в оглавление
назад
Миграции (продолжение)
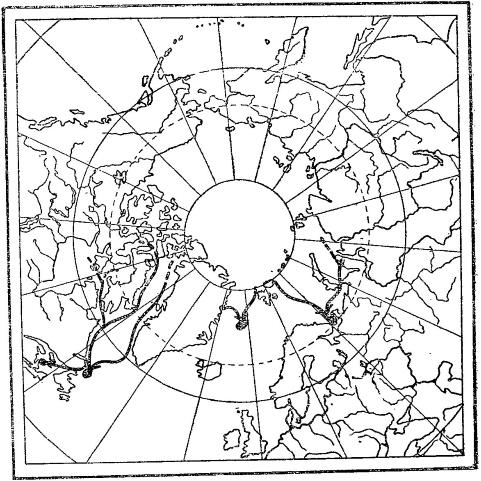
Направление осенних миграций трех стад гренландского тюленя (Pagophilus groenlandicus) из районов нагула к местам щенки и линьки. Тюлени беломорского стада трогаются в путь в конце сентября, в декабре они появляются у берегов Мурманска и в Горле Белого моря.
В отличие от рукокрылых, сезонные миграции морских млекопитающих связаны как с питанием, так и с размножением. Ластоногие широко кочуют, но для размножения каждый год возвращаются на одни и те же ограниченные участки суши или покрытые льдами моря. Так, гренландские тюлени (Pagophilus groenlandicus) образуют три стада, проводящие летний период нагула в прибрежных водах Канадского архипелага и Гренландии, а также у кромки льдов от Шпицбергена до Земли Франца Иосифа и отчасти в Карском море. В конце зимы одно стадо мигрирует к Ньюфаундленду, другое — к острову Янмайену, третье — в Горло Белого моря. Здесь на морских льдах у тюленей находятся места щенки и спаривания. Северные морские котики (Callorhinus ursinus) образуют два стада — азиатское и американское. Первое на зиму мигрирует от Командорских островов и острова Тюленьего к восточным берегам Кореи, островов Хоккайдо и Хонсю, второе — от островов Прибылова вдоль западного берега Америки в район Южной Калифорнии. После зимовки в южных морях котики возвращаются на острова в Беринговом море, где у них происходит щенка, выкармливание молодых и спаривание. Путешествие котиков в оба конца составляет около 10 000 км, в море они проводят до 8 месяцев в году. Миграции моржей (Odobenus rosmarus) с юга на север весной и обратно осенью обусловлены в основном тем, что мелководья, где эти животные могут добывать донных моллюсков, при появлении льдов становятся для них недоступными, и отчасти связаны с продвижениями к местам, удобным для размножения. Самые далекие миграции из ластоногих (и млекопитающих в целом) совершают северные морские слоны (Mirounga angustirostris). Дважды в году их стада перемещаются между местами кормежки в прибрежных мексиканских и калифорнийских водах и местами размножения на побережье Аляски и в северной части Тихого океана, преодолевая в общей сложности расстояние в 20 000 км.
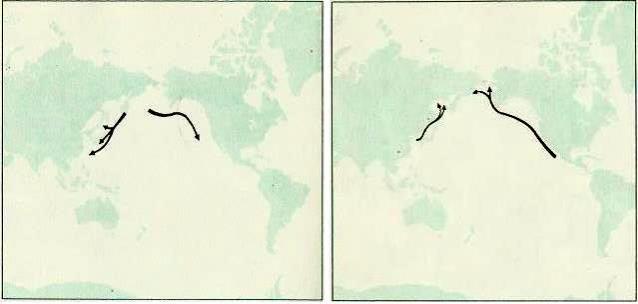
Маршруты весенних миграций двух популяций северного морского котика (Callorhinus ursinus) (слева) и двух популяций серого кита (Eschrichtius robustus) (справа).
Перемещения китообразных также зависят от необходимости менять места нагула и переходить в районы, благоприятные для выведения молодняка. Большинство китов-планктонофагов совершает хорошо выраженные миграции к кромке полярных льдов, где массово развивается зоопланктон. Рачки и привлеченные ими рыбы служат китам обильным нажировочным кормом. На богатых полях питания эти животные образуют достаточно плотные скопления, именно в таких местах раньше производился наиболее успешный их промысел. К осени (в северном полушарии — к весне) зоопланктон исчезает, льды смыкаются, киты разбиваются на небольшие группы и уходят зимовать в теплые воды. Здесь кормовая база гораздо беднее, планктоном и рыбой изобилует лишь район Галапагосских островов, Карибское море, северо-западное побережье Африки, Аденский и Бенгальский заливы, а также ряд других мест. Синие киты (Balaenoptera musculus), финвалы (Balaenoptera physalus), сейвалы (Balaenoptera borealis) и горбатые киты (Megaptera novaeangliae) приплывают сюда из полярных морей, преодолевая более 6000 км через бедные пищей пространства. Средняя скорость мигрирующего кита составляет 7 км/ч. Во время путешествия эти животные очень мало едят — по меньшей мере 4 месяца их желудок почти пуст, вследствие чего они сильно худеют. Кроме того, переход из холодных вод в теплые необходим китам для щенки, поскольку их новорожденные не имеют развитой жировой прослойки и могут существовать только в условиях достаточно высоких температур. Спустя полгода киты опять возвращаются в полярные воды.
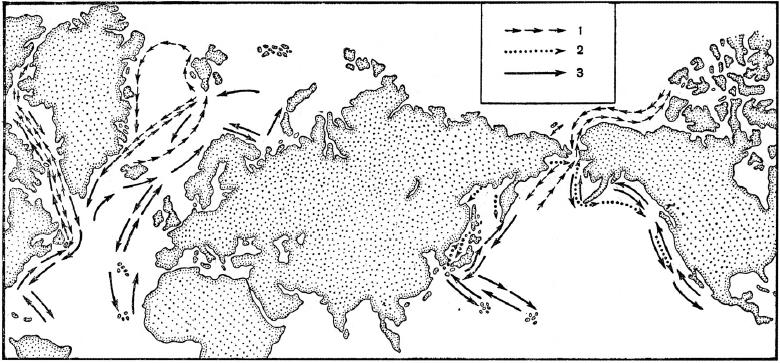
Схема миграций китов: 1 — гренландский кит (Balaena mysticetus); 2 — серый кит (Eschrichtius robustus); 3 — горбатый кит (Megaptera nodosa).
Пути миграции синих китов и финвалов пролегают не так близко к побережьям материков, как у горбатых китов. Существует мнение, что беременные самки финвалов дольше остаются в антарктических водах, чем беременные самки синих китов, и на несколько недель позже производят на свет потомство, однако точное расположение зимних квартир и путей миграции финвалов и синих китов неизвестно; кроме того, не все особи мигрируют зимой в теплые воды. Хотя финвалы, синие и горбатые киты иногда пересекают экватор, считается, что их южные популяции изолированы от популяций, обитающих в северном полушарии. Ежегодно в тропических водах обнаруживаются представители обеих групп, однако они проводят здесь разные месяцы и потому не перемешиваются.

Схема миграций горбатых китов (Megaptera nodosa). Популяции Северного полушария проводят лето на кормовых территориях в северной части Атлантического и Тихого океанов, где отъедаются на косяках мелкой рыбы и накапливают жир для последующего долгого пути. С приближением зимы они мигрируют примерно на 5000 км к югу в теплые, но малокормные воды у Карибских островов, около Западной Африки, Японии, Гавайских островов и у берегов Мексики. Здесь после спаривания самки приносят потомство. В это время взрослые особи практически не питаются, существуя за счет накопленного под кожей жира, а детеныши пьют материнское молоко. В конце концов, они возвращаются в богатые кормом полярные воды. Горбатые киты Южного полушария следуют той же схеме, питаясь летом, с ноября по апрель, в богатых крилем водах возле берегов Антарктиды. Затем они мигрируют к экватору, чтобы перезимовать в тропических водах у берегов Южной Африки, Австралии, Новой Зеландии и Южной Америки. Таким образом, южная крайняя точка их ежегодных миграций отстоит от северной не менее чем на 40 ° широты. Популяция в северной части Индийского океана весь год может быть оседлой.
Основные места нагула кашалотов (Physeter macrocephalus) находятся в тропическом, субтропическом и отчасти в умеренном поясах. Его рацион в основном состоит из головоногих моллюсков, для которых воды с низкой температурой и пониженной соленостью неблагоприятны. Самки кашалотов за редкими исключениями не выходят за пределы тропической и субтропической зон, где щенятся и растят своих детенышей, а кочующие крупные самцы в летнее полугодие в северном полушарии достигают Баренцева и Берингова морей, а в южном — Антарктики. Молодые самцы, не способные удерживать гарем или еще не достигшие половозрелости, совершают те же путешествия несколько позднее. Миграции на значительные расстояния совершают и различные виды рыбоядных дельфинов.

Во время миграций горбатые киты (Megaptera nodosa) периодически вертикально поднимают голову из воды и, кроме того, поворачиваются, чтобы оглядеться. Возможно, что так они ищут известные им береговые ориентиры.
Другие китообразные держатся на более ограниченных акваториях. Гренландские киты (Balaena mysticetus), белухи (Delphinapterus leucas) и нарвалы (Monodon monoceros), например, обитают и размножаются в арктических водах, тогда как карликовые киты (Neobalaena marginata) и дельфины Коммерсона (Cephalorhynchus commersonii) встречаются лишь в южных полярных водах, правда, на огромных пространствах. Высоколобые бутылконосы (Hyperoodon ampullatus) и атлантические ремнезубы (Mesoplodon bidens) живут в северной половине Атлантики, щенясь в теплой сфере влияния Гольфстрима. Серые киты (Eschrichtius robustus) в зимние для северного полушария месяцы производят на свет детенышей в укрытых лагунах, расположенных по берегу Калифорнийского залива, а с наступлением тепла начинают продвигаться вдоль берега на север. Летом они плавают в северной части Тихого океана и в арктических водах, питаясь обильными скоплениями планктона.
Рыбоядные киты и дельфины мигрируют в зависимости от гидрологических условий и распределения стадных рыб. Так, белухи Карского стада в летние месяцы держатся в прибрежных водах Карского моря от Новой Земли до Северной Земли. Здесь они охотятся за сайкой, а в опресненных заливах и устьях рек главным образом за сиговыми рыбами, образующими большие косяки перед ходом на нерестилища. В конце ноября - начале декабря белухи в массе уходят из Карского моря на запад через проливы между островами Новой Земли и проводят зиму у кромки льдов в восточной и юго-восточной части Баренцева моря. Нередко массовый ход этих дельфинов через проливы совпадает с миграцией сайки, следом за которой часто идут и стада гренландских тюленей (Pagophilus groenlandicus).
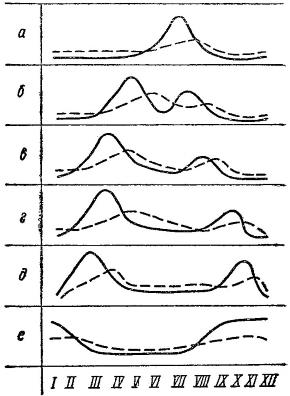
Схема сезонных изменений биомассы планктона в морях и океанах на разных широтах. Сплошная линия — фитопланктон, прерывистая — зоопланктон: а — арктическая область; б — субарктическая; в — северная бореальная; г — южная бореальная; д — субтропическая; е — тропическая.
Утверждать, что животные совершают подобные путешествия, повинуясь сознательному побуждению, конечно же, нельзя. Стремление к миграции, как и многие другие формы поведения, основываются на физиологических стимулах. Главными двигателями в этом случае являются подчиняющаяся сезонному ритму деятельность желез внутренней секреции и другие физиологические изменения в организме животного. Именно они и диктуют ему, что делать.
В связи с миграциями неизбежно возникает вопрос: каким же образом животные находят дорогу? Прежде чем пытаться ответить, приведем несколько подтвержденных экспериментами примеров возвращения млекопитающих в родные места. Так, известно, что оленьи хомячки (Peromyscus sp.), индивидуальный участок которых обычно составляет около 100 м в диаметре, выпущенные на волю в 3 км от места их постоянного обитания, возвращались домой через 48 часов. Способность к возвращению у летучих мышей еще более замечательна. Рыжие вечерницы (Nyctalus noctula) прилетали на свой индивидуальный участок за 24 часа с расстояния 45 км. Что касается более дальних перелетов, то имеются достоверные сведения о случаях возвращения летучих мышей домой с расстояний до 265 км. Во всем этом есть много общего со способностью к ориентировке, присущей домашним животным, особенно собакам (Canis lupus familiaris) и кошкам (Felis silvestris catus), которые нередко находят обратный путь из мест, удаленных от дома на 15-40 км, а то и больше.
Чтобы попытаться понять механизм этих перемещений, необходимо в первую очередь рассмотреть гипотезы, предполагающие существование у животных «здравого смысла». Например, собака, которую увезли на поезде за 30 км, бежала назад вдоль железнодорожного пути, пока не достигла знакомой территории. Конечно, не следует сбрасывать со счетов возможность счастливой случайности. Вполне вероятно, что в отдельных случаях животное искало путь наугад в разных направлениях, пока не натыкалось на приметы знакомой местности. Но эти объяснения применимы не ко всем подобным случаям и уж определенно не подходят к массовым сезонным перемещениям.
Некоторый свет на эту проблему проливают опыты, проведенные с птицами. Представляется вполне вероятным, что мигрирующие птицы во время своих долгих перелетов в значительной степени ориентируются по звездам. Млекопитающие, в особенности летучие мыши, при дальних миграциях также, возможно, пользуются средствами небесной навигации. Следует принять во внимание и существование давно используемых привычных маршрутов, и передачу знания примет местности от поколения к поколению. Но тем не менее приходится признать, что в самой сути явления кроется еще много непонятного.
В отличие от регулярных обратимых миграций нерегулярные необратимые миграции, или эмиграции, представляют собой массовые выселения животных, как правило, без последующего возвращения на прежние места обитания. Вызываются такие выселения резкими ухудшениями жизненных условий и бескормицей в связи с перенаселенностью, лесными или степными пожарами, сильными засухами, наводнениями, чрезмерным многоснежьем и другими причинами.
Одним из наиболее драматических примеров эмиграции являются выселения южноафриканских антилоп спрингбоков (Antidorcas marsupialis), совершаемые когда-то в абсолютно неправдоподобных количествах. Во время наиболее крупных миграций стада по 10-20 тыс. голов каждое собирались вместе в колонны, насчитывавшие сотни миллионов животных. В 1849 г. городок Бофорт-Уэс был наводнен стадами спрингбоков, которых сопровождали бурчелловы зебры (Equus quagga), беломордые бубалы (Damaliscus pygargus), белохвостые гну (Connochaetes gnou), канны (Taurotragus oryx) и антилопы ряда других видов. Насколько хватало взгляда, животные заполнили улицы и сады городка. Когда через три дня нашествие схлынуло, местность выглядела так, будто по ней прошелся пожар. Описана еще одна грандиозная миграция, во время которой колонны копытных, плотность которых, правда, не была постоянной, несколько дней двигались через одну и ту же местность. Многие животные, в особенности старые и молодые, погибали, но уцелевшие возглавляли шествие, пока стада не достигли моря, где животные тонули в столь огромных количествах, что на протяжении почти 50 км пляж был завален горами трупов. Миграция, вероятно, была вызвана голодом, последовавшим за вспышкой численности популяции. После одной такой миграции, когда спрингбоки, подобно наводнению, огромными массами заполнили долины и склоны холмов в Намакваленде, а затем достигли побережья Атлантического океана, бросились в волны и погибли.
Местные жители подразделяют спрингбоков на две категории — «хоу-боккеров», постоянно обитающих на одних и тех же участках вельда, и «трек-боккеров», занимающих менее благоприятное местообитание и являющихся источником описанных выше нашествий. При этом предполагается, что основным фактором, стимулирующим нашествия спрингбоков, в особенности спрингбоков пустыни Калахари, способных существовать без воды, на сухой траве и сочных корешках, является не засуха, а недостаток пищи в сочетании с ростом численности популяции. Во время нашествий обычное поведение спрингбоков меняется. Животные становятся беспокойными, беспричинно суетятся, то и дело срываются с места и скачут куда попало, пока не сбиваются в стада. При этом они утрачивают свою природную пугливость настолько, что даже заходят в деревни и города. В XIX в. во время великих нашествий спрингбоков некоторые животные утоляли жажду прямо из фонтанов на городских улицах. Последнее нашествие спрингбоков наблюдалось в 1954 г. и было сравнительно небольшим.
Массовые эмиграции каролинских белок (Sciurus carolinensis), обитающих на востоке США, наблюдались еще во времена колонизации североамериканского континента. Зверьки стремились вперед, поедая на своем пути все, что придется им по вкусу, и опустошая кукурузные и пшеничные поля фермеров. Самое масштабное переселение белок было зарегистрировано в 1842 г. в Висконсине и длилось 4 недели. Основываясь на данных очевидцев, было подсчитано — возможно, с некоторыми преувеличениями — что в этом нашествии принимало участие около 450 млн. белок. Какова бы ни была действительная цифра, несомненно, что в поход были вовлечены огромные полчища зверьков. Во время передвижений серых белок в 1930-х гг. в штатах Коннектикут и Нью-Йорк тысячи животных гибли на проезжих дорогах или тонули, переправляясь через реку Коннектикут.
Однако наиболее известны нашествия леммингов (Lemmini) — небольших грызунов, населяющих тундры и редколесья Евразии и Северной Америки. Так, у норвежских леммингов (Lemmus lemmus) каждые 3-4 года число новорожденных в пометах оказывается больше обычного, молодняк быстрее достигает половой зрелости и сам приступает к размножению. Причины подобных вспышек численности окончательно не выяснены, но можно предположить, что все начинается с группы леммингов, попавших в исключительно благоприятные условия. Интенсивно плодясь, популяция достигает максимальной численности к концу лета, когда и без того скудные запасы пищи уже сильно истощены. Голодающие животные, обитающие на границе перенаселенных областей, начинают разыскивать пищу на стороне. В первую очередь это молодые самцы, потерпевшие неудачу в попытках занять собственный индивидуальный участок; за ними следуют другие мигранты. Постепенно образуются скопления в районах, удаленных от наиболее страдающих от голода популяций. Со временем выселение принимает все более широкие масштабы, так как подходящие биотопы вокруг либо уже заняты, либо полностью опустошены прошедшими зверьками.
Выселение леммингов длится много дней или даже недель и носит стихийный характер. Множество зверьков движется поодиночке на некотором расстоянии друг от друга в общем направлении, которого придерживаются с большим постоянством. Натолкнувшись на препятствие, лемминги не обходят его, а идут напрямик, стараясь преодолеть. При этом они очень возбуждены, утрачивают чувство страха и не боятся привлечь к себе внимание. Лемминги кидаются с горных круч, скатываются по склонам в канавы и ямы; заходят в города и даже внутрь жилищ, где становятся добычей собак и кошек; дерутся друг с другом и нападают на всякого, кто преградит им путь, несмотря на сильное истощение и отсутствие шансов на успех.
Достигнув протоки или реки, лемминги бегают взад и вперед вдоль побережья, отыскивая пологое место с медленным течением, где водное пространство пересечь безопаснее. Найдя такое место, они спрыгивают в воду и быстро плывут к другому берегу; там они встряхиваются и, как ни в чем не бывало, продолжают свое путешествие. Лемминги — хорошие пловцы, однако большие реки и озера переплыть удается лишь немногим из них. Достигнув берега моря, зверьки бросаются и в него. Их спасает только, если поблизости окажется остров или противоположный берег фьорда. Так, известны случаи, когда лемминги вплавь пересекали фьорды шириной более 4 км. Не удивительно, что их тела иногда выбрасывает на берег прилив. В 1868 г. во фьорде Тронхейм пароход шел по морю, на протяжении 3-5 км сплошь усеянному плывущими леммингами. По приблизительным подсчетам, их число выражалось в миллионах, но это была всего лишь отдельная ветвь огромной орды, размеры которой установить вообще не представлялось возможным. Похожие наблюдения были сделаны и позднее.
Определенные пути миграции у леммингов отсутствуют. Общее направление движения в некоторой степени определяется рельефом местности, по которой зверьки путешествуют. В Скандинавии, например, лемминги населяют склоны гор и плоскогорья, лежащие выше пояса леса, и отдельные популяции разделены высокими вершинами или глубокими долинами. Поэтому направление миграции весьма ограниченно, и полчища леммингов устремляются вниз, следуя определенными путями, на которых образуют огромные скопления. Гренландские лемминги (Dicrostonyx groenlandicus) во время миграций идут по льду, покрывающему море, от островов, лежащих в 80 км от берега, к материку. Более мелкие лесные лемминги (Myopus schisticolor) также претерпевают вспышки численности и мигрируют в лесах, но их перемещения менее заметны, к тому же животные быстро достигают новых кормовых угодий. Иногда «поход» зверьков прерывается в результате их массовой гибели от инфекционных заболеваний. Поредевшие ряды леммингов успокаиваются и останавливаются, начинают размножаться. И все же такая колония обычно очень скоро вымирает.
С высокой плотностью популяции и сокращением кормовой базы связаны, по-видимому, наблюдающиеся время от времени выселения кабанов (Sus scrofa), северных оленей (Rangifer tarandus), лосей (Alces alces) и сибирских косуль (Capreolus pygargus). В частности, имеется много указаний на вынужденные выселения зайцев и копытных, связанные с многоснежьем и буранами. Так, в чрезвычайно снежную зиму 1923 г. в Мариупольском районе (Украина) наблюдалось движение огромного количества зайцев-русаков (Lepus europaeus) на юг, причем животные продолжали следовать и по льду замерзшего Азовского моря. Они погибали здесь массами. В феврале 1928 г. в районе степного хозяйства Аскания-Нова (Украина) разразился сильнейший буран, длившийся двое суток. В результате тысячи русаков, спасаясь от голодной смерти, ринулись в поселок Аскания и заполнили его площадь и дворы; в степи было обнаружено много мертвых зайцев. Большое количество ослабевших животных было выбито населением просто палками. В феврале 1952 г. на льду и на островах северного побережья Каспия появились тысячные стада сайгаков (Saiga tatarica). С потеплением и таянием льда копытные оказались отрезанными от берега, что привело к их массовой гибели от голодовки. В Сибири в многоснежную зиму 1913/14 гг. лоси (Alces alces), изюбры (Cervus canadensis xanthopygus) и сибирские косули (Capreolus pygargus) были вынуждены оставить места постоянного обитания и искать себе пропитание вблизи человеческих поселений; отдельные особи при этом заходили даже в деревни. Сильные засухи и бескормица регулярно вызывают откочевку джейранов (Gazella subgutturosa).
Особую категорию представляют передвижения расселяющихся молодых животных после распада семей. Это обратимые и полезные для вида передвижения. Например, эмиграции белок (Sciurus vulgaris) часто начинаются с расселения подросших детенышей. Если данный год на большой лесной площади неурожайный на корма, следом за молодыми или одновременно с ними начинают откочевывать и взрослые. Вскоре в поток эмигрантов вливаются тысячи зверьков. Сезонные и многолетние колебания частоты эмиграции белок определяются в основном условиями питания, реже изменениями защитных свойств стаций, вызванных лесными пожарами, метеорологическими условиями и т. п. Во время выселений белки проходят расстояния до 500 км; наибольшее известное расстояние, преодоленное отдельной особью, составляет 240 км. Длительность эмиграции может достигать 6 месяцев. Ширина фронта, которым движется масса эмигрирующих белок, достигает 300 км и более. Подобно другим мигрирующим животным, белки бесстрашно форсируют встречающиеся на пути водоемы. Так, в 1867 г. они вплавь пересекли реку Чусовую и достигли Нижнего Тагила на Урале.
У мелких мышевидных грызунов массовые выселения представляют более редкое явление, они кратковременны и не захватывают больших пространств. Осенью 1940 г., при массовом размножении грызунов в степях Предкавказья наблюдались крупные передвижения домовых мышей (Mus musculus). В 1945 г. при подобных обстоятельствах на территории Центрально-черноземной полосы отмечались массовые передвижения полевок — обыкновенной (Microtus arvalis) и водяной (Arvicola terrestris).
Массовые истерии и потеря осторожности у мигрирующих животных может быть связана с расстройством гормонального баланса в результате перенаселенности или недостатка пищи. Перевыпас, болезни и стресс также могут быть существенными факторами. И хотя, например, мигрирующие спрингбоки (Antidorcas marsupialis) бывали иногда хорошо упитаны, но куда чаше оказывались истощенными и очень страдали от паразитов. Во время нашествия 1896 г. они не остановились, достигнув долины реки Оранжевой, а проследовали дальше в сухие местообитания, где и произвели на свет потомство. Вообще, выведение потомства во время миграций для животных в порядке вещей.
Сопряженными миграциями называют связанные друг с другом передвижения двух или нескольких видов. Таковы, в частности, миграции и эмиграции хищников, связанные с передвижениями их жертв. В основе своей это кормовые переселения, но стимулирует их не бескормица, а обилие пищи. Многие хищники, следуя за мигрирующими животными, охотятся за ними, другие подбирают остатки трапезы более сильных собратьев или поедают трупы павших животных. Так, за горными копытными следуют снежные барсы (Panthera uncia) и волки (Canis lupus), за кабанами (Sus scrofa) — тигры (Panthera tigris), за сибирскими косулями (Capreolus pygargus) и вапити (Cervus canadensis) — рыси (Lynx lynx), волки (Canis lupus) и росомахи (Gulo gulo), за северными оленями (Rangifer tarandus) — тундровые волки (Canis lupus), песцы (Vulpes lagopus) и росомахи (Gulo gulo), за сайгаками (Saiga tatarica) и джейранами (Gazella subgutturosa) — волки (Canis lupus), за африканскими копытными — львы (Panthera leo), леопарды (Panthera pardus), гепарды (Acinonyx jubatus), пятнистые гиены (Crocuta crocuta), гиеновые собаки (Lycaon pictus) и африканские волки (Canis anthus). Мигрирующих леммингов сопровождают горностаи (Mustela erminea), волки (Canis lupus), песцы (Vulpes lagopus), а также оазличные хищные птицы. Их охотно поедают даже северные олени (Rangifer tarandus), вследствие чего северным скотоводам бывает нелегко собрать свои стада.
Из приведенных примеров можно сделать вывод, что направления путей расселения определяются экологическими руслами, которые могут служить и путями сезонных миграций соответствующих видов. Впрочем, в некоторых случаях особенности передвижения млекопитающих полного объяснения еще не нашли. К таковым, например, относятся переходы песцов (Vulpes lagopus) на расстояние более 1000 км за один зимний сезон. По-видимому, они представляют собой крайние варианты обычных осенних выселений.
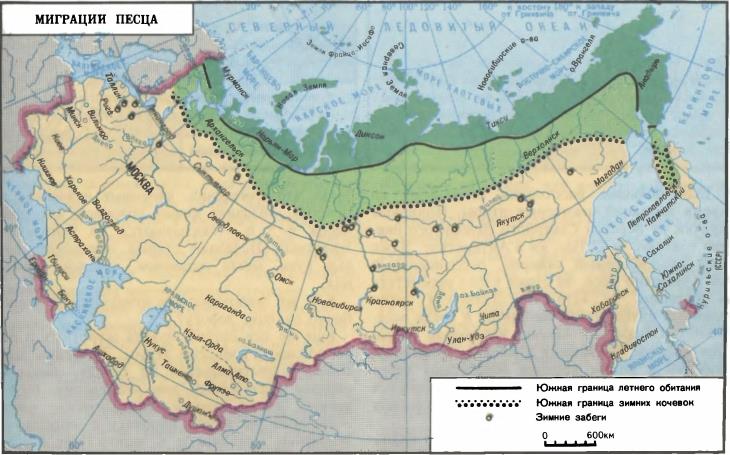
Карта летнего и зимнего распространения песца (Vulpes lagopus).
Все рассмотренные примеры относятся к категории активных миграций. Пассивные миграции также важны, хотя и встречаются у млекопитающих сравнительно редко. К ним можно отнести перемещения моржей (Odobenus rosmarus) на дрейфующих льдинах; нередко на большие расстояния на льдах уносит белых медведей (Ursus maritimus) и песцов (Vulpes lagopus). Совершенно случайный характер носят перемещения мелких млекопитающих на речных льдинах во время ледохода, на плавучих островках и бревнах в половодье. Кроме того, животные могут перемещаться на различных видах человеческого транспорта, вследствие чего случайно или вполне намеренно завозятся в новые для них местообитания.
Таким образом, миграция должна рассматриваться как одна из возможных стратегий, посредством которых животные реагируют на сезонные или нерегулярные изменения условий своего существования, оказывающие влияние на их жизненное пространство, размножение и потребности в пище. Кроме того, миграции обеспечивают широкий обмен особями между разными популяциями, устраняют близкородственное спаривание, предотвращают перенаселение, а также придают необходимую гибкость в распределении особей по угодьям. В то же время процесс эволюции приводит к многочисленным компромиссам и миграция наряду с преимуществами имеет и оборотную сторону. В частности, мигрирующие животные, находясь вдали от сравнительно безопасных мест, где они проводят большую часть года, оказываются более уязвимыми для своих естественных врагов и неблагоприятных абиотических факторов.
Однако даже масштабные выселения, сопровождающиеся массовой гибелью, скорее полезны для вида, чем вредны. Они не только изреживают достигшие высокой плотности популяции, но и в некоторых случаях приводят к расширению ареала. Любой вид может закрепиться на новом месте и в новых условиях, если имеется достаточно свободного пространства или в экосистеме существует незанятая экологическая ниша, либо если он обладает конкурентным преимуществом над иным, ранее закрепившимся здесь видом и в состоянии вытеснить его. Так, с эмиграцией обыкновенной белки (Sciurus vulgaris) связано заселение ею в 1920-х гг. Камчатки, где этот вид ранее отсутствовал. Принявшее в середине ХХ века широкие масштабы эмиграционное движение лося (Alces alces) и кабана (Sus scrofa) привело к быстрому восстановлению их населения на территориях, где они были истреблены 150-250 лет назад.
вперед
в оглавление