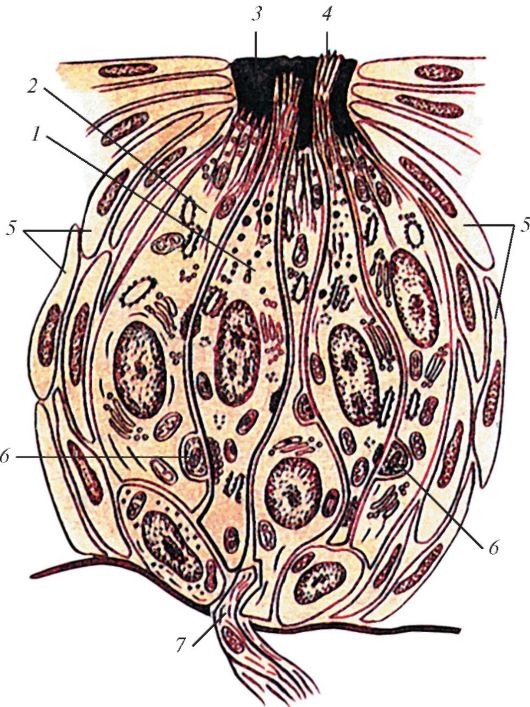
Схема строения вкусовой луковицы млекопитающих: 1 — вкусовая клетка; 2 — поддерживающая клетка; 3 — вкусовая пора; 4 — микроворсинки; 5 — эпителиальные клетки; 6 — нервное окончание; 7 — нервное волокно.
Традиционно считалось, что разные вкусы воспринимают различные участки языковой поверхности. Однако полученные молекулярные и функциональные данные показывают, что различные типы вкусовых луковиц распределены по всей поверхности языка и отличаются лишь плотностью своего распределения. Таким образом, никакой «карты языка» не существует, вопреки ошибочным популярным представлениям. Необходимо также отметить, что вкусовое восприятие в обыденном понимании на самом деле представляет собой преимущественно обонятельное восприятие содержимого рта и другие ощущения от пребывания пищи в ротовой полости. Именно поэтому пища кажется нам безвкусной, когда нос «заложен» во время насморка и пахучие частицы не могут попасть изо рта к обонятельным рецепторам в задней части носовой полости.
Со стороны базальной мембраны к хемочувствительным клеткам вкусовой луковицы подходят волокна лицевого нерва (передние 2/3 языка), языкоглоточного нерва (задняя 1/3 языка и твердое небо) и блуждающего нерва (глотка и надгортанник), оканчивающиеся во вкусовых долях (первичных вкусовых центрах) продолговатого мозга. От вагусных и вкусовых долей в продолговатый мозг отходят вторичные вкусовые тракты. От продолговатого мозга идут восходящие пути к крыше среднего мозга, переднему ядру таламуса и к ядрам гипоталамуса. Далее обработанный сигнал передается в соответствующую зону коры больших полушарий. В составе блуждающего, языкоглоточного и лицевого нервов обнаружены не только вкусовые волокна, но и волокна механорецепторов головы и ротовой полости, что обеспечивает корреляцию вкусовой и механической афферентации.
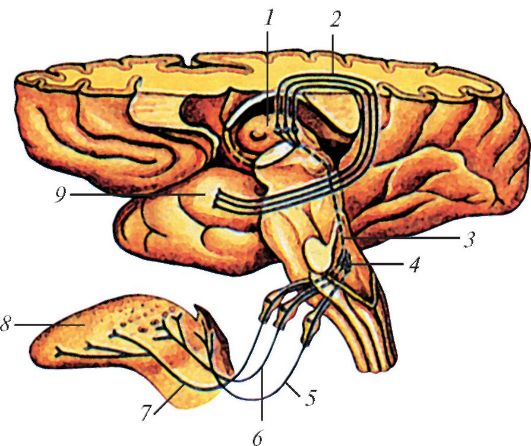
Проводящий путь органа вкуса млекопитающих на примере человека: 1 — таламус; 2 — волокна, соединяющие таламус и крючок; 3 — волокна, соединяющие ядро одиночного пути и таламус; 4 — ядро одиночного пути; 5 — вкусовые волокна в составе блуждающего нерва; 6 — вкусовые волокна в составе языкоглоточного нерва; 7 — вкусовые волокна в составе барабанной струны; 8 — язык; 9 — крючок.
Вкусовая привлекательность пищевых средств определяется многими причинами и в случае человека и животных имеет существенные различия. Более того, пищевые предпочтения этологически схожих животных (например, кошек и собак) не всегда совпадают. Так, собаки частенько отдают предпочтения несвежим продуктам, в то время как кошки таких продуктов избегают, а в случае их поедания страдают от отравления. Кошки в большей мере представляют группу плотоядных животных. Собаки же, будучи изначально охотниками, в тяжелых обстоятельствах превращаются во всеядных животных и демонстрируют склонность к копрофагии, включая автокопрофагию.
Вкусовая чувствительность животных изменяется на протяжении онтогенеза. Например, у собак вкусовая чувствительность возникает еще в дородовой период. Однако пренатальная вкусовая сенсорика специфична. Она настроена на восприятие химических веществ, циркулирующих в крови матери и способных преодолеть плацентарный барьер и амниотическую жидкость. В конце онтогенеза вкусовая чувствительность животных, как и чувствительность всех остальных сенсорных систем, понижается.
У человека в процессе эволюции снизилась чувствительность многих сенсорных систем (зрительной, слуховой, обонятельной), но не вкусовой. Вероятно, последняя не только не атрофировалась, но и получила развитие в связи с расширением вкусовых ощущений. Кулинарная обработка природных пищевых средств, появление в питании человека искусственных продуктов, возможно, привели к обострению и расширению вкусовой чувствительности современного человека.
Таким образом, вкусовая рецепция базируется на хорошо развитой морфофункциональной основе и обеспечивает животным разного уровня эволюционного развития сложные стереотипы пищевого поведения. Кроме того, вкусовая сенсорика вносит свою корректировку в формирование ориентировочного, полового и оборонительного поведения.
Обоняние у млекопитающих развито гораздо лучше, чем у других наземных позвоночных, что указывает на его огромное значение. О последнем говорит хотя бы то, что наиболее обширное семейство генов в их геноме образуют как раз гены, кодирующие белки обонятельных рецепторов (установлено, что у человека и мыши они составляют 3-5 % от общего количества генов). Более того, высшие нервные центры сформировались у млекопитающих именно в тех областях мозга, которые первоначально были связаны с обонянием, что является свидетельством его первостепенной исторической роли. Разрешающая способность обонятельной системы многих рыб значительно выше — они воспринимают запаховые стимулы в концентрациях, меньших на четыре порядка. Несмотря на это, собака способна запомнить индивидуальный запах любого из 7 млрд. человек, живущих сейчас на Земле, и отличить его в условиях чрезвычайного зашумления среды.

Нос собаки (Canis lupus familiaris).
Для улавливания различных запахов немаловажен мясистый наружный нос, особенно развитый у насекомоядных, хищных, копытных и хоботных. Носовая полость черепа, сообщающаяся с ротоглочной полостью хоанами, равным образом обслуживает и дыхание, и обоняние. Обонятельные рецепторы располагаются в верхнезадней части носовой полости — обонятельной капсуле. Ее прогрессивное развитие, обуславливающее остроту обоняния, выражается в основном в увеличении объема и усложнении вследствие образования системы ответвлений — обонятельных раковин (turbinalia). В зависимости от локализации они делятся на несколько групп — ethmoturbinalia, nasoturbinalia и maxilloturbinalia. Раковины представляют собой тонкие костные лепестки, направленные внутрь полости и выстланные тонкой чувствительной мембраной — обонятельной слизистой. Она образована крупными клетками опорного эпителия, в толще которого скрыты мелкие рецепторные клетки, способные улавливать пахучие вещества (ароматизированные молекулы различной химической природы). Расположенные здесь же слизистые клетки покрывают мембрану слоем слизи, предотвращающим ее от высыхания.
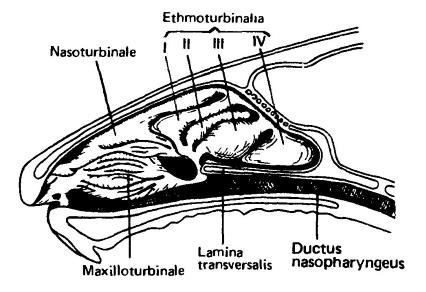
Разрез через носовую область головы морской свинки (Cavia porcellus). Плоскость разреза располагается несколько правее средней линии, чтобы были видны носовые раковин maxilloturbinale, nasoturbinale и ethmoturbinalia. Lamina transversalis — остаток первичного неба, отделяющий сенсорную область носовой полости от носоглоточного хода.
Обонятельные рецепторные клетки представляют собой биполярные нейроны с 1 неветвящимся дендритом. Он проходит между базальными клетками и оканчивается небольшим вздутием — обонятельной булавой. Из нее выходит щетка коротких ресничек, которые представляют собой сенсорную поверхность обонятельной клетки. Реснички, как правило, погружены в слой слизи, покрывющей эпителий, и формируют с ним плотный матрикс. В совокупности реснички образуют как бы ворсистый ковер, густота которого зависит от вида животного. За счет подобных структурных особенностей у обонятельного нейрона многократно увеличивается реакционная способность, т. е. возможность плотного контактирования с одорантом. Аксоны обонятельных нейронов объединяются в пучки — обонятельные нити, которые выполняют передачу сигналов в центральную нервную систему. Обонятельные нейроны способны к замещению путем деления базальных клеток. Кроме обонятельных клеток, связанных с обонятельным нервом, в слизистой оболочке носа имеются также свободные окончания тройничного нерва; они способны реагировать на некоторые агрессивные запахи, например, кислотные или аммиачные испарения.
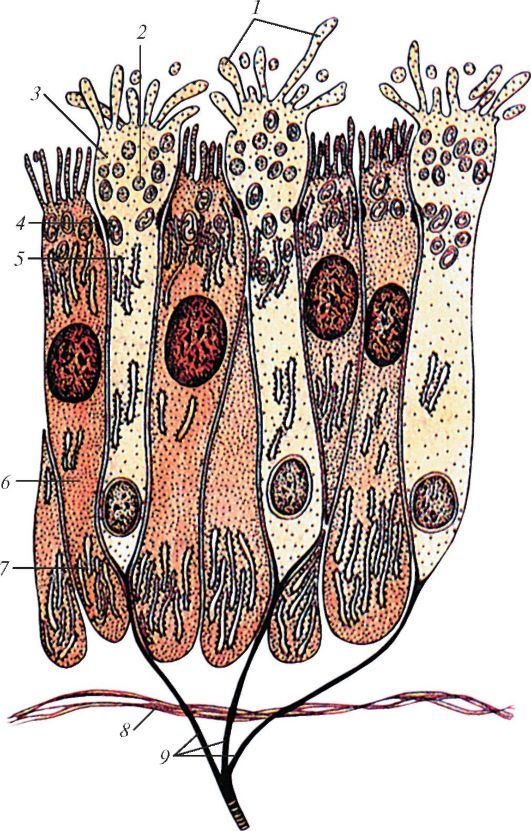
Схема строения обонятельного эпителия млекопитающих: 1 — микроворсинки; 2 — пузырьки; 3 — обонятельная булава; 4 — замыкательная пластинка; 5 — тело обонятельной нейросенсорной клетки; 6 — поддерживающая клетка; 7 — эндоплазматическая сеть; 8 — базальная мембрана; 9 — аксоны обонятельных нейросенсорных клеток, образующие обонятельные нити.
При дыхании воздух поступает в общий, нижний и средний носовые ходы, и лишь малая его часть уходит в носовой лабиринт и околоносовые полости. Основной поток через хоаны направляется к носоглотке, а далее в трахею. Если в составе вдыхаемого воздуха млекопитающее почувствовало необычный запах, оно приостанавливает ритмичное дыхание и начинает принюхиваться, совершая частые и поверхностные вдохи и выдохи. При этом воздушный поток радикально перераспределяется по носовым ходам. Большая его часть теперь засасывается в верхний носовой проход и прокачивается через обонятельные раковины, не достигая легких. В случае, когда собака принюхивается, скорость движения воздуха по носовым ходам достигает 1 л/с. Это в 10 раз быстрее, чем при спокойном дыхании животного. Система извитых ходов, каналов и складок вынуждает воздух двигаться не прямолинейно, а с завихрениями. Следовательно, пахучие вещества неизбежно вступают в контакт со слизистой обонятельного лабиринта.
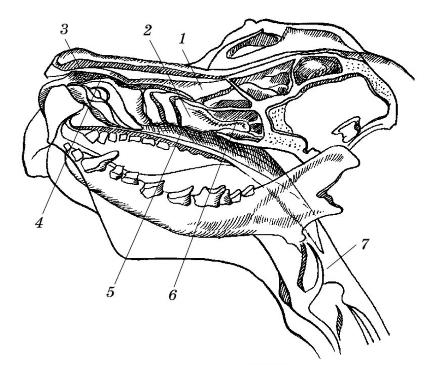
Структуры носовой полости, в которые закачивается воздух в процессе принюхивания собаки (Canis lupus familiaris): 1 — верхняя носовая раковина; 2 — нижняя носовая раковина; 3 — верхний носовой проход; 4 — средний носовой проход; 5 — нижний носовой проход; 6 — пазухи решетчатой кости; 7 — глотка.
Итак, вдыхаемые вместе с воздухом микродозы химических веществ попадают на обонятельную мембрану и растворяются в слизи. Рецепторные белки располагаются на мембране ресничек. Как отмечалось выше, они кодируются весьма обширным семейством генов, поэтому возможно, что одна обонятельная клетка экспрессирует лишь один из этих генов. Однако не все эти гены могут экспрессироваться; так, у человека их задействовано около 40 %. Обонятельные клетки определенных типов специфичны к конкретным соединениям, поскольку распознают особые структурные мотивы в них. Ранее полагали, что обонятельная чувствительность основывается на ощущении различных комбинаций всего нескольких основных запахов, однако сегодня установлено, что млекопитающими обнаруживается и воспринимается свыше 10 тыс. различных запахов. Проведение обонятельного стимула осуществляется следующим образом. Пахучее вещество связывается с рецептором в мембране обонятельной клетки. Начинается сложная химическая реакция, результатом которой становится запуск электрического потенциала действия, передающегося затем на афферентный нейрон.
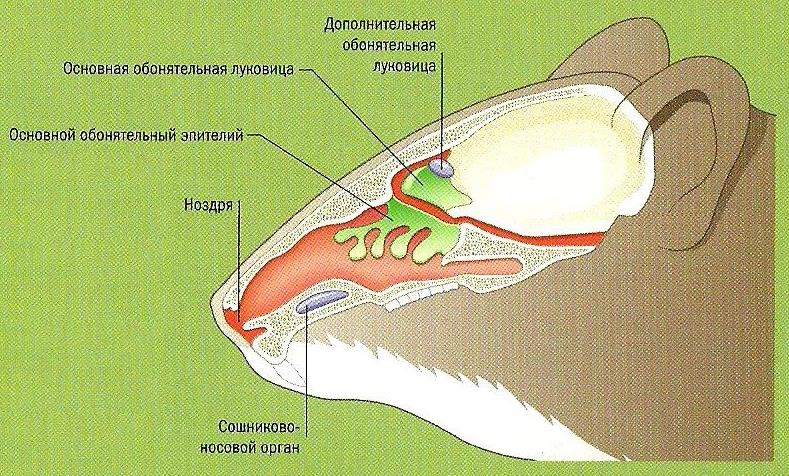
Схема обонятельной системы домовой мыши (Mus musculus).
Из обонятельной капсулы обонятельные нити тянутся к обонятельным луковицам (bulbi olfactorii), расположенным впереди больших полушарий переднего мозга. При этом луковицы развиты тем лучше, чем большую роль играет обонятельная сенсорная система в жизни животного. Спереди и с боков луковицы одеты ажурной костной оболочкой — продырявленной пластинкой (lamina cribrosa) срединной обонятельной кости (mesethmoideum). Из луковиц обонятельные сигналы передаются в обонятельный центр больших полушарий, где происходит окончательное формирование запахового образа, а также в лимбическую систему, которая отвечает за эмоционально-мотивационное состояние животных. Таким образом, определение конкретного запаха является результатом совместной работы рецепторов и мозга.
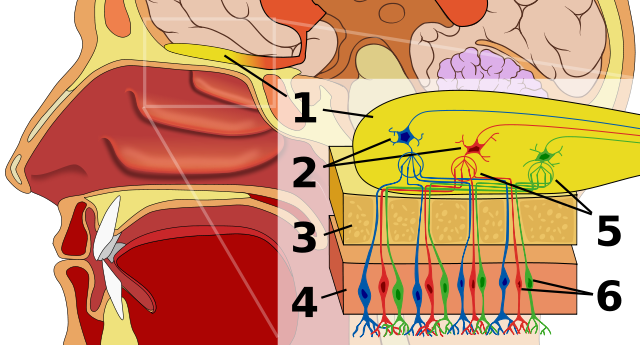
Схема начального отдела нервных обонятельных путей: 1 — обонятельная луковица; 2 — митральные клетки; 3 — кость; 4 — носовой эпителий; 5 — обонятельный клубочек; 6 — обонятельные рецепторы.
По степени развития обонятельной функции млекопитающих разделяют на макросматов с исключительно острым обонянием (сумчатые, ксенартры, насекомоядные, грызуны, большинство хищных и копытных) и микросматов с относительно слабо развитым обонянием (рукокрылые, приматы, ластоногие, сирены). Различие между данными группами хорошо видно при сопоставлении степени развития обоняния у человека и собаки. Если у человека имеется около 5 млн. обонятельных рецепторов на слизистой площадью в 2-3 кв. см, то у собаки средних размеров их больше 200 млн. на площади 85 кв. см. Экспериментальные исследования показали, что порог восприятия запаховых стимулов собакой оценивается в 106-108 раз ниже по сравнению с чувствительностью человека, а острота обоняния на три порядка выше по сравнению с человеческой. Тонкость обоняния коррелирует с числом генов, кодирующих различные типы функциональных белков обонятельных рецепторов. У макросматов этих генов обычно более 1000, у многих приматов — около 500, у человека — 387, у утконоса (Ornithorhynchus anatinus) — всего 262.
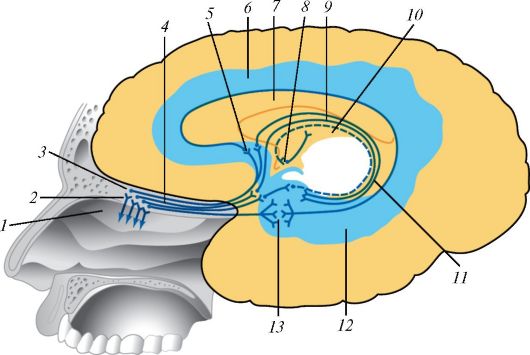
Проводящий путь органа обоняния: 1 — верхняя носовая раковина; 2 — обонятельные нервы; 3 — обонятельная луковица; 4 — обонятельный тракт; 5 — подмозолистое поле; 6 — поясная извилина; 7 — мозолистое тело; 8 — сосцевидное тело; 9 — свод; 10 — задний таламус; 11 — зубчатая извилина; 12 — извилина гиппокампа; 13 — крючок.
Об остроте обоняния макросматов свидетельствует следующий пример. Экспериментально доказано, что собака способна распознать запах раствора, в котором на каждый миллиард частей смеси приходится всего 1-2 части пахучего вещества. Поэтому она легко находит дичь по следу, оставленному несколько часов назад на чистом снегу, определяя при этом не только присутствие объекта, но и направление его движения. В следовой работе собака фиксирует малейшие нюансы интенсивности запаха, которая возрастает в сторону удаляющегося объекта. Наилучшие условия для следовой работы складываются тогда, когда температура земли несколько выше температуры воздуха. При этом летучие вещества, входящие в состав запаха следа, испаряются и держатся в прилегающих к земле слоях воздуха. Собачье обоняние издавна используется для поиска людей, веществ и предметов. По запаху выдыхаемого человеком воздуха собака даже способна почувствовать наличие у него рака груди или легких. Благодаря этому болезнь может быть диагностирована раньше, что способствует более эффективному ее лечению. Многие другие виды тоже способны чувствовать запахи за несколько сот метров и обнаруживать пищевые объекты, находящиеся под землей. Хорошо известна практика поиска подземных трюфелей при помощи специально обученных свиней, которые способны учуять их на расстоянии до 20 м. Уникальной для млекопитающих является способность чувствовать запах воды (отсутствует у видов, проводящих всю жизнь в воде, и видов, получающих всю необходимую воду с животной или растительной пищей).
У микросматов доминирующим чувством является зрение или слух. Обоняние летучих мышей не слишком развито, но обладает стереоскопичными свойствами. Нюховые рецепторы, расположенные у них в каждой из ноздрей, передают в головной мозг обонятельную информацию независимо друг от друга. Мозг перерабатывает эти сигналы и создает стереоскопическую «обонятельную картину мира». Это дает летучим мышам огромные преимущества перед их жертвами и другими хищниками. У ластоногих обонятельные центры мозга малы, но запахи в их социальной жизни играют важную роль.
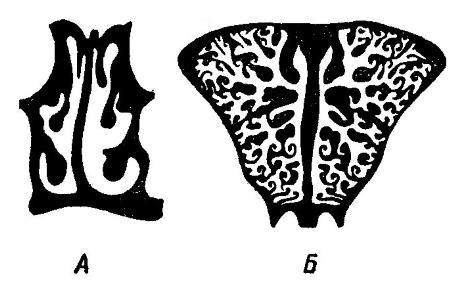
Схема поперечного среза через заднюю часть носовой полости: А — микросмат человек (Homo sapiens); Б — макросмат косуля (Capreolus capreolus).
Чувствительность животных к запаховым раздражителям среды обитания не постоянна. Она зависит от ряда обстоятельств. Так, острота обоняния повышена у голодной собаки и понижена у животных после кормления. В одинаковых условиях суки более чувствительны к запахам по сравнению с кобелями. В то же время обонятельная чувствительность суки в разные периоды полового цикла не одинакова. По мере старения животного обонятельная чувствительность снижается.
У большей части млекопитающих также сохраняется обособленный отдел обонятельной капсулы — вомероназальный, или якобсонов, орган (organum vomeronasale, или organum Jacobsonii), который также имеется у амфибий и многих рептилий. У млекопитающих он замурован в дне носовой полости под обонятельными раковинами и имеет вид парных симметрично расположенных сигарообразных мешков, слепые концы которых доходят до уровня III-IV-го коренного зуба. Мешки построены из хряща, имеют обильное кровоснабжение и богатую иннервацию. Изнутри они выстланы хорошо развитой слизистой оболочкой, нашпигованной чувствительными нервными окончаниями (обонятельными рецепторами), и заполнены слизистой жидкостью. У большинства форм мешки спереди переходят в парные небно-носовые каналы (ductus nasopalatinus), которые связаны с ротовой полостью посредством резцовых отверстий (foramina incisiva), расположенных на переднем конце неба.
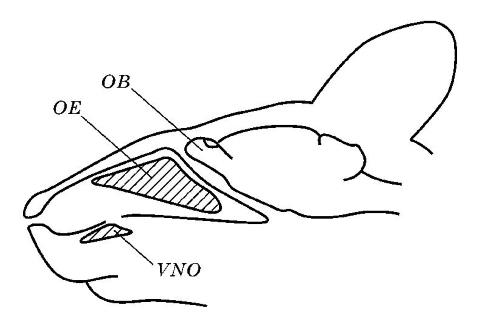
Схематическое расположение вомероназального органа у мыши (Mus musculus): VNO — вомероназальный орган; ОЕ — обонятельные мешки; ОВ — обонятельная луковица.
Вомероназальный орган хорошо развит у однопроходных, сумчатых, ксенартр, насекомоядных, рукокрылых, грызунов и копытных, но редуцирован у сухоносых приматов, части рукокрылых и различных водных зверей. Он реагирует на феромоны и другие летучие ароматные вещества, в большинстве своем не ощущаемые как запах или слабо воспринимаемые обонянием. Функции и механизмы работы этого органа окончательно не установлены. Установлено, например, что запаховые сигналы в моче самок мышей, содержащихся большими группами и испытывающих поэтому сильный стресс, подавляют половое развитие молодых особей того же пола, а запаховые сигналы самцов, наоборот, ускоряют созревание самок. Другие проведенные на грызунах исследования показывают, что вомероназальный орган играет важную роль не только в половом поведении. Воспринимаемые им стимулы также необходимы для формирования и поддержания социальных связей между сородичами. Определенную роль восприятие сигналов вомероназальным органом играет и в агрессивном, и в родительском поведении.
У некоторых млекопитающих присутствует характерное движение губ — флемен, которое связано с захватом летучих ароматных веществ в зону вомероназального органа. Почуяв запах самки, самцы копытных вытягивают вперед голову и шею, напрягают мышцы ноздрей и широко раздвигают губы, оголяя верхние резцы и десну. При этом животные совершают частые вертикальные движения головой и сильно, но поверхностно втягивают ртом воздух. Самцы хищных поднимают и сморщивают верхнюю губу, совершая языком движения, напоминающие процесс лакания жидкости. Благодаря этим действиям источаемые самкой феромоны доставляются в вомероназальный орган и после их анализа позволяют определить, готова ли она к спариванию, или нет. Перед поединком жеребцы откладывают катышки навоза, приглашают друг друга к их обнюхиванию и «флемуют», а кобыла после родов обнюхивает плодные оболочки и жидкость на месте родов, а затем, опять же, «флемует».

Флемен у зебры.
У грызунов мешки вомероназального органа соединены проходом с основной носовой полостью. Одоранты, растворенные в слизи обонятельного эпителия, через этот проход накачиваются в полости вомероназального органа за счет изменения кровотока в последнем. Рецепция феромонов в чувствительном эпителии вомероназального органа осуществляется тем же самым образом, что и в обонятельном эпителии. Рецепторные биполярные нейроны направляют свои аксоны в обонятельные луковицы. Аксоны от рецепторов обонятельного эпителия и эпителия вомероназального органа идут параллельно. Но аксоны релейных нейронов обонятельных луковиц далее расходятся. Если пути запаховой рецепции направляются в новую кору больших полушарий, включая фронтальную область, то афферентный поток вомероназального органа уходит в лимбическую систему и структуры гипоталамуса, связанные с размножением. Функциональная активность вомероназального органа возникает на фоне высокого уровня половых гормонов, а следствием его активизации является формирование соответствующих поведенческих стереотипов.
У китообразных наличие обоняния и вкуса долгое время полностью отрицалось, а самих их называли аносматами. Однако выяснилось, что дельфины имеют пахучие железы, открывающиеся близ анального отверстия. Эти животные способны определять по следам их секрета направление прошедшего стада, а также воспринимают запах крови как сигнал опасности. У корня языка зубатых китов расположены продолговатые ямки, напоминающие вкусовые луковицы других млекопитающих, а в ротовой полости усатых китов имеются парные углубления на конце верхней челюсти, гомологичные вомероназальному органу. Можно предположить, что с помощью этих приспособлений киты распознают запахи и ориентируются, различая течения с разным химизмом. Мозг китообразных сохраняет в коре полушарий структуры, связанные с анализом химических сигналов, вместе с тем характеризуясь редукцией обонятельных долей. Показателен также самый высокий среди млекопитающих процент нефункциональных обонятельных генов. Таким образом, хеморецепция развита у китообразных весьма умеренно и ее разрешающая способность относительно невелика.

Флемен у тигра (Panthera tigris).
Таким образом, различение запахов используется млекопитающими для ориентации в пространстве, при поисках пищи и воды, во внутривидовых и межвидовых коммуникациях. При этом ряд приятных запахов может запускать секрецию слюны и желудочного сока, а неприятные предупреждают о потенциальном вреде (например, запах испорченной пищи). Большая разрешающая способность хеморецепторов позволяет различать отдельные специфические вещества или их сочетания, характерные для вида, группы особей и даже отдельных индивидов. В этой связи у многих представителей класса развились специфические адаптации к формированию индивидуального запаха, его фиксации и запаховому мечению территории. В то же время, некоторые повадки хищников направлены на маскировку собственного запаха, способного предупредить потенциальную жертву о приближении охотника.
вперед
в оглавление