в оглавление
назад
Равновесие и слух
Звук возникает вследствие ничтожных изменений давления, вызываемых источником колебаний. С точки зрения физики звук характеризуется частотой колебаний волны сжатия (измеряется в герцах) и силой сжатия среды (амплитуда колебаний). Субъективно эти характеристики звука оцениваются в терминах «высокие - низкие» и «тихие - громкие» звуки. Дополнительно частота колебаний оценивается обертонами. Физические свойства почвы, воздушной и водной среды различны, поэтому характеристики и закономерности распространения звуковой волны в них неодинаковы. В воздушной среде звук распространяется со скоростью около 340 м в секунду, причем в горячем воздухе — быстрее, чем в холодном. В воде скорость звука в 4,5 раза выше, чем в воздухе.
Наличие двух ушей позволяет животному определять нахождение источника звука в пространстве. Если морда не направлена прямо на источник звука, возикает разница во времени между восприятием звука ухом, расположенным ближе к источнику, и тем, что находится от источника дальше. С учетом этой разницы в доли секунды, головной мозг определяет направление звука. Звуковые рецепторы в принципе представляют собой механорецепторы с быстрым восстановлением чувствительности, что делает их восприимчивыми к колебаниям давления среды. Слух обслуживает такие важные в жизни животных явления, как поиск пищи, распознавание опасностей, опознавание особей своего и чужих видов.
Равновесно-слуховой аппарат млекопитающих организован иначе, чем у рептилий и птиц. Он включает наружное, среднее и внутреннее ухо, слуховой нерв, проводящие тракты и анализирующие центры в головном мозге. Наружное ухо, зачаточное у рептилий и слабо развитое у птиц, представлено у млекопитающих глубоким наружным слуховым проходом и обрамляющим его вход ушной раковиной. Стенки прохода образованы краями барабанной кости, изнутри он замыкается барабанной перепонкой. Обычно она представляет собой тонкую мембрану, способную колебаться под действием воздушных волн. Мембрана образована листком соединительной ткани, выстланной снаружи и изнутри слоистым эпителием. У китообразных узкий слуховой проход кнаружи заканчивается слепо, а барабанная перепонка неподатлива; хорошо распространяющиеся в воде звуковые колебания передаются сразу на слуховые косточки этих животных.
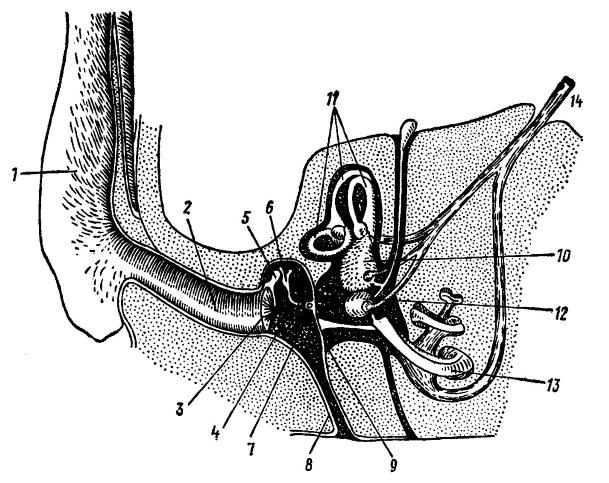
Схема органа слуха млекопитающих на примере человека: 1 — наружное ухо; 2 — наружный слуховой проход; 3 — барабанная перепонка; 4 — полость среднего уха; 5 — молоточек; 6 — наковальня; 7 — стремечко; 8 — евстахиева труба; 9 — круглое окно; 10 — овальный мешочек; 11 — полукружные каналы; 12 — круглый мешочек; 13 — улитка; 14 — слуховой нерв.
Ушная раковина представляет собой кожную ушную складку, армированную хрящевыми элементами. Раковина служит рефлектором, отбирающим и усиливающим важные звуки, и частотным фильтром, ослабляющим посторонние шумы. У большинства млекопитающих она приводится в движение специальными мышцами (в частности, у кошки (Felis silvestris) их 20), что способствует сравнительно высокой точности (до 5-8 °) локализации источников звуков. У многих видов ушные раковины способны поворачиваться и наклоняться независимо друг от друга.
Форма и величина ушей сильно варьирует. Очень крупных размеров достигают ушные раковины у летучих мышей (Microchiroptera), зайцев (Leporidae), тушканчиков (Dipodidae) и других зверей, обладающих острым слухом, у которых они действуют как резонаторы. В то же время, размеры раковин могут быть связаны с терморегуляцией — большие уши лучше рассеивают тепло. У ряда полуводных зверей (бобров (Castor), калана (Enhydra lutris), ушастых тюленей (Otariidae)) ушные раковины при нырянии способны замыкаться. Водными формами (китообразными (Cetacea), сиренами (Sirenia), настоящими тюленями (Phocidae)) и землероями (кротами (Talpidae), слепышами (Spalacidae)) они вторично утрачены.

Ушные раковины фенека (Vulpes zerda) достигают 15 см в длину при размере зверя 60 см от носа до хвоста. Уши обладают огромной поверхностью, достаточно подвижны и обеспечивают фенеку острый слух. Он улавливает малейшие шорохи потенциальной добычи — насекомых, мелких рептилий и грызунов. Охотится фенек ночью, а в сильную дневную жару прячется в подземной норе. Кроме того, ушные раковины пронизаны сетью капилляров, расположенных близко к поверхности кожи. Вероятно, они участвуют в теплообмене — через капилляры хорошо происходит отдача тепла.
В заполненном воздухом среднем ухе (барабанной полости) расположены слуховые косточки (ossicula auditis). Наружная косточка — молоточек (malleus) прикреплена своим длинным отростком (рукояткой) к барабанной перепонке, а другим концом сочленяется со второй косточкой — наковальней (incus), которая своим коротким отростком прикреплена к стенке барабанной полости, а длинным, служащим рычагом отростком соединяется с третьей косточкой — стремечком (stapes), закрывающим своим основанием овальное окно (fenestra ovalis). Так возникает трехчленная цепочка, передающая звуковые колебания от большей по величине барабанной перепонки, отделяющей среднее ухо от наружного, на меньшую по площади мембрану овального окна внутреннего уха. При прохождении через слуховые косточки амплитуда колебаний уменьшается, а сила увеличивается. В итоге звуковое давление на овальном окне значительно превосходит таковое на барабанной перепонке (у человека — приблизительно в 22 раза), что улучшает обнаружение слабых звуков. Слуховые косточки человека усиливают слышимый звук в 18 раз, но это далеко не предел. Например, у кенгурового прыгуна (Dipodomys sp.) звук усиливается в 100 раз, что позволяет улавливать шорох чешуек охотящейся на него змеи, а медоед (Mellivora capensis) способен расслышать движения личинок навозных жуков в коконах под землей.
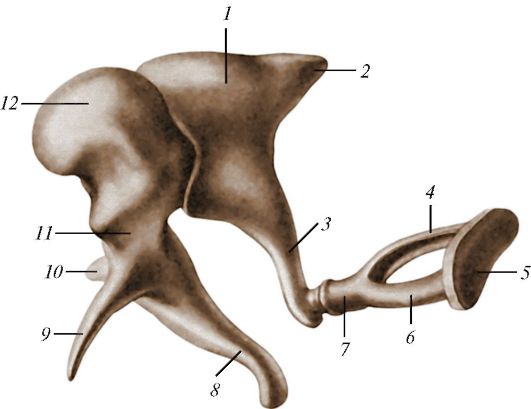
Слуховые косточки млекопитающих на примере человека: 1 — наковальня; 2 — короткая ножка наковальни; 3 — длинная ножка наковальни; 4 — задняя ножка стремечка; 5 — основание стремечка; 6 — передняя ножка стремечка; 7 — головка стремечка; 8 — рукоятка молоточка; 9 — передний отросток молоточка; 10 — латеральный отросток молоточка; 11 — шейка молоточка; 12 — головка молоточка.
К 2 слуховым косточкам прикрепляются маленькие мышцы, которые осуществляют регулировку звукопроводящей системы. Примечательно, что мускул, связанный с молоточком, происходит из мускулатуры челюстной дуги и иннервируется тройничным нервом, тогда как мускул, связанный со стременем, происходит из мускулатуры гиоидной дуги и иннервируется лицевым нервом. Судя по всему, тройственность слуховых косточек представляет собой не средство повышения функциональной эффективности, а случайный по отношению к этим свойствам результат уникального пройденного млекопитающими эволюционного пути.
У многих млекопитающих полость среднего уха значительно увеличивается вследствие связи с вмещающим ее костным слуховым барабаном (bulla tympani). Слуховые барабаны образуют ряд камер-резонаторов, усиливающих биологически важные звуки; у некоторых видов, особенно норников, они порой заполняются губчатой костной массой, гасящей паразитические шумы. Близ основания барабанной перепонки от нижней полости среднего уха отходит канал евстахиевой, или слуховой, трубы (tuba Eustachii, или tuba auditiva), сообщающей среднее ухо с полостью глотки. Такое соединение обеспечивает выравнивание давления воздуха по обе стороны барабанной перепонки, чему способствует жевание, глотание и зевота.

Череп льва (Panthera leo), вид снизу. В задней части хорошо видны слуховые барабаны.
Замурованное в каменистой части височной кости внутреннее ухо состоит из слухового и вестибулярного отделов. Слуховой отдел имеет следующее строение. Снаружи он ограничен костным лабиринтом каменистой кости, подобно футляру вмещающим в себе перепончатый лабиринт (labyrinthus membranaceus). На обращенной к среднему уху стенке костного лабиринта имеются 2 отверстия, затянутые перепонками. Верхнее из них, овальное окно (fenestra ovalis), закрыто основанием стремечка; нижнее, круглое окно (fenestra rotunda), затянуто только перепонкой. Основным элементом перепончатого лабиринта является улитка (cochlea) — спирально изогнутая трубка, выстланная эпителием и заполненная жидкостью. У архозавров и птиц улитка лишь намечается, у однопроходных она также не делает оборотов и представляет собой бутылкообразное выпячивание — т. н. выступ (lagena). У сумчатых и плацентарных улитка вполне развита. У китообразных она делает 1,5 спиральных оборота, у копытных — 2-3,5, у хищных — 3, у грызунов — от 4 до 5. Выступ в таких случаях превращен в длинную слепо заканчивающуюся трубку — улитковый проток. Улитковый проток разделен 2 широкими каналами — верхней вестибулярной лестницей и нижней барабанной лестницей, между которыми залегает узкий канал — средняя лестница.
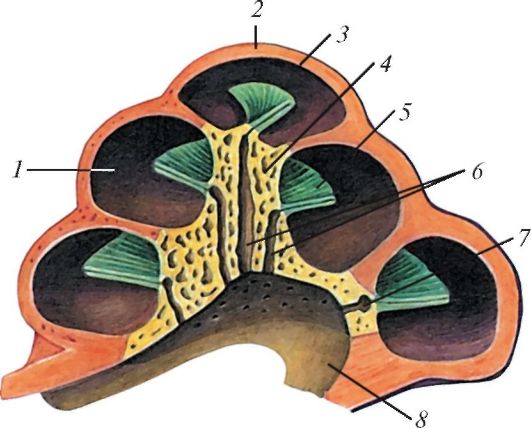
Костная улитка человека на предельном ее разрезе: 1 — спиральный канал улитки; 2 — купол улитки; 3 — геликотрема; 4 — стержень улитки; 5 — костная спиральная пластинка; 6 — продольные каналы стержня; 7 — спиральный канал стержня; 8 — основание улитки.
Вдоль улиткового протока в полости средней лестницы расположен кортиев орган (organum spirale), представляющий собой сложный набор сенсорных и опорных клеток. Данное образование присуще только млекопитающим, у прочих амниот его аналогом служит базилярный сосочек. Снизу кортиева органа располагается базилярная мембрана, пересекаемая множеством тончайших фибрилл — слуховых струн (у человека их количество может достигать 20 тыс.). Длина струн постепенно уменьшается соответственно сужению диаметра канала, т. е. по направлению к вершинной части улитки. Сверху помещается перепончатая покровная мембрана (membrana tectoria), соответствующая купуле. В толще кортиева органа проходит узкий треугольный туннель, ограниченный с обеих сторон столбчатыми клетками. По бокам от этой структуры по всей длине улитки располагаются рецепторные волосковые клетки. Их основания покоятся на базилярной мембране, а кончики связаны с покровной мембраной. Волосковые клетки обычно представлены единственным рядом ближе к оси улитки и тремя или четырьмя рядами по ее периферии. Ближе к краям базилярной мембраны по обе стороны расположены ряды опорных клеток.
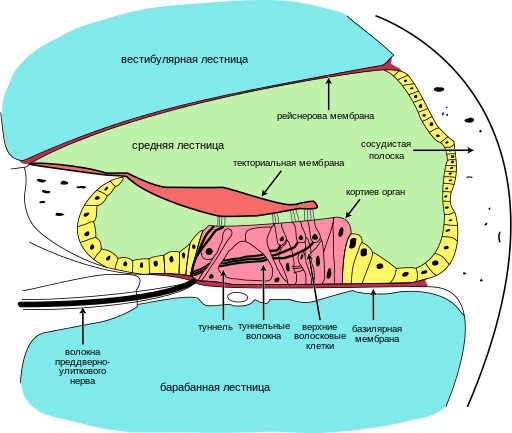
Схема строения улиткового протока внутреннего уха млекопитающих.
Волосковые клетки представляют собой механорецепторы, снабженные пучком из нескольких десятков «волосков» — стереоцилий, среди которых имеется расположенный обособленно самый длинный «волосок» — киноцилия. Внешний механический стимул вызыват отклонение (изгиб) пучка стереоцилий. Если стереоцилии отклоняются в сторону киноцилии, волосковая клетка деполяризуется. В результате этого внутрь клетки поступает кальций, и она высвобождает в синаптическую щель нейромедиатор глутамат. Последнее в свою очередь увеличивает частоту генерации электрического потенциала действия афферентного нейрона, передающего сигнал в центральную нервную систему. При отклонении стереоцилий от киноцилии имеет место обратный эффект, и волосковая клетка гиперполяризуется.
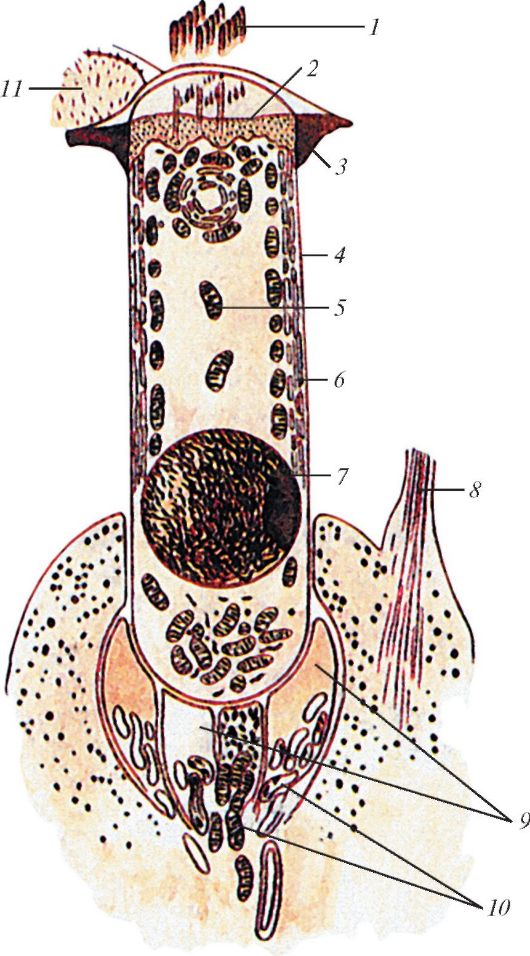
Схема строения волосковой клетки кортиева органа: 1 — слуховые волоски; 2 — кутикула; 3 — сетчатая мембрана; 4 — цитолемма клетки; 5 — митохондрия; 6 — эндоплазматическая сеть; 7 — ядро; 8 — фаланговая пластинка наружной поддерживающей клетки; 9 — нервные окончания; 10 — митохондрии в нервном окончании; 11 — микроворсинки на опорных клетках.
Вибрации от стремечка, расположенного напротив овального окна, передаются перилимфе — жидкости, заполняющей вестибулярную и барабанную лестницы. При прогибании мембраны овального окна внутрь улитки волна перелимфы доходит до конца вестибулярной лестницы и заворачивает в барабанную лестницу, откуда движется к основанию улитки и достигает круглого окна. От этого мембрана круглого окна выгибается наружу, и давление в улитке снижается. За полный цикл жидкость движется сначала в одну, а затем в другую сторону. Эти движения в улитке происходят с частотой колебаний наружного воздуха. Их энергия заставляет вибрировать туго натянутую базилярную мембрану. Отклоняясь вверх и вниз, она вызывает сдвиг покровной мембраны и последующее смещение волосков чувствительных клеток, погруженных во внутреннюю жидкость средней лестницы — эндолимфу. Подобная стимуляция и порождает слуховые ощущения. С возрастом волосковые клетки млекопитающих атрофируются, что приводит к ухудшению слуха и даже глухоте. Единственным исключением из этого правила являются летучие мыши: вместо старых волосков у них образуются новые.
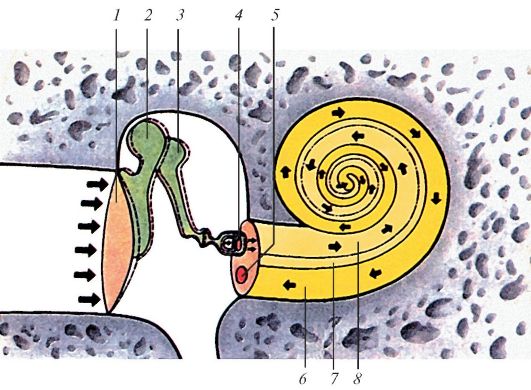
Распространение звуковой волны (показано стрелками) в наружном, среднем и внутреннем ухе млекопитающих: 1 — барабанная перепонка; 2 — молоточек; 3 — наковальня; 4 — стремечкп; 5 — круглое окно; 6 — барабанная лестница; 7 — улитковый проток; 8 — лестница преддверия.
Функциональной причиной удлинения улитки, а вместе с ней и кортиева органа, явилась потребность млекопитающих в различении звуков разной частоты. Различия в ширине базилярной мембраны, которая сужается от одного конца кортиева органа к другому, приводят к тому, что разные ее участки способны резонировать с волнами различной длины, чем обеспечивается тональное разрешение. Основание улитки, где базилярная мембрана шире и сильнее натянута, наиболее чувствительно к высоким частотам, а вершина, где узкая мембрана натянута слабо, улавливает низкие частоты. Улитка большинства млекопитающих обладает прекрасной разрешающей способностью, при этом диапазон слышимых колебаний зависит от конкретного вида.
Импульсы, воспринятые волосковыми клетками, передаются на лежащие у их основания дендриты сенсорных нейронов. Их тела залегают в спиральном ганглии, расположенного в толще спиральной костной пластинки, а аксоны образуют подходящий к внутреннему уху слуховой нерв. В этом нерве происходит первичный анализ и кодирование звуковых сигналов. Затем слуховые импульсы направляются в верхнюю оливу продолговатого мозга, далее попадают в задние бугры четверохолмия, потом проходят в медиальное коленчатое тело таламуса и, наконец, достигают слуховой коры, расположенной в височной доле больших полушарий. В коре нейроны располагаются тонотопически в соответствии с частотами слухового диапазона. У человека среди них имеются нейроны, избирательно реагирующие на звуки различной продолжительности, повторяющиеся звуки, шумы с широким частотным диапазоном. В результате происходит объединение информации о высоте, интенсивности тона и временных интервалах между отдельными звуками.
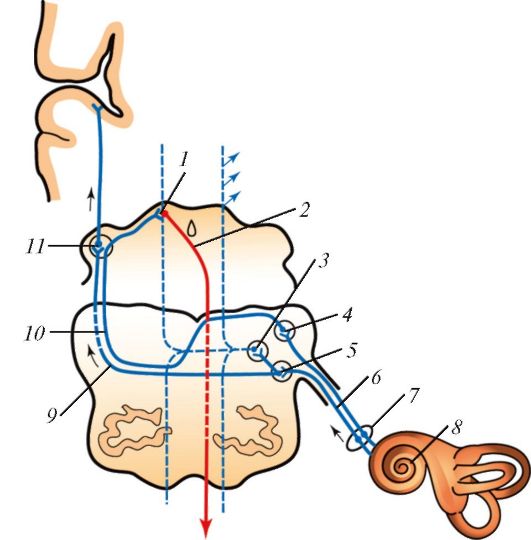
Проводящий путь слухового анализатора млекопитающих: 1 — нижний холмик среднего мозга; 2 — покрышечно-спинномозговой путь; 3 — ядро трапециевидного тела; 4 — заднее улитковое ядро; 5 — переднее улитковое ядро; 6 — улитковая часть преддверно-улиткового нерва; 7 — спиральный узел; 8 — улитка; 9 — трапециевидное тело; 10 — латеральная петля; 11 — медиальное коленчатое тело. Стрелками показан путь распространения.
Частотный диапазон слуха млекопитающих чрезвычайно высок — он лежит в примерных границах от 10 Гц до 300 кГц и охватывает 17 октав, причем слуховая система обладает исключительно ровной чувствительностью в пределах всей воспринимаемой звуковой полосы. Однако как диапазон воспринимаемых звуков, так и границы оптимальной звуковой чувствительности определяются экологическими факторами и акустическими свойствами среды обитания конкретного вида. Так, человек может слышать звуки в пределах от 20 Гц (в исключительных случаях — от 16 Гц) до 20 кГц, тогда как многие млекопитающие способны издавать и воспринимать инфра- и ультразвуки — неслышимые человеком звуковые колебания более низкой и высокой частоты. Например, инфразвуки используют в коммуникации усатые киты (Mysticeti), слоны (Elephantidеa), носороги (Rhinocerotidae) и бегемоты (Hippopotamidae), а ультразвуки воспринимают лошади (Equidae), кошки (Felidae), собаки (Canidae) и некоторые мышевидные грызуны. Долгопяты (Tarsiidae) — единственные известные приматы, общающиеся «на чистом ультразвуке». Они способны улавливать звуки частотой до 90 кГц и кричать на частотах около 70 кГц.
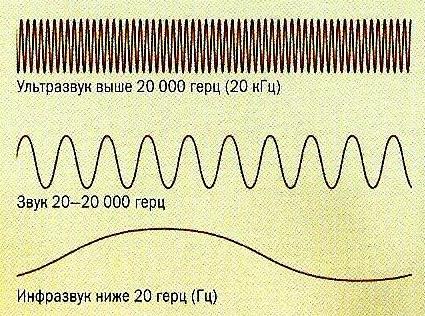
Типы звуковых колебаний.
Тонкость слуха различна в разных отрядах, но обычно весьма значительна. Частотный слуховой диапазон у человека не превышает 8,5 октав, а у кошки (Felis silvestris catus) составляет 10 октав и не менее 10 обертонов в пределах одной октавы. Возню мыши в норе кошка может расслышать с расстояния в 20 м. Готовность слухового анализатора к восприятию звуков постоянна. Даже во время сна кошка не прекращает акустический мониторинг окружающей среды. Об этом можно судить по движениям ее ушных раковин во время сна.

Кошка (Felis silvestris) способна услышать возню грызуна на расстоянии в несколько десятков метров.
Развитому слуху зверей соответствует и сложное устройство их голосового аппарата, производящего разнообразные звуки, часто образующие сложные, организованные во времени последовательности. По широте звукового диапазона млекопитающие превосходят птиц, широко используя как высокие, так и низкие частоты. Наиболее обычными издаваемыми зверьми звуками являются рев, стон, мяуканье и шипенье. Большей частью они производятся колебаниями голосовых связок верхней гортани и резонируют с воздухоносными путями. Ультразвуковые сигналы летучих мышей генерируются ротовым аппаратом и носом. У китообразных в образовании звуков участвуют гортань в целом, края черпаловидных хрящей, воздушные мешки надчерепного носового прохода и наружное дыхало. Помимо голоса некоторые млекопитающие издают механические звуки: клацание и скрежетание зубами (хищные, некоторые копытные и грызуны, приматы), стук рогами, удары ногами о грунт (многие норники, копытные), шум от трения игл (дикобразы (Hystricidae)) и т. п. Звуковая сигнализация и слух обслуживают поиски пищи, распознавание опасности, опознавание особей своего и чужих видов, различие индивидов в группе, отношения родителей и детенышей и многие другие жизненные явления.
Частотный слуховой диапазон некоторых млекопитающих
Слон — 1 - 20 000 Гц
Человек — 20 - 20 000 Гц
Собака — 20 - 45 000 Гц
Кошка — 70 - 64 000 Гц
Летучая мышь — 10 - 160 000 Гц
Дельфин — 110 - 270 000 Гц
По диапазону восприятия и участкам оптимальной слуховой чувствительности млекопитающих можно разделить на четыре группы.
I-я группа представлена подземными животными (кроты (Talpidae), слепыши (Spalacidae), гоферы (Geomyidae)), у которых слух в значительной мере заменяет слабое или совсем отсутствующее зрение. Характерной особенностью этой группы является низкочастотная звуковая рецепция. Подземные животные реагируют на звуки в частотном диапазоне до 20 кГц. Зона оптимальной чувствительности этих животных лежит в пределах 300-500 Гц и 3-6 кГц. Верхний предел чувствительности крота составляет 18 кГц. Под землей хорошо распространяются именно низкочастотные звуки. А в условиях подземного хода (земля + воздушные пробки) наилучшее распространение получают звуки с частотой 300-500 Гц. Для коммуникации эти животные используют звуки в частотном диапазоне 6-7 кГц.
II-ю группу составляют животные открытых и закрытых биотопов, например, всевозможные белки (Sciuridae). Сюда же относят и полуводных животных — ондатру (Ondatra zibethicus), водяную полевку (Arvicola terrestris), сирен, некоторых ластоногих (Pinnipedia). Характерной особенностью большинства представителей этой группы является высокая чувствительность к звукам с частотой 5-15 кГц. Верхний предел чувствительности у этих животных уходит в ультразвуковой диапазон — 50-60 кГц. Они не чувствительны к низкочастотным звукам и имеют пониженную чувствительность к частотам до 1 кГц.
III-я группа — ночные животные — имеет высокую чувствительность к частотам, лежащим на границе звука и ультразвука. В целом животные способны реагировать на широкий спектр звуковых частот — 300 Гц - 100 кГц. Однако наилучшая звуковая чувствительность у сонь (Gliridae), полевок (Arvicolinae), тушканчиков (Dipodidae) регистрируется в диапазоне частот 15-50 кГц. Именно этот диапазон звуковых частот оптимален для животных, активность которых приходится на темное время суток. В ночное время естественный шумовой фон лежит в диапазоне до 10 кГц. Использование звуков за пределами естественных шумовых помех (свыше 10 кГц) позволяет ночным животным беспрепятственно осуществлять дистантную коммуникацию в условиях плохой или нулевой видимости.
Наконец, к IV-й группе относят животных из разных экологических групп, которых объединяет наличие ультразвуковой эхолокации. В оптически неблагоприятных условиях (водная среда на больших глубинах, сильная замутненность, темное время суток, пещеры) они осуществляют дистантную коммуникацию, поиск пищи и ориентацию в пространстве генерируя ультразвуковые сигналы и улавливая их отражение. По уловленному эху головной мозг формирует картину окружающей среды. Верхний предел слуховой чувствительности по условно-рефлекторным реакциям определен в 200-300 кГц. Как правило, различные частотные диапазоны используются млекопитающими данной группы для разных целей — эхолокация и поиск добычи происходят на высоких и сверхвысоких частотах, связь и общение с сородичами — на относительно низких.
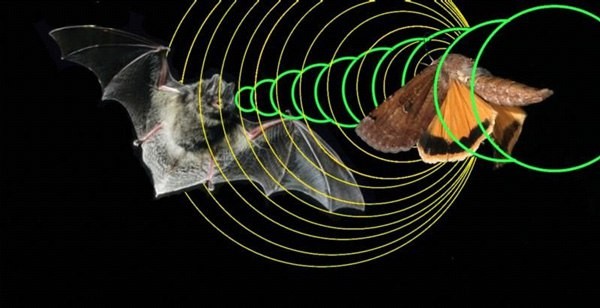
Схема распространения ультразвуковых импульсов, испускаемых летучей мышью, и их отражения от добычи.
Наиболее известны в этом отношении рукокрылые — летучие мыши (Microchiroptera) и некоторые крыланы (Megachiroptera), ведущие ночной и сумеречный образ жизни. В полете эти зверьки испускают залпы ультразвуковых импульсов малой длительности (примерно от 5 до 15 мс) и высокой частоты (от 20 до 160 кГц). Такие короткие импульсы позволяют с высокой точностью определять время образования эха, а значит расстояние до образующего его препятствия или живого объекта. Звуки, производимые другими животными и ветром, обычно бывают низкочастотными, поэтому маловероятно, чтобы на ультразвуковые сигналы рукокрылых накладывались помехи (впрочем, некоторые насекомые специально их создают). Лабораторные опыты показали, что искусственные звуки частотой выше 20 кГц дезориентируют полет летучих мышей.

Частота ультразвуковых импульсов, издаваемых западным нетопырем (Pipistrellus hesperus) на различных стадиях охоты. Длительность сигнала каждой «ноты» составляет около 4-5 мс. (По Уленбрук, 2009.)
Другое преимущество высоких частот состоит в возможности точной фокусировки, что позволяет не только обнаруживать порхающую в воздухе добычу на расстоянии до 5,5 м, но и определять, что она собой представляет, скорость и направление ее полета. В зависимости от размеров жертвы и частоты взмахов ее крыльев, ультразвуковые сигналы по-разному от нее отражаются. Эхо, вернувшееся к летучей мыши от маленького комара-долгоножки, например, будет звучать иначе, чем от большого твердотелого майского жука. Особенности движения летающих насекомых дают охотящейся летучей мыши дополнительную информацию. Если крылья насекомого движутся вверх и вниз, то в точке, где они будут перпендикулярны направлению ультразвукового импульса, издаваемого зверьком, происходит короткое увеличение уровня энергии отраженного звука, известное как «вспышка».
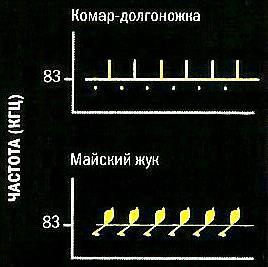
Отражение ультразвуковых импульсов большого подковоноса (Rhinolophus ferrumequinum) от двух разных видов добычи.
Различные виды рукокрылых издают сигналы, зависящие от места их обитания и охоты. Быстро летающие виды, которые охотятся на открытых пространствах, используют сравнительно низкочастотные сигналы. Медленно летающие виды, кормящиеся в зарослях и среди деревьев, издают сигналы большей частоты, что позволяет им различать детали ландшафта. Помимо диапазона частот, эхолокационные сигналы отличаются по продолжительности, повторяемости и форме.
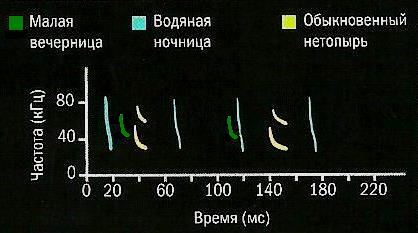
Сонограммы малой вечерницы (Nyctalus leisleri), водяной ночницы (Myotis daubentonii) и обыкновенного нетопыря (Pipistrellus nathusii).
Рукокрылые производят ультразвуковые сигналы особо устроенной гортанью и испускают их через рот, как подбородколисты (Mormoopidae), или из видоизмененных ноздрей, как подковоносы (Rhinolophidae) и различные листоносы (Phyllostomidae). Помимо формирования ультразвукового пучка и придания ему направления, сложные кожные выросты на конце морды зачастую выполняют еще одну важную функцию — воспринимают тепло, излучаемое телом жертвы (некоторые зверьки способны ощущать его на расстоянии до 16 см). Последнее особенно важно для вампиров (Desmodontinae), паразитирующих на крупной теплокровной добыче. У рукокрылых также имеется много приспособлений, позволяющих определять время и локализировать место возникновения эха от их ультразвуковых сигналов. У большинства зверьков, хватающих насекомых на лету, характерны крупные, направленные вперед ушные раковины с выступом — козелком, который помогает улавливать возвращающееся эхо. Специфическая форма ушей повышает их дирекциональную чувствительность. На каждый удар крыльями в полете летучие мыши обычно издают один крик. Эти крики настолько громки, что животные рискуют оглушить сами себя. Чтобы избежать этого, мышцы среднего уха сокращаются, а затем расслабляются, позволяя слышать возвращающееся эхо, прежде чем зверек снова издаст сигнал.

Наросты на морде и сложные уши складчатогубого листоноса (Centurio senex).
При очень коротких испускаемых импульсах, как у кожанов (Vespertilionidae), конец импульса не перекрывается с началом его эха. Поскольку эхо приходит быстрее от близких объектов, импульсы постепенно укорачиваются по мере приближения объекта, и в результате такое перекрывание устраняется. У других летучих мышей издаваемые импульсы и эхо от них перекрываются, поэтому, чтобы улучшить обнаружение эха, им нужны другие средства. Например, большой подковонос (Rhinolophus ferrumequinum) приспосабливает частоту своих сигналов так, чтобы частота возвращающегося эха поддерживалась в узких пределах. Для зверька, летящего к объекту, воспринимаемая частота эха всегда выше, чем частота испускаемого ультразвука. Это объясняется эффектом Доплера, возникающим вследствие относительного движения животного и объекта: чем быстрее они движутся навстречу друг другу, тем выше воспринимаемая частота излучения. Для компенсации такого эффекта летучие мыши меняют частоту своих сигналов так, чтобы воспринимаемая частота была как можно ближе к постоянной. Таким способом мышь может оценить скорость своего полета, а также направление и относительную скорость полета жертвы.
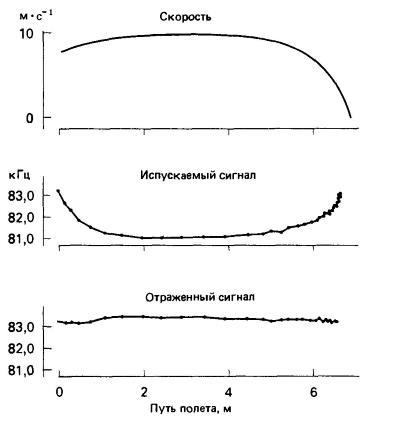
Эхолокация большого подковоноса (Rhinolophus ferrumequinum), приближающегося к неподвижному предмету. По мере приближения к нему скорость полета снижается (верхняя кривая). Летучая мышь приспосабливает частоту своих сигналов (средняя кривая) так, чтобы частота отражаемого ультразвука (нижняя кривая) оставалась постоянной.
Как частичная альтернатива зрению эхолокационные способности рукокрылых производят большое впечатление. Лабораторные исследования показывают, что летучая мышь с размахом крыльев в 40 см может пролететь в полной темноте сквозь сетку с ячейками 14 на 14 см из нейлоновых нитей толщиной всего 80 мкм. Летучие мыши также научаются ловить мелкие частички пищи, брошенные в воздух в полной темноте, и отличать съедобные предметы от несъедобных по небольшим особенностям их формы. Малая бурая ночница (Myotis lucifugus) может ловить на лету очень мелких насекомых, например плодовых мушек и комаров, с «результативностью» 2 насекомых в секунду. Помимо используемых для эхолокации ультразвуковых частот, рукокрылые способны издавать инфразвуки до 12 Гц.

Подковонос (Rhinolophus sp.) пролетает через сетку из нейлоновых нитей толщиной 80 мкм с ячейками 14 на 14 см. Слева — длительность и частота эхолокационных сигналов.
Несмотря на развитую способность к эхолокации, ни один вид рукокрылых не лишен зрения. Небольшие глаза летучих мышей приспособлены для работы в условиях низкой освещенности. В их сетчатке преобладают рецепторы темнового зрения — палочки, спектр чувствительности которых сдвинут в ультрафиолетовый диапазон. Если при тусклом освещении остроту зрения летучих мышей можно считать относительно хорошей, то в ясную ночь их глаза без труда улавливают даже незначительные изменения в яркости света. Таким образом, острота зрения у летучих мышей примерно такая же, как и у других ночных животных, но визуальным ориентированием они пользуются в основном для обнаружения дальних объектов, когда радиуса действия эхолокации не хватает, и при перелете на большие расстояния, например, во время сезонной миграции и смене мест кормежки. У фруктоядных крыланов большие глаза и хорошо развитое дневное зрение. Эхолокацией обладают только некоторые их виды, и она довольно примитивна. Например, египетская летучая собака (Rousettus aegyptiacus) издает парные пронзительные щелчки своим языком.
Китообразные благодаря эхолокации могут быстро передвигаться даже в совершенно непрозрачной воде, не наталкиваясь на препятствия. Используемый ими звуковой диапазон еще шире, чем у рукокрылых — от нескольких герц до 200 кГц и выше. Зубатые киты (Odontoceti) производят высокочастотные последовательности щелчков и свиста. Щелчки состоят из отдельных посылок частотой 20-50 кГц с периодом нарастания порядка 0,1 мс. С помощью отраженных ультразвуков эти животные способны обнаружить предметы величиной с желудь на расстоянии до 15 м и определять местоположение косяков рыб с расстояния до 3 км. Эксперименты с афалинами (Tursiops truncatus) показали, что у них чрезвычайно сложные локаторы, способные различать объекты по размеру, форме и составу. Это позволяет им запоминать звуковые образы своей любимой добычи. При этом для обнаружения добычи афалины издают быстрые серии ультразвуковых щелчков, а для внутривидовой коммуникации воспроизводят свисты, скрипы и ряд других низкочастотных звуков.
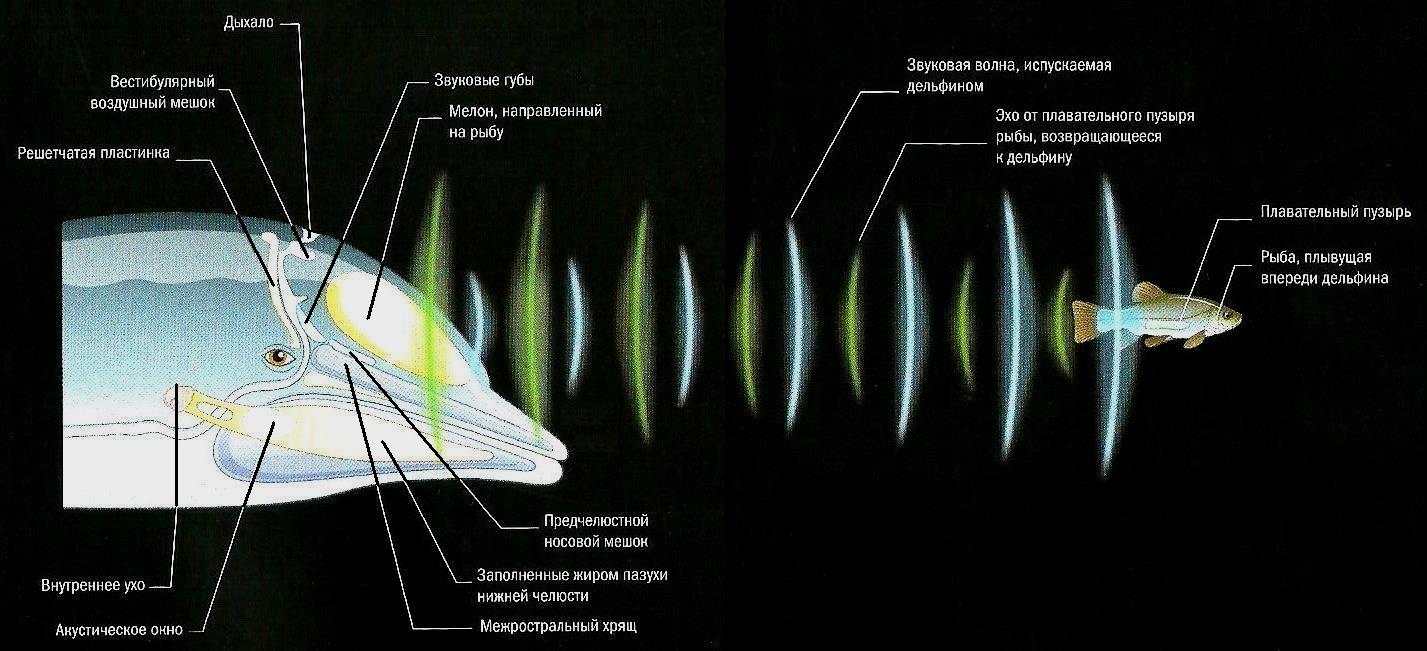
Действие эхолокации у дельфина. С помощью эхолокации можно получить информацию не только об одной рыбе, но и о нескольких рыбах сразу. Звуковые волны хорошо отражаются от их плавательных пузырей.
Дельфины и морские свиньи производят ультразвуковые щелчки следующим образом. Они всплывают на поверхность и набирают через дыхало воздух для дыхания. Накопленный воздух выпускается через 3 пары воздушных мешков. При этом он с силой проходит между т. н. звуковыми губами, расположенными в носовых проходах. Губы открываются и закрываются, заставляя окружающие ткани вибрировать и создавать звуковые волны. Звуки отражаются от костной решетчатой пластинки в передней части черепа и фокусируются с помощью жировой линзы на лбу — мелона. Сформированный таким образом звуковой пучок дельфин направляет на свою добычу. Отраженные от цели звуковые волны (эхо) через заполненные жиром пазухи нижней челюсти проходят к внутреннему уху. Кашалот (Physeter macrocephalus) имеет другое анатомическое приспособление. Его левый носовой проход используется для дыхания, тогда как правый — для производства локационных щелчков. Звуковые волны проходят сквозь заполненный восковидным веществом спермацетовый орган в голове, отражаются от воздушного мешка в задней части черепа и несколькими жировыми линзами фокусируются в звуковой пучок.

Сонограмма кашалота (Physeter macrocephalus).
Усатые киты (Mysticeti) издают звуки низкой частоты (1-2 кГц) большой силы и продолжительности. Зачастую они складываются в повторяющиеся мелодичные последовательности, напоминающие человеческое пение. Полагают, что самые сложные песни используются в брачных играх. Более простые сигналы используются круглый год и, вероятно, служат для повседневного общения и навигации. Физической особенностью сверхнизкочастотных колебаний является их способность распространяться с минимальными изменениями через различные среды. Поэтому усатые киты воспринимают инфразвуковые сигналы, посланные их соплеменниками, на расстоянии в несколько сот километров. Для животных с такими крупными размерами это свойство имеет принципиальное значение. Являясь потребителями большого количества криля, рыбы или кальмаров, киты быстро опустошают пищевую базу. Большая удаленность друг от друга гарантирует китам рациональное использование пищевых ресурсов. В то же время, благодаря низкочастотному каналу связи животные обеспечивают свои зоосоциальные потребности.
Менее развитой способностью к эхолокации наделены тюлени, активные по ночам тенреки и землеройки, а также некоторые мышевидные грызуны, ведущие норный образ жизни. Морские львы (Otariinae) при поиске добычи издают звуки частотой 20-72 кГц. Помимо собственного использования эхолокации, некоторые тюлени прекрасно распознают звуки, издаваемые китообразными. По ним они определяют, приближается ли стадо безобидных планктоноядных китов, или хищных косаток. Землеройки (Soricidae) при помощи гортани испускают крики частотой 30-60 кГц и продолжительностью 5-33 мкс, а тенреки (Tenrecidae) — специализированные щелчки языком. Примитивная эхолокация используется этими зверьками для охоты и исследования местообитаний, на ультразвуковых частотах они также общаются со своими сородичами.
вперед
в оглавление