в оглавление
назад
Миграции
Помимо каждодневных перемещений на более или менее ограниченной территории, некоторые млекопитающие ежегодно совершают путешествия на гораздо более значительные расстояния — кочевки, или миграции. Они носят приспособительный характер и связаны либо с изменениями метеорологических и кормовых условий, либо с определенными стадиями жизненного цикла (размножением, расселением), или же с теми и другими вместе. Нередко кочевки рассматриваются как особый тип передвижений, отличающийся от собственно миграций своей сравнительно небольшой протяженностью. Однако провести грань между кочевками и миграциями очень трудно, тем более, что дальность передвижений у одного и того же вида может в зависимости от обстоятельств изменяться в самых широких пределах. Различают регулярные, периодические и обратимые миграции и нерегулярные, обычно необратимые. Существуют и другие, второстепенные типы миграций: горизонтальные и вертикальные, суточные и сезонные, активные и пассивные. В целом кочевки и миграции у млекопитающих присущи значительно меньшему числу видов, чем у птиц и рыб, и они не столь дальние. В наибольшей мере они развиты у хоботных, копытных, стайных хищников, морских млекопитающих и летучих мышей. У мелких хищных и грызунов кочевничество встречается редко, а у насекомоядных оно практически отсутствует.
Изучение миграций значительно подвинулось вперед с того времени, как животных начали систематически метить. Применение мечения дало ключ к пониманию сущности этих явлений, хотя многие детали требуют еще уточнения. Мечение млекопитающих чаще всего производится при помощи нумерованных металлических колец, продеваемых в ушные раковины (у большинства видов зверей) или закрепляемых на конечностях (например, у летучих мышей — на предплечья), на корне хвоста (у выхухолей (Desmana moschata)) и пр. Для мечения китов используются заостренные полые цилиндры из нержавеющей стали, которыми стреляют из гарпунных ружей. Вонзаясь в толщу подкожного жирового слоя, эти метки не причиняют животным вреда. Применяется также татуировка — накалывание татуировочными щипцами номера на внутренней голой поверхности уха с последующим втиранием туши или краски, а также надрезы или пробивание специальными щипцами отверстий в ушных раковинах и в перепонках лап по условному ключу. Мышевидных грызунов метят, отрезая у них отдельные пальцы (также по особому ключу). Сейчас для мечения широко применятся радиоактивные изотопы, стойкие красящие вещества и портативные радиостанции, прикрепленные к телу животных. Лучшему пониманию миграций также способствуют наблюдения с судов, самолетов и вертолетов, сопровождение на автомобилях, изучение питания прирученных животных и химического состава потребляемой ими пищи и воды, анализ содержимого желудочно-кишечного тракта добытых особей и ряд других методов.

Окольцованная летучая мышь.
Периодические, направленные, исторически сложившиеся миграции наиболее распространены. Чаще всего это правильно повторяющиеся сезонные передвижения по определенным путям или направлениям. Суточные переходы из одних стаций в другие совершаются в основном от мест отдыха к местам кормежек, водопоев или к солонцам. Чем дальше друг от друга отстоят эти места, тем протяженней перекочевки. Кочевой образ жизни особенно характерен для обитателей полузасушливых и засушливых областей. Если растительность слишком скудна для того, чтобы популяция животных могла постоянно кормиться в данном районе, обитатели этих мест вынуждены совершать дальние переходы от одного кормового участка к другому. В тех случаях, когда такие кочевки приобретают регулярный характер благодаря сезонному выпадению осадков, речь идет уже о настоящих миграциях: животные покидают район постоянного обитания, а спустя некоторое время вновь туда возвращаются. Когда приближается пора появления на свет потомства, кочующие виды обычно устремляются во вполне определенные места, а поскольку одно из отличительных свойств настоящих миграций — их связь с размножением, то в таких случаях грань между кочевкой и миграцией почти стирается.
Сравнительно небольшие масштабы имеют вертикальные и отчасти горизонтальные сезонные передвижения типично горных копытных — горных козлов (Capra), баранов (Ovis), горалов (Naemorhedus), серн (Rupicapra rupicapra) и ряда других. С приближением зимы они спускаются на расположенные ниже части склонов с перемежающейся снежностью или переходят в другие участки той же зоны, где снежный покров менее глубок, а значит, более доступны корма. Весной происходит их обратное перемещение в высокогорья, где много пищи и мало кровососущих насекомых. Значительные вертикальные передвижения — от субальпийских лугов до лесов подошвы гор, совершают кабан (Sus scrofa), европейская (Capreolus capreolus) и сибирская (Capreolus pygargus) косули, чернохвостый (Odocoileus hemionus) и благородный (Cervus elaphus) олени, вапити (Cervus canadensis), лось (Alces alces), толсторогий баран (Ovis canadensis). С выпадением снега спускаются в малоснежные предгорья лисица (Vulpes vulpes), дикая кошка (Felis silvestris) и рысь (Lynx lynx). Некоторые летучие мыши — двуцветный кожан (Vespertilio murinus), северный кожанок (Eptesicus nilssonii) и др. — перелетают осенью с гор в низины, а весной обратно. Регулярные горизонтальные миграции в условиях гор хорошо выражены у лося (Alces alces) и сибирской косули (Capreolus pygargus), населяющих Уральский хребет. Зимой они переходят с западного, раньше и обильнее покрывающегося снегом склона хребта на его восточный малоснежный склон, а весной возвращаются обратно.
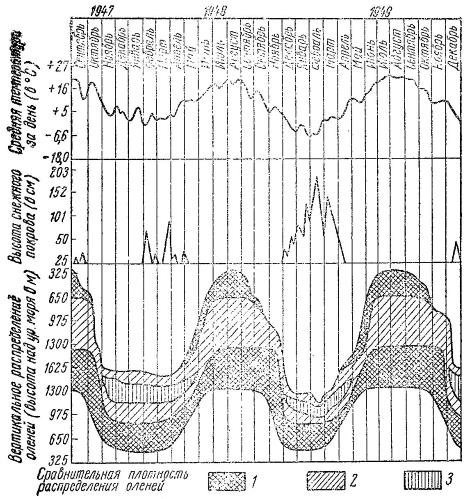
Распределение чернохвостых оленей (Odocoileus hemionus) по вертикальным поясам западного склона хребта Сьерра-Невада (Калифорния) в зависимости от времени года, температуры воздуха и высоты снежного покрова. В многоснежную зиму 1948/49 гг. вся масса оленей сосредоточилась на небольшой площади. В теплую половину года животные распределены широко в большинстве поясов обоих макросклонов хребта. 1 — высокая, 2 — средняя, 3 — низкая плотность населения оленей.
В условиях равнин горизонтальные миграции могут иметь самую различную протяженность. Практически не пытаются переместиться на зиму в более теплые места овцебыки (Ovibos moschatus) и волки (Canis lupus), обитающие на островах Северного Ледовитого океана. Песцы (Vulpes lagopus) на это время года порой даже переселяются еще дальше к северу, чтобы держаться рядом с белыми медведями (Ursus maritimus) и кормиться остатками добытых ими тюленей. Черный медведь (Ursus americanus) не уходит на юг, разве что в самые суровые зимы, когда он переселяется в места, где можно спокойно залечь в спячку, не подвергаясь столь тяжелым испытаниям (при перерасходе энергетических ресурсов он рискует не проснуться весной). Зайцы-беляки (Lepus timidus) на большей части своего ареала ведут оседлый образ жизни, однако в северных частях тундры осенью предпринимают миграции на юг, а весной — в обратном направлении. Протяженность миграционного пути порой доходит до сотен километров; если на пути встречаются реки, зайцы их переплывают. Лоси (Alces alces), ограничивающиеся в малоснежных районах небольшими местными перекочевками, в многоснежных местах предпринимает миграции протяженностью в несколько сотен километров. Прежде, когда американские бизоны (Bison bison) еще были многочисленны, они предпринимали впечатляющие походы, двигаясь по более или менее замкнутому кругу, так что зимой стада иногда оказывались на 650 км южнее своих летних пастбищ.
Далекие сезонные миграции совершают северные олени (Rangifer tarandus). К июлю их стада сосредотачиваются в лесотундре или в северной части тайги, где находят надежную защиту от зимних снежных буранов. С приходом весны олени возвращаются в северные подзоны тундры или даже переплывают на прибрежные острова, где в это время лучше кормовые условия и где их меньше беспокоят кровососущие двукрылые. Скорость стада оленей во время миграции может достигать 80 км/час. При этом в весеннее время скорость выше, а осенью, когда происходит спаривание, она снижается. Переселяющиеся животные из года в год следуют по одним и тем же маршрутам, так что в иных местах их копыта пробили в каменистом грунте тропку полуметровой глубины. Случается, что во время переправ через полноводные реки многие олени тонут; в одном таком месте однажды было найдено 525 их трупов. Самцы северных оленей порой сбиваются в плотные группы, насчитывающие от 100 до 1000 голов, но такие однополые скопления животных неустойчивы, поскольку во время осенней миграции происходит спаривание.
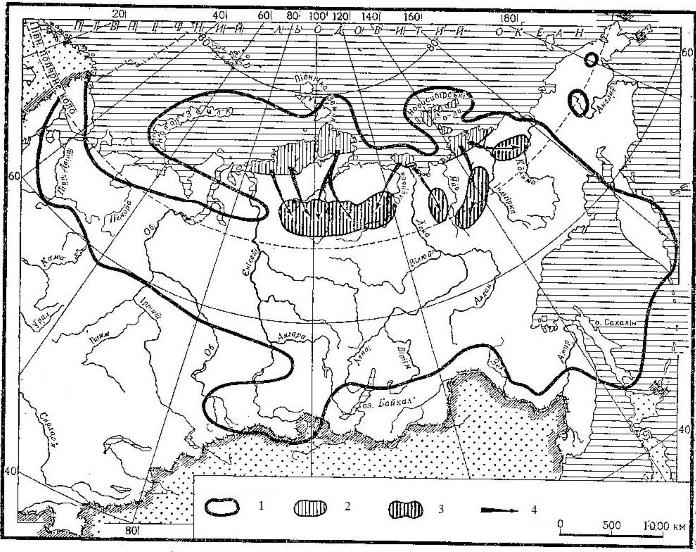
Основные направления кочевок дикого северного оленя (Rangifer tarandus) к местам зимовок в России: 1 — граница распространения; 2 — районы летней концентрации; 3 — районы зимней концентрации; 4 — пути миграции.
Дальше всего перемещаются североамериканские карибу, путь которых в целом может достигать 600-800 или даже 1000 км. Их огромные стада держатся на одном месте только во время рождения молодняка, всего около 14 летних дней. Азиатские олени, уступая американским, порой все же уходят больше чем на 500 км. В Западной Европе дикие северные олени сохранились в небольшом числе только на юге Норвегии. Для них характерны лишь ограниченные местные перемещения. Горные популяции зимой спускаются в долины, а летом держатся преимущественно на больших высотах. Примечательно, что сфера странствий домашних северных оленей Скандинавии значительно шире. Иногда их летние и зимние пастбища разделяет более 250 км нелегкого пути, причем инициатива перехода принадлежит самим оленям, а не их владельцам. При резком снижении численности оленей их миграции становятся незначительными или вовсе прекращаются, потому что из-за отсутствия перевыпаса даже медленно восстанавливающиеся в арктических условиях пастбища не истощаются.
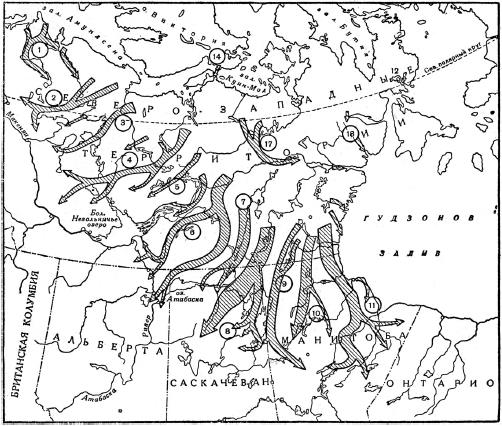
Пути осенних миграций наиболее крупных стад тундровых северных оленей (Rangifer tarandus) в Канаде.
Миграции типичны и для обитателей африканских саванн, где развитие растительности обуславливается чредованием сухих и влажных сезонов. Вообще говоря, саванных млекопитающих в зависимости от их потребности в воде можно разделить на три группы. К первой категории следует отнести животных, постоянно нуждающихся в воде, таких, как гиппопотам (Hippopotamus amphibius), которому требуются местообитания, где всегда достаточно воды. Однако это требование не мешает гиппопотамам в случае засухи или местной перенаселенности совершать утомительные сухопутные переходы из одной реки в другую.
Ко второй категории относятся виды, приспособившиеся к засушливому климату. Потребность в воде у таких животных очень ограничена. Для питья они либо используют поверхностную воду, либо довольствуются влагой, содержащейся в сочных частях растений, корни которых глубоко уходят в землю. Такими частично приспособившимися к сухому климату и немигрирующими млекопитающими являются белый (Ceratotherium simum) и черный (Diceros bicornis) носороги. Их существование зачастую зависит от того, имеются ли поблизости заросли, где можно отыскать сочные побеги, и есть ли тень, в которую можно забраться и долго лежать, сводя потери воды до минимума. В особо засушливые годы носороги в какой-то степени зависят от того рвения, с которым слоны и другие млекопитающие разрывают дно пересыхающих луж и речек, чтобы добраться до почвенных вод.
В третью категорию попадают животные, более или менее широко кочующие в поисках воды. Среди представителей этой группы на первом месте стоит саванный слон (Loxodonta africana), за ними следует африканский буйвол (Syncerus caffer) и, наконец, такие хищники, как лев (Panthera leo), гепард (Acinonyx jubatus), гиеновая собака (Lycaon pictus), пятнистая гиена (Crocuta crocuta), земляной волк (Proteles cristatus), медоед (Mellivora capensis) и большеухая лисица (Otocyon megalotis). Копытные млекопитающие обычно не имеют жестко ограниченной территории, их жизнь проходит в постоянном движении — в промежутках между направленными миграциями они кочуют с места на место.
В прошлом, когда на пути кочевых животных не стояли поселения людей и возможности их передвижения не были ограничены территориями резерватов и заповедными зонами, они каждый год совершали масштабные сезонные миграции. В последние десятилетия поселения и сельскохозяйственные угодья крайне отрицательно сказываются на жизни диких животных, перекрывая пути их миграций и вынуждая животных довольствоваться сравнительно небольшими территориями, на которых в результате часто наблюдается перевыпас и эрозия почв. Эти участки, где ныне только и сохранились многие виды крупных африканских животных, представляют собой остатки угодий, лежащих на путях былых миграций.
Некогда саванные слоны (Loxodonta africana) совершали дальние миграции, чтобы в нужный момент обеспечить себе подходящее убежище и на протяжении всего года иметь в достаточном количестве разнообразную пищу, воду и необходимую им соль. Во время таких миграций стада слонов получали удобную возможность перегруппировываться, и временами случалось наблюдать крупные, до 100 голов, скопления животных. Миграции эти были двух типов: в сезон дождей слоны беспорядочно кочевали с места на место по ограниченной территории, но, кроме того, каждый год совершали направленные миграции, проходя многие сотни километров.
В разные сезоны слоны предпочитают разные местообитания: в сезон дождей они держатся на открытых пространствах, а в засуху скрываются в лесах. Так, близ Килиманджаро слоны в апреле спускаются по северным склонам гор Усамбара и следуют, пересекая плато Ньика, почти до самого побережья. Передвижения слонов в этом и в других районах Восточной Африки определяются наличием пищи. В период размножения животные из разных стад сливаются в особые, репродуктивные стада, которые движутся от южной Лайкипии к Абердарским горам, затем на северо-восток к болотам Лориан, потом на северо-запад к Марсабиту и снова на юг к Абердарским горам. Весь этот кольцевой маршрут протяженностью около 650 км животные проходят за 3 года. В лесах возле Марсабита слоны обзаводятся потомством и домой возвращаются с годовалыми слонятами.
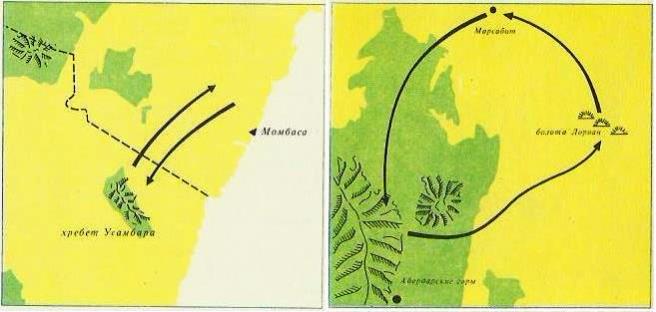
Маршруты миграций саванных слонов (Loxodonta africana).
Вместе с тем, склонность тех или иных животных к миграциям может наблюдаться далеко не на всей протяженности их ареала. Например, один саванный слон из Заира несколько лет жил на участке радиусом не больше нескольких десятков километров. Таким образом, подвижность в значительной мере свойство сугубо индивидуальное. Она различна не только у представителей разных видов, но и у отдельных особей одного и того же вида.
Ежегодно между Кенией, юго-западной Эфиопией и Суданом происходит массовая миграция копытных. Она начинается в мае, когда поднимается уровень воды в болотах верховий Нила, и тогда животные устремляются в юго-восточном направлении к засушливым областям на границе Кении. Основную массу мигрантов составляют кобы (Kobus kob), топи (Damaliscus korrigum) и газели Томсона (Gazella thomsoni). Здесь же можно увидеть большие стада бурчелловых зебр (Equus quagga), газелей Гранта (Gazella granti), канн (Taurotragus oryx) и африканских буйволов (Syncerus caffer). К этим основным стадам присоединяются жирафы (Giraffa camelopardalis), лошадиные антилопы (Hippotragus equinus), ориксы (Oryx gazella), водяные козлы (Kobus ellipsiprymnus) и ряд других. Хотя путь мигрантам преграждает пустыня Туркана, это не мешает им в довольстве проводить здесь три-четыре месяца, пока выпадают сезонные дожди. К концу лета потребность в свежем корме побуждает стада пуститься в обратный путь на север, где после дождей уже появилась новая растительность. В сентябре эти места вновь пустеют, а долины выгорают под палящими лучами солнца.
Однако наиболее широкую известность получили регулярные миграции, совершаемые голубыми гну (Connochaetes taurinus) совместно с бурчеловыми зебрами (Equus quagga), газелями Томсона (Gazella thomsoni) и каннами (Taurotragus oryx) в Национальном парке Серенгети и на соседних охраняемых территориях Танзании. Когда в мае или июне начинается сухой сезон, 1,3 млн. гну покидают низкотравные пастбища на плато в южном секторе Серенгети. Сначала животные спускаются в долины рек, бегущих с гор, и следуют вдоль них на северо-запад к озеру Виктория, а затем на север в Масаи Мара, где раскинулись обширные заливные луга. Здесь гну пребывают в течение всего засушливого периода года, принося в конце осени - начале зимы детенышей. Когда в январе или феврале опять приходят дожди, гну пускаются в обратный путь к низкотравным равнинам на плато. По пути мигрирующие животные пересекают районы, покрытые густыми кустарниковыми зарослями — т. н. буш, часто являющийся рассадником мухи цеце. Общая протяженность миграции составляет около 320 км. Однако мигрируют не все гну. Мелкие стада в течение года постоянно остаются на месте.
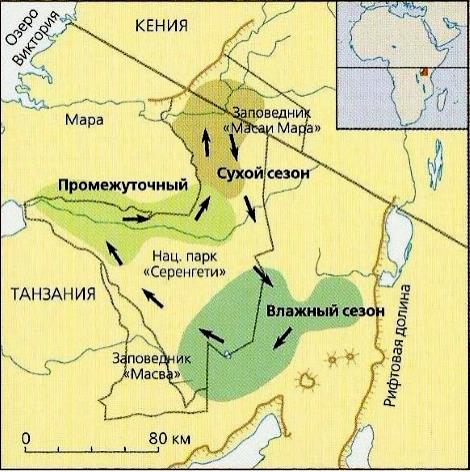
Пути миграций голубых гну (Connochaetes taurinus). Вместе с другими копытными они кочуют в поисках свежей, богатой минералами травы, рост которой зависит от наличия влаги. Когда сезон дождей наступает на юге Африки (с середины зимы до конца весны), животные пасутся на равнинах Серенгети. Когда рост травы здесь останавливается, голубые гну спариваются. В конце весны - начале лета животные перемещаются на северо-запад по направлению к озеру Виктория. В это время дожди перемещаются на север Африки. И хотя травы в приозерных районах не так много, ее здесь больше, чем на равнине. Осень и начало зимы антилопы проводят на севере в Масаи Мара, где сухой сезон более влажный, чем в других местах, поэтому травы больше.
Мигрирующие сообщества данного региона исследуются начиная с 1950-х гг., и за это время по поводу миграций высказывались самые разные предположения. Сначала считалось, что переселяющиеся гну пытаются снизить риск нападения хищников, ведь рядом с ними собирается не менее 3000 львов (Panthera leo) и 9000 пятнистых гиен (Crocuta crocuta). Крупные плотоядные во влажный период следовать за стадами действительно не способны, поскольку привязаны к своим логовищам с детенышами. Кроме того, в подтверждение данной гипотезы указывалось, что хищники поддерживают уровень численности копытных ниже емкости данного местообитания, тогда как мигрирующие гну достигают более высокой плотности и снижают продуктивность угодий. Однако это объяснение имеет два недостатка. Во-первых, было показано, что гиены регулярно удаляются от логовищ на расстояние до 40 км, что позволяет им находиться в постоянном контакте с мигрирующими стадами на большей части их ежегодного маршрута. Во-вторых, таким способом нельзя объяснить, почему местные копытные также не мигрируют, чтобы сбросить с себя бремя хищников. Согласно другой точке зрения, причиной массовых выселений копытных являются сезонные вспышки численности мухи цеце. Однако если это верно, то они, скорее всего, вызывают лишь местные передвижения. Вряд ли серьезной причиной исходов могут быть и черные туфы, которые местами покрывают равнины Серенгети. Наконец, существует предположение, что миграция вызвана недостатком минеральных солей (фосфатов) на заливных лугах, что побуждает стада покидать в сухой сезон свои местообитания и двигаться к югу, к короткотравным равнинам.
К настоящему времени испытание временем и фактами выдержало предположение, что миграции антилоп дают преимущества в использовании различных по качеству и обилию кормовых растений, а также содержащихся в них минералов. Закладывая пробные площадки на пастбищах Серенгети во влажные и сухие сезоны и собирая образцы трав дважды в месяц, исследователи смогли оценить динамику содержания натрия, кальция, фосфора, белков и энергетические запасы в пище на протяжении миграционного цикла, чтобы сравнить их с физиологическими потребностями гну. В большинстве испытаний оказалось, что любой участок пастбищ вполне мог удовлетворить животных зеленой массой на протяжении всего года. Пищевая ценность надземных частей трав была одинаковой на всех сравниваемых участках. Содержание белка колебалось в пределах 3 %, никогда не опускаясь ниже того уровня, когда растительность становится непригодной для поддержания жизнедеятельности, а концентрация натрия и кальция повсюду сохранялась на уровне, соответствующем потребностям кормящих самок гну.

Мигрирующие стада голубых гну (Connochaetes taurinus).
Вместе с тем выяснилось, что содержание фосфора ниже требуемого уровня во всех пробах, исследованных в районе, где не было осадков, но заметно превышает этот уровень на территории, где животные держатся во влажный период. Фосфор и кальций, из которых животные «строят» свой скелет, жизненно необходимы для растущего организма. Однако если кальций содержится в достаточных количествах на всех пастбищах, то доступность фосфора ограничена лишь в определенных районах Серенгети. Как показано на домашнем скоте, недостаток фосфора приводит к снижению рождаемости детенышей, аппетита и производства молока у самок, замедляет рост молодняка, может быть причиной аномалий в строении скелета и зубов, повышает смертность. Для того чтобы глубже изучить роль минералов в миграциях гну, были собраны образцы крови и мочи у временно обездвиженных животных на влажных и сухих территориях. Было обнаружено, что содержание фосфора в сыворотке крови у гну, обследованных на заливных лугах, было вдвое ниже критического уровня, достигая того же минимума, который был обнаружен у домашних овец в экспериментах по искусственному ограничению содержания фосфора в пище. Это приводило к тому, что копытные страстно набрасывались на кости, птичий помет и другие природные источники фосфора.
На основании полученных данных вырисовывается следующая картина. Гну покидают низкотравные равнины по окончании сезона дождей вследствие недостатка пищи и воды на пористых вулканических грунтах, а также из-за увеличения солености воды в немногих оставшихся источниках. Они отправляются на заливные луга, где обильна высокая трава с пятнами молодой поросли, пробивающейся после коротких местных ливней, и имеются постоянные источники пресной воды. Однако за время пастьбы на этих участках у копытных в организме постепенно становится все меньше фосфора. Поэтому, когда на юге Серенгети вновь проливаются дожди, стада животных собираются в обратный путь на низкотравные равнины, чтобы быстро восполнить недостаток этого минерала, которым богата молодая растущая трава. Странствуя в поисках пищи, воды и минералов, копытные одновременно избавляются и от преследования хищников, что приводит к увеличению их численности. Однако это скорее следствие «миграционной политики», чем ее причина. В итоге, чем крупнее стада мигрирующих копытных, тем большую территорию они охватывают своими странствиями, чтобы найти зеленую траву, богатую нужными минералами. Все это, вместе взятое, вносит свой вклад в формирование такого «бродячего» образа жизни у гну. Немигрирующие антилопы, по-видимому, компенсируют общий недостаток фосфора тем, что пасутся меньшими группами. В засушливый период они обычно собираются на легко распознаваемых пятачках с более высоким, чем на соседних участках, содержанием минералов. При такой манере пастьбы они могут избирательно поедать богатую минеральными веществами растительность, например, вокруг термитников.

Голубые гну (Connochaetes taurinus) и бурчелловы зебры (Equus quagga), пересекающие реку во время сезонных миграций, держатся тесными группами, стараясь забраться в центр. Те, что остаются снаружи, больше рискуют подвергнуться нападению крокодилов или потревоженных гиппопотамов (Hippopotamus amphibius).
Мигрирующие и оседлые виды в равной степени хорошо приспособлены к свойственному им образу жизни. Сравнение крупных антилоп, голубых гну (Connochaetes taurinus) и хартбистов (Alcelaphus buselaphus), показывает следующее. Оседлые хартбисты отличаются вытянутой узкой мордой, позволяющей им выбирать нужную пищу среди высокой травянистой растительности, низким уровнем обмена, дающим возможность нормально существовать на низкокалорийном рационе, и совпадением сроков размножения со временем, когда в их организме накоплено больше всего жира. С другой стороны, мигрирующим гну свойственен высокий уровень обмена, необходимый в беспрестанной бродячей жизни, их широкая морда более всего пригодна для ощипывания короткой зеленой травы, а сроки размножения точно подогнаны под сроки миграций.
Регулярные миграции полупустынных и пустынных копытных также зависят от сезонных изменений растительности, а местами — и от характера снегового покрова. Например, сайгаки (Saiga tatarica) в Казахстане летом чаще держатся в северных глинистых полупустынях, а к зиме откочевывают к югу в область менее снежных полынно-типчаковых и полынно-солянковых полупустынь. Джейраны (Gazella subgutturosa) в Средней Азии на зимовку уходят из более многоснежной северной полосы пустынь к югу или в предгорья, где находят лучшие укрытия от непогоды и больше доступных кормов.
Переселяясь, многие наземные звери могут далеко заплывать в море. Даже обыкновенная ласка (Mustela nivalis) однажды проплыла 1,5 км. Сибирская косуля (Capreolus pygargus) и вапити (Cervus canadensis) нередко преодолевают морские проливы. На еще большие расстояния от берега удаляются полуводные млекопитающие. Выдра (Lutra lutra) заселила острова, отстоящие от материка более чем на 15 км, а в отдельных случаях — до 20 км. Гиппопотам (Hippopotamus amphibius) осиливает путь более чем в 30 км между Африкой и Занзибаром. Почти на таком же расстоянии от ближайшей суши встречали белого медведя (Ursus maritimus).

Максимальные расстояния, на которые отдалялись от берега некоторые млекопитающие.
Своеобразны сезонные миграции летучих мышей, населяющих полосу лесов и лесостепей в Евразии и Северной Америке. С приближением зимы они вынуждены либо впадать в спячку в непромерзаемых зимних убежищах, либо улетать в более теплые области, где возможна их зимняя активность. Виды, днюющие в заброшенных зданиях, на чердаках, в дуплах деревьев и других укромных местах, в большинстве случаев здесь и зимуют, а в тех районах, где морозы особенно жестоки, отыскивают пещеры или штольни шахт, достаточно глубокие для того, чтобы в них все время сохранялась постоянная, примерно нулевая, температура. В восточных областях США таких убежищ немного и они расположены далеко друг от друга, за исключением районов выхода известняковых пород, где часто встречаются обширные системы подземных пещер. Мыши, образующие в этих пещерах огромные зимние скопления, прилетают в основном из соседних районов, куда они возвращаются с наступлением весны. Эти перелеты относительно коротки. Те виды летучих мышей, которые живут на деревьях, прячась среди листьев или цепляясь за стволы, напротив, не забиваются в пещеры, а совершают дальние перелеты к югу, очень похожие на сезонные миграции насекомоядных птиц. Перелеты совершаются в основном в сумерках, по пути животные охотятся на насекомых. Большинство летучих мышей успевает мигрировать за свою жизнь несколько раз.
У восточного красного (Lasiurus borealis) и обыкновенного серого (Aeorestes cinereus) волосатохвостов из Северной Америки длинные узкие крылья и они так хорошо летают, что это позволяет им совершать зимние миграции на юг США, а весной возвращаться обратно. Оба вида можно встретить осенью на Бермудских островах, в 1000 км к юго-востоку от Нью-Йорка, а красных волосатохвостов ловили в августе в 385 км от полуострова Кейп-Код. Большое число летучих мышей вынуждены совершать перелеты над морем, хотя до тех пор, пока это возможно, они предпочитают лететь вдоль берега. Североамериканский серебристый гладконос (Lasionycteris noctivagans) также совершает более или менее регулярные ежегодные миграции, а мексиканский складчатогуб (Tadarida brasiliensis) мигрирует на 1280 км с юга США в центральные районы Мексики.
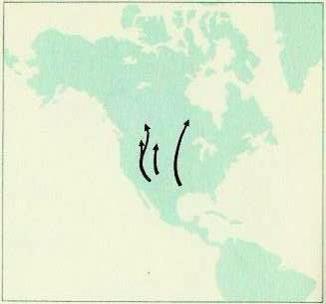
Миграционные маршруты мохнатого волосатохвоста (Lasiurus cinereus).
Расстояния между зимними и летними местообитаниями у разных видов и даже у разных особей летучих мышей сильно различаются. Для европейского обыкновенного ушана (Plecotus auritus) наибольшее расстояние составляет порядка 260 км, наименьшее — около километра, основная же масса мышей этого вида мигрирует на расстояния от 40 до 80 км. Нетопырь-карлик (Vespertilio pipistrellus), окольцованный в районе Днепропетровска (Украина) в июне 1939 г., был пойман в сентябре того же года в южной части Болгарии (у Пловдива) на расстоянии по прямой более 1000 км от места кольцевания. Северный кожанок (Eptesicus nilssoni), окольцованный в конце июня 1939 г. в районе Днепропетровска, через несколько недель был пойман возле Пловдива (Болгария), на расстоянии 1150 км. Лесной нетопырь Натузиуса (Vespertilio nathusii) с кольцом, надетым в Воронежском заповеднике в июле 1958 г., был найден в ноябре того же года вблизи Стамбула (Турция). Он преодолел расстояние около 1750 км. В 1935 г. в пещере возле Дрездена было окольцовано около 600 находившихся в спячке особей рыжей вечерницы (Nyctalus noctula). При этом три из них попались вторично: одна была из Гановера, вторая из Сухи (Польша), а третья из Литвы и пролетела около 750 км. Особи этого же вида, окольцованные летом в Воронежской области, были обнаружены на зимовках в предгорьях Кавказа, в Крыму и в Закарпатье, т. е. на расстояние 1000 и более км. Максимальное же расстояние, преодоленное одной меченой рыжей вечерницей, составило 2350 км. Пока для рукокрылых это рекорд. Таким образом, летучие мыши одного вида могут и впадать в спячку на месте, и мигрировать.
Обыкновенный длиннокрыл (Miniopterus schreibersi) и большой подковонос (Rhinolophus ferrumequinum) обычно проводят зиму в спячке, скрываясь в пещерах, расположенных далеко от их летних местообитаний, то же самое можно сказать о малом подковоносе (Rhinolophus hipposideros) и средиземноморском нетопыре (Pipistrellus kuhlii). Тысячи двухцветных кожанов (Vespertilio murinus) проводят лето в старых укреплениях крепости Эг-Морт, у устья Роны, а осенью покидают эти места. Скорее всего, подземные укрепления не подходят для спячки, и летучим мышам приходится искать более закрытые убежища, с постоянной температурой. Однако никаких свидетельств о том, что европейские летучие мыши зимуют в Африке, как это делают многие птицы, не существует. Тем не менее, в Европе известны случаи, когда летучих мышей видели летящими днем вместе со стаями ласточек и других птиц.
Осенний перелет евразиатских и североамериканских летучих мышей совершается весьма неспешно, продолжаясь с середины августа до ноября, но самый разгар его приходится на сентябрь. Факт миграции некоторых летучих мышей с юга Южной Америки к северу с приближением зимы не вызывает сомнений, но о том, как они происходят в этой части света, почти ничего не известно.
Живущие в тропиках крыланы совершают достаточно регулярные массовые перелеты в поисках поспевающих плодов. Пальмовый крылан (Eidolon helvum) вначале следует за дождями в леса Западной Африки и к низовьям Нила, а затем, в конце дождливого сезона, возвращается на юг континента. Фруктоядные австралийские летучие лисицы — сероголовая (Pteropus poliocephalus), бавеанская (Pteropus alecto) и очковая (Pteropus conspicillatus) — в течение летне-осеннего сезона постепенно перелетают из Квинсленда в Новый Южный Уэльс. Более мелкая австралийская красная летучая лисица (Pteropus scapulatus), кормящийся в основном на цветках, совершает миграции менее регулярно, поскольку необходимость в них возникает, лишь когда прекращается цветение эвкалиптов и других деревьев, чьим нектаром питаются эти животные. Наоборот, перелеты сероголовой летучей лисицы вызываются в основном сезонным созреванием мелких плодов, особенно дикого инжира, составляющего основу их рациона.
Перелеты летучих мышей имеют много общего с перелетами птиц и связаны с аналогичным наследственным инстинктом. У бухарского подковоноса (Rhinolophus bocharicus), например, осенний отлет начинают молодые особи, к тому же летящие на большом расстоянии друг от друга, что полностью исключает возможность научения. Кроме того, как и у многих птиц, стаи летучих мышей во время миграций состоят из животных одного пола. Такое разделение без сомнения указывает на то, что миграции связаны с поисками пищи, а не с размножением.
вперед
в оглавление