в оглавление
назад
Опыт и научение
Личный, или индивидуальный, опыт можно определить как результат встраивания приобретенных форм поведения в генетически детерминированные стереотипы. Личный опыт следует считать адаптацией животного к конкретным условиям жизни. В случае, если такая индивидуальная адаптация сопряжена с большими биологическими преимуществами, она распространяется и на другие особи, а в конечном счете захватывает целые поколения. Со временем такая изначально индивидуальная адаптация превращается в видотипичное свойство, закрепленное наследственностью.
Согласившись с концепцией двухфазной структуры инстинктивного поведения, мы не можем не касаться проблемы соотношения инстинкта и приобретенных адаптаций в поисковой и завершающей фазе инстинкта. Исследователи солидарны в том, что встраивание индивидуально изменчивых компонентов возможно в обеих фазах инстинкта. Очевидно, что поисковая фаза за счет новых поведенческих адаптаций существенно расширяет биологические возможности животного. По существу, при этом изначально индифферентные признаки окружающего мира превращаются в особо значимые сигнальные раздражители. Изменения, возникающие в поисковой фазе, приводят и к изменениям поведения в эффекторной фазе. Эффекторные изменения представляют собой перекомбинацию врожденных двигательных актов. Вновь приобретенные движения, в свою очередь, стимулируют развитие психических процессов.
Отмеченные изменения поведения в онтогенезе отражают сущность научения. Научение представляет собой сложный и многокомпонентный процесс, центральное место в котором занимает условный рефлекс. Но условный рефлекс — это не единственный путь расширения личного и, следовательно, филогенетического опыта. Научение — это совокупность изменений в поведении животного, возникающих в процессе онтогенеза.
Все способы научения можно разделить на две группы: облигатное и факультативное. Облигатное научение включает поведенческие адаптации, приобретение которых необходимо всем представителям данного вида независимо от того, в каких условиях они существуют. Факультативное научение включает в себя адаптации, необходимые для выживания конкретной особи или популяции, оказавшейся в конкретной (возможно, редкой) обстановке, т. е. факультативное научение не обязательно для всех представителей вида. Однако оно видотипично. Следует подчеркнуть, что факультативное научение характеризуется большей гибкостью и лабильностью. Тем не менее животное нельзя научить «чему угодно». Например, нельзя научить собаку вилять хвостом, или выработать условный рефлекс слюноотделения у взрослой кошки.
Научение возникает как мера облегчения или расширения спектра способов достижения цели в эффекторной фазе. Иначе говоря, существуют видотипичные генетически фиксированные рамки научения. Рамки способностей животного к научению, хотя потенциально и ограничивают животное, но способствуют удержанию видовой стабильности, ограничивая видовую изменчивость. К тому же у многих животных эти ограничительные рамки научения шире реальной необходимости. И в этом заложен определенный биологический смысл. В случае экстремальных ситуаций (катастроф, катаклизмов) эти расширенные возможности к научению (адаптации) будут востребованы. Выжившие особи станут основой для возрождения популяции и всего вида.
Способность к научению ограничивается уровнем эволюционного развития животного. Это справедливо в целом для всего царства животных. Все высшие животные имеют больше шансов на выживание. Низшие животные имеют узкие рамки способностей к научению. Однако в пределах одного класса отдельные виды проявляют нетипичные для своего класса способности к научению. Среди млекопитающих признанными эрудитами и прилежными учениками справедливо считаются китообразные и приматы. Повышенная способность к научению обеспечивает им адаптационные преимущества и биологическую экономичность. Человекообразные обезьяны и дельфины имеют лучшую приспособляемость к среде обитания не за счет многоплодия (как это имеет место у грызунов), а благодаря способности к факультативному научению в широком диапазоне условий на протяжении всего онтогенеза.
Способность к научению отражает уровень психических процессов. Чем лабильнее психика животного, тем пластичнее будет поисковая фаза и оптимальнее эффекторная фаза сложного поведенческого акта. Причина (усложнение поведения) и следствие (психика, способность к научению) в процессе эволюции постоянно меняются местами. Поэтому поведение и психика животных эволюционируют как единое целое. Эволюция совершенствует программу поведения и рамки реализации этой программы за счет научения, индивидуального опыта.
В самой основе научения лежат следующие элементарные процессы: привыкание, сенситизация, тренировка, суммация сенсорики. Эти явления обеспечивают адаптации, относящиеся к неассоциативному научению.
Привыкание (габитуация) — постепенное угасание реакций на малозначимые стимулы при их многократном предъявлении или длительной экспозиции раздражителя. Этот вид научения является наиболее распространенным способом приобретения личного опыта. Он используется животными всех систематических групп. Тем не менее надо подчеркнуть, что данный способ научения имеет первостепенное значение прежде всего для животных с несложно организованной нервной системой (кишечнополостные, плоские черви), а также для высших животных на ранних этапах онтогенеза. Привыкание рассматривается как универсальный способ «не замечать» огромное количество факторов среды, имеющих второстепенное значение. Например, кошки спят при включенном радио или телевизоре и не реагируют на речь или музыку, но моментально просыпаются при писке мышей. Для привыкания к звуку телевизора котенку требуется не более 5-7 включений. А привыкание взрослой деревенской кошки, выкармливающей котят, к вокализациям мышей, вероятно, вообще невозможно выработать.
Меньшее количество повторений для формирования привыкания предполагает наличие более высокоорганизованной психики. Так, коровы в коровнике быстро привыкают к шуму и движению трактора, раздающего корма, но в то же время пугаются при появлении в коровнике постороннего человека. Привыкание у животных с примитивно устроенной нервной системой требует на порядок больше предъявлений стимула.
Снижение реактивности нервной системы возможно и по другим причинам, отличным от явления габитуации. Это прежде всего относится к сенсорной адаптации. Многие органы чувств перестают отвечать на действие раздражителя при частых и многократных повторениях. Однако сенсорная адаптация развивается у животных с хорошо развитыми анализаторами. Кроме того, дополнительно следует отметить кратковременный характер сенсорной адаптации. У животных с высокоорганизованной нервной системой, как правило, сенсорная адаптация и габитуация совпадают во времени, что создает большие трудности при изучении явления привыкания.
Сенситизация — свойство нервной системы усиливать ответ на слабый стимул, если он сочетается во времени с другим неприятным воздействием. Так, если действие слабого раздражителя (например, легкое прикосновение, на которое животное не отвечает) сочетать с ударом электрического тока, то вскоре только на прикосновение животное будет отвечать бурной реакцией. У животных с высокоорганизованной психикой сенситизацию нелегко отличить от ассоциативного научения. В последнем случае возникает временная связь между центрами двух рефлексов. В результате второй условный раздражитель запускает реакцию, обычно проявляющуюся в ответ на предъявление первого раздражителя.
Тренировка представляет собой многократно повторяющиеся стереотипические действия приводят к оптимизации эффекторных реакций как с точки зрения затрат как времени на их выполнение, так и энергии. Особую значимость тренировка как способ неассоциативного научения имеет в раннем пренатальном и постнатальном онтогенезе.
Новорожденный жеребенок (верблюжонок, теленок, олененок) многократно повторяет попытки встать на ноги, тренируя при этом конечности и локомоторные центры, управляющие движением конечностей. В результате через час-два новорожденный уверенно следует за матерью. Сложные локомоции молодняк тренирует в процессе игр. Так формируются охотничьи навыки у котят и щенков, оптимизируется техника вскрытия ореха у молодых белок. Особое значение отводится тренировкам в процессе формирования поведенческих проявлений, сопряженных с опасностью для жизни. Именно благодаря тренировкам котята, хорьки, молодые ласки научаются наносить молниеносный смертельный укол в основание черепа животным, которые по своим физическим возможностям превосходят охотника.
Следует иметь в виду, что в процессе тренировки происходит морфофункциональное становление как эффекторов, так и нервных структур, обслуживающих данный поведенческий (тренируемый) акт. Под влиянием проприорецепторной афферентации происходит активизация локомоторных центров спинного мозга, возрастает спонтанная электрическая активность нейронов. По системе восходящих путей локомоторные центры спинного мозга активизируют ядра и ретикулярную структуру ствола, мозжечок, базальные ганглии, а также способствуют развитию двигательной коры. В нервных структурах, регулирующих работу опорно-двигательного аппарата, происходят и морфологические изменения: формирование новых синаптических связей, образование шипиков на дендритах.
Справедливости ради надо отметить, что многие сложные формы поведения не нуждаются в тренировках, например, осенняя подготовка берлоги молодняком бурого медведя и строительная деятельность бобров.
Суммация сенсорики заключается в следующем. Рецепторный аппарат животного организма отслеживает происходящие в окружающей среде изменения по целому ряду параметров физико-химической природы. Рецепторы, представляющие периферические элементы сенсорных систем, обеспечивают контроль за изменением стимулов различной модальности (свет, звук, химический состав среды, электромагнитное поле, механические воздействия). Перцептивная психика по своему определению предполагает суммацию сенсорной информации при формировании целостного образа.
Суммация сенсорики приводит к формированию доминанты. При этом центральные нейроны приобретают состояние высокой возбудимости под влиянием нескольких последовательно или одновременно действующих стимулов, каждый из которых в отдельности не вызывает адаптивных реакций. Суммация может быть следствием и предварительно активированных гуморальными агентами нейронов.
С точки зрения зоопсихологии важно отметить особые возможности двигательной активности передних конечностей у позвоночных. Имеется в виду не опорная, а именно двигательная активность передних конечностей. Их эволюционное развитие через манипулирование привело, в конце концов, к орудийной деятельности приматов и трудовой деятельности древних людей. Разнообразные дополнительные функции характерны и для ротового аппарата позвоночных. Между передними конечностями и ротовым аппаратом у животных устанавливаются сложные функциональные связи, обогащающие их поведение и психику. Например, у лисиц (Vulpes) насчитывается по крайней мере 45 дополнительных функций передних конечностей и ротового аппарата. У барсуков (Meles) их уже 50, у енотов (Procyon) 80, у низших обезьян — 150.
Способность удерживать предмет перед собой на весу позволяет животному комплексно и наиболее полно его обследовать. При этом используется оптическая, химическая, тактильная, звуковая и гравитационная сенсорика. Чем более развита способность к фиксации предмета передними конечностями и ротовым аппаратом (манипулирование), тем сложнее поведение животного. Так, медведи (Ursus) обладает всего тремя способами фиксации предмета на весу, еноты — шестью, низшие обезьяны (макаки (Macaca), павианы (Papio)) — более чем тридцатью способами фиксации. Причем обезьяны располагают высококоординированной функцией деструктивного анализа фиксируемого объекта (расчленение на отдельные компоненты). Еще более совершенна эта способность у человекообразных обезьян и человека. Они способны к фиксации и расчленению предмета одной конечностью (рукой). Расчленение предмета здесь рассматривается как разновидность исследовательского поведения с целью получения информации о его внутреннем строении.
Таким образом, прогрессивное развитие дополнительных двигательных (не опорных) функций передних конечностей животных, их приспособленность к процессу манипулирования предметами служит мощным фактором психического развития животных.
Комфортное поведение (интравертное) является разновидностью манипулирования. Особенностью комфортного поведения выступает то, что в этом случае объектом манипулирования выступает собственное тело животного. К этому виду поведенческой активности относят и движения тела без пространственной направленности (смена ног при стоянии, переворачивание животного с одного бока на противоположный при лежании).
Комфортное поведение для удобства изучения может быть подразделено на несколько видов: очищение тела, потряхивание, почесывание о предмет, катание по субстрату (пыль, зола, песок, грязь), купание (в воде, золе, пыли). Комфортное поведение обеспечивает решение одной задачи — устранение состояния дискомфорта, которое развивается на основе антисанитарного состояния тела (пот, грязь, паразиты, мелкие травмы), а также в результате однообразного положения тела в пространстве и возникающей неравномерной нагрузке отдельных частей тела под влиянием сил гравитации. Кроме того, дискомфорт может развиться вследствие социальной активности животных, а также под влиянием неблагоприятных погодных явлений (дождь, град, снег, ветер).
В суточном балансе поведения млекопитающих комфортное поведение занимает большой удельный вес. Скажем, лошади в состоянии покоя переминаются с ноги на ногу, меняют положение в состоянии лежа. Они же могут почесываться о столб или перекладину ограждения, валяться на земле при наличии зуда в области спины.
Кошки известны как чистоплотные животные, прежде всего благодаря тому, что очень много времени уделяют грумингу. В течение суток они многократно подвергают вылизыванию собственное тело. При этом животные пользуются не только языком, но и передними конечностями, которые предварительно обильно смачивают слюной. Кошки — одни из немногих животных, которые в состоянии обработать языком и лапами все части собственного тела, включая такие труднодоступные участки, как спина, затылок, хвост и даже ушные раковины.

Кошка (Felis silvestris catus) вылизывается.
Комфортное поведение инстинктивно, что доказывается его видоспецифичностью, т. е. генетической запрограммированностью. При изучении поведения зубра (Bison bonasus), бизона (Bison bison) и их гибридов у животных обнаружили 107 видотипичных поз. Но интересны следующие наблюдения. Только бизон в положении стоя способен вылизать весь хвост от кончика до основания. Зубр до корня хвоста не дотягивается. В то же время лишь зубр может в положении стоя вылизать полностью свое бедро. Бизон этого не делает. Интересно, что у телят зубра и бизона видовых различий комфортного поведения очень мало. Видовые различия груминга у них формируются на поздних стадиях онтогенеза. С другой стороны, у гибридов зубра и бизона можно наблюдать признаки комфортного поведения обоих видов. Другими словами, арсенал поз и действий комфортного поведения гибридов богаче по сравнению с исходными формами.
Перцептивная психика базируется на хорошо развитых анализаторах. Характерной особенностью всех представителей животного мира является активное использование химического анализатора во взаимоотношениях индивидуума с внешней средой. Химический анализатор является самым древним и включает обонятельную и вкусовую. Органы химической рецепции хорошо развиты у всех животных независимо от их эволюционной организации и среды обитания. Однако высокая химическая чувствительность животных больше связана с сенсорной психикой.
Для образного восприятия окружающего мира нужны дополнительные системы взаимодействия с внешней средой. Необходимым условием сложной психики является наличие у животного хорошо развитого зрения. Зрительный анализатор обеспечивает предметное восприятие, включая восприятие формы. Все позвоночные животные способны именно к такому восприятию внешнего мира благодаря зрению. Эволюционная потеря зрения приводит к обеднению психики животного.
Лучше всего перцептивная психика развита у млекопитающих. Это ясно подтверждают опыты с отсроченными стимулами. Животному позволяют отыскивать спрятанный корм спустя некоторое время после его предъявления. С такой задачей легко справляются такие животные, как обезьяны, собаки, кошки, свиньи. Птицам и тем более рыбам такие задачи не по силам. Успешное решение задачи производится на отсутствующий стимул, что возможно лишь при наличии у животного чувственных представлений.
Экспериментально доказано, что высшие позвоночные животные реагируют на корм, а низшие — на кормушку с кормом. В опытах с отсроченными стимулами с использованием обезьян в качестве объекта наблюдений у животных на виду прятали банан, который впоследствии незаметно заменяли на капусту. Понятно, что банан и капуста имеют разный уровень привлекательности для обезьяны. Обезьяна быстро находила корм, но, обнаружив на месте захоронения не вкусный банан, а капусту, бурно протестовала визгом, гримасами, а затем продолжала поиск исчезнувшего банана. Подобное поведение характерно и для собак, свиней, дельфинов. Это примеры когнитивного (познавательного) поведения. В приведенных примерах для успешного решения задачи от животных требуется мысленное представление (образ) спрятанного предмета.
К разряду когнитивного поведения можно отнести и поведение животных в лабиринте, где быстрое решение поставленной задачи методом проб и ошибок невозможно. Наличие у крысы пространственного «плана» лабиринта позволяет ей решать задачи, поставленные экспериментатором, очень быстро, без лишних действий — посещения заведомо пустых коридоров лабиринта.
Примером когнитивного поведения можно считать и работу служебных собак — поиск преступника по следу и его задержание, выборку вещи, поиск наркотиков и оружия. Во всех этих случаях собака отыскивает объект (вещество) с определенными свойствами, т. е. собаке задается некий образ, который она и держит в голове. При следовой работе собака не бросается на первого попавшегося на ее пути человека. Так же и при выборке животные отыскивают и выбирают из множества предметов только тот, образ которого им задан. Обоняние у собаки служит основой для поискового поведения. Но на завершающем этапе решение задачи происходит на базе афферентного потока, поступающего от зрительного анализатора. Скажем, если собака вышла по следу на источник заданного ей запаха, но вместо человека обнаружила укрытие или автомобиль, она испытывает разочарование, подобно той обезьяне с исчезнувшим бананом.

Собака (Canis lupus familiaris) идет по следу.
В основе когнитивного научения зачастую лежат условные рефлексы второго и более высокого порядка. В раннем онтогенезе позвоночных животных велика роль более простых форм научения, таких как импринтинг, подражание, упражнение.
Импринтинг (запечатление) выгоден прежде всего зрелорождаемым видам животных. Он гарантирует быстрое и своевременное узнавание матери и представителей своего вида. Импринтинг оказывает большое влияние на формирование личного опыта в ранний постнатальный период.
Подражание тесно связано с импринтингом и реакцией следования. Научение путем подражания часто называют имитационным научением, имея в виду, что индивидуальное формирование новых форм поведения у животного происходит в результате непосредственного восприятия и копирования действий других особей.
Как всякое научение, подражание может быть облигатным (видоспецифичным) и факультативным. Первое обеспечивает молодежь репертуарными действиями своего вида. Этот способ научения можно наблюдать у стадных животных. Приобретаемые путем подражания навыки позволяют молодняку распознавать представителей своего вида, отыскивать пищевые объекты, адекватно реагировать на появление опасности. Например, ягнята и козлята первых дней жизни подражают матери на пастбище — совершают захват ртом травы. Однако траву в это время они еще не едят. Но совершая такие действия, они научаются отыскивать кормовые виды растений и определять их съедобность. Факультативность данного поведения ягнят подтверждают эксперименты. Ягнята и козлята, выращенные до 5-месячного возраста на молочно-концентратной диете изолированно от взрослых животных, на пастбище первое время не проявляли интереса к траве. Однако при выпасе вместе со взрослыми животными через некоторое время они начинали им подражать — хватать траву, но не есть ее. Спустя какое-то время козлята и ягнята научились питаться на пастбище.
Подражание материнскому поведению у козленка горала (Naemorhedus caudatus).
Подражание у хищных млекопитающих формирует охотничье поведение. Так, котята учатся выслеживать жертву и умерщвлять ее. Многократные повторения этих действий в коллективных играх котят превращают их в искусных охотников. Многие животные путем подражания приобретают очень сложные поведенческие формы. Так, некоторые обезьяны через подражание научаются строить гнезда.
Факультативное научение через подражание описано у кошек, собак, обезьян. Котята и щенки подражают родителям при встрече с незнакомцем. Котята при этом могут шипеть, U-образно изгибать спину, надувать трубой хвост. Щенки, подражая старшим, рычат и лают. Щенки мужского пола, глядя на взрослых кобелей, пытаются помочиться «по-взрослому», поднимая заднюю ногу. Обезьяны, вероятно, обладают наиболее развитыми способностями к факультативному научению через подражание. Именно этим можно объяснить их склонность водружать на нос очки, надевать на голову шляпу или брать в рот сигареты. Исследователи сообщают, что шимпанзе, подражая человеку, пытаются «читать» газеты и книги. Последние примеры научения через подражание имеют сугубо факультативное назначение.
Таким образом, подражание позволяет животным существенно обогатить свой личный опыт. Путем подражания не могут формироваться лишь инструментальные навыки животных. Однако этот тезис относится хоть и к абсолютному большинству животных, но все-таки не ко всем. В случае наличия у животного высокоразвитой перцептивной психики возможно и инструментальное научение животных через подражание. Именно такие случаи описаны исследователями, работающими с приматами. Так, некоторые обезьяны научаются открывать клетку ключом, подражая при этом человеку. Молодые обезьяны путем подражания старшим научаются доставать пищу при помощи других предметов (палки).
Ряд поведенческих навыков животных как облигатного, так и факультативного характера формируются под влиянием многократно повторяющихся действий, т. е. упражнения, или тренировки, которую можно рассматривать как врожденный метод приобретения личного опыта. Приведем несколько примеров.
Бельчата, выращенные до 66-дневного возраста в неволе и не видевшие орехов, вначале прокусывают орехи бессистемно, т. е. каждый раз в разных местах ореха. Через определенное количество попыток вскрытие ореха оптимизируется: все белки прогрызают скорлупу в строго определенном месте ореха, чем облегчают извлечение ядрышка.
Надо подчеркнуть, что упражнение как метод научения широко представлен в играх молодняка. Он используется для отработки сложных локомоций, связанных с добыванием пищи, защитой от нападения, половыми рефлексами. Естественно, что сложность тренируемых локомоций возрастает по мере развития молодняка.
Ласка (Mustela nivalis) убивает свою жертву (мышь, крысу) всегда одинаково. Она наносит молниеносный укол острым и относительно длинным клыком в основание черепа своей жертвы, чем вызывает временный паралич животного. Ласка, выращенная в неволе и изоляции от соплеменников и не имевшая возможности охотиться на грызунов, при первой встрече с жертвой убивает ее неумело. Она наносит многочисленные укусы в самые разные части тела, что небезопасно для маленького хищника в случае нападения, скажем, на крупную крысу. Однако через какое-то количество повторений ласка начинает убивать свою жертву стереотипично — как все — при помощи молниеносного укола клыком в основание черепа.
Очевидно, что упражнения видотипичных локомоций доводят их до совершенства только в случае, когда тренировки происходят на определенном (раннем) этапе онтогенеза. Таким образом, можно заключить, что даже многие врожденные поведенческие стереотипы нуждаются в своевременных упражнениях (тренировках).
Все формы ассоциативного научения предполагают образование условного рефлекса. Поскольку условные рефлексы возникают в определенных условиях и, строго говоря, не обязательны для всех представителей вида, то они определяют индивидуальность поведения животных. Безусловные рефлексы и таксисы обеспечивают жесткую связь животного со средой, в которой оно живет. Условные рефлексы создают пластичную основу для взаимоотношений индивидуума со средой обитания.
Существует несколько схем классификации условных рефлексов.
Классический условный рефлекс вырабатывается при предъявлении животному условного раздражителя с последующим предъявлением подкрепляющего раздражителя. Так, например, в классическом павловском слюноотделительном рефлексе включение звонка подкрепляется дачей собаке пищи. На этапе выработки условного рефлекса подкрепление условного стимула натуральным безусловным раздражителем обязательно и не связано с тем, будет ожидаемая реакция (слюноотделение) или нет. После нескольких повторений опыта у собаки усиленное слюноотделение происходит в ответ на предъявление условного раздражителя (звонок) без подкрепления пищей.
В приведенном примере классический рефлекс имеет эффекторную фазу в форме вегетативной реакции — слюноотделения. Однако это необязательное свойство классического условного рефлекса. Он может иметь и другие поведенческие проявления — локомоции, сосудодвигательные реакции. Если периодически условный раздражитель не подкреплять безусловным, происходит угасание условного рефлекса, поскольку условный раздражитель утрачивает свое первоначальное значение. Это явление известно и как угасательное торможение. Угасательное торможение защищает организм от бесполезных адаптаций и высвобождает нервные структуры для того, чтобы обеспечивать адекватные реакции животного организма на изменения, происходящие во внешней среде.
Инструментальный условный рефлекс предполагает обязательное выполнение животными определенных движений с участием опорно-двигательного аппарата (как правило, конечностей или головы). Только после выполнения той или иной двигательной реакции животное получает вознаграждение в форме положительного подкрепления (или избавления от действия отрицательного стимула). Таким образом, инструментальный условный рефлекс характеризуется активным поведением животного на предъявление условного раздражителя.
Фактически, инструментальный рефлекс — это частный случай классического условного рефлекса. В инструментальном условном рефлексе животное получает подкрепление (пищу или избавление от неволи) только после выполнения определенного двигательного акта. При этом время рефлекса от опыта к опыту сокращается. Первое решение задачи является результатом случайного поиска (метод проб и ошибок): кошка случайно нажимает на рычаг и открывает дверцу клетки. Во второй раз животное затрачивает меньше времени на то, чтобы открыть дверцу. А после некоторого количества опытов кошка затрачивает минимум времени и движений для того, чтобы выбраться из клетки.
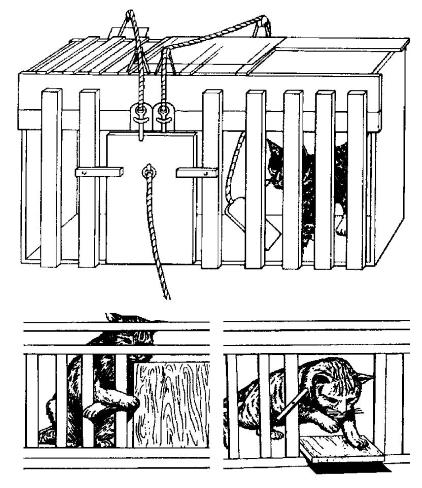
Схема выработки инструментального условного рефлекса.
Условные рефлексы можно подразделять по характеру ответной реакции на двигательные, сосудодвигательные и секреторные.
По биологическому значению условные рефлексы классифицируют на пищевые, половые, оборонительные. В этом случае в их основе будут лежать соответствующие врожденные рефлексы.
Еще один принцип классификации условных рефлексов базируется на том, какой модальности условный раздражитель и какая сенсорная система используются при их формировании. В связи с этим можно говорить о зрительных, обонятельных, слуховых, тактильных, термических условных рефлексах.
Наконец, условные рефлексы классифицируют и по тому, на какие рецепторы — экстерорецепторы, интерорецепторы или проприорецепторы — воздействует условный раздражитель при выработке условного рефлекса. Соответственно такие условные рефлексы будут называться экстерорецептивными (например, рефлексы на различные звуки, запахи), интерорецептивными (например, рефлексы, вызванные раздражением слизистой желудка, влагалища, прямой кишки) или проприорецептивными условными рефлексами (например, при раздражении мышечных веретен или рецепторов суставов конечностей).
Экстерорецептивные рефлексы обслуживают процесс взаимодействия животного со средой обитания. Зачастую от них зависит выживание индивидуума. Поэтому они формируются за более короткий промежуток времени и при меньшем количестве безусловных подкреплений. Так, большинство экстерорецептивных двигательных условных рефлексов собак устанавливаются после 5-20 подкреплений, а на выработку интерорецептивного условного рефлекса у той же собаки потребуется от 50 до 150 безусловных подкреплений, т. е. на порядок больше. Интерорецептивные условные рефлексы связаны с поддержанием постоянства внутренней среды (гомеостаза). Они не требуют и не могут обеспечить высокой скорости образования временных связей, поскольку в их эффекторной реализации задействуются многие звенья, включая медленные (на фоне локомоций) биохимические реакции.
Образование условного рефлекса проходит несколько стадий. В самом начале при сочетанном предъявлении условного и безусловного стимулов прежде всего развивается ориентировочная реакция. На этой стадии отмечается повышение электрической активности нейронов головного мозга. Особенно выражен этот процесс в проекционных зонах коры двух сочетаемых раздражителей. Ориентировочная реакция предельно обостряется при наличии доминанты (пищевой, половой, материнской и др.). Доминанта повышает реактивность нервной системы на раздражители различной модальности.
После первых сочетаний условного и безусловного раздражителей в центральной нервной системе возникает явление генерализации, т. е. некоторое время все индифферентные стимулы воспринимаются равно условными. Лишь при многократных сочетаниях одного и того же условного раздражителя с безусловным натуральным раздражителем происходит его избирательная рецепция как биологически более значимого фактора среды. Эта стадия известна как специализация.
Центральным элементом в механизме выработки условного рефлекса является образование временных связей между отдельными функционально обособленными группами нейронов. Ранние исследования говорили о том, что временные связи образуются в коре больших полушарий между группой нейронов, воспринимающих действие условного раздражителя и корковым представительством центра безусловного рефлекса. Безусловный раздражитель, имеющий нервный центр в подкорковых образованиях головного мозга, обязательно имеет свое представительство еще и в коре больших полушарий.
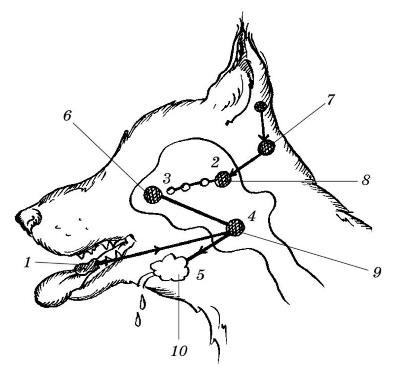
Схема образования временной связи в коре больших полушарий: 2-3 — временная связь в коре между представительствами слухового рецептора и центра слюноотделения; 1-4-5 — дуга безусловного рефлекса слюноотделения; 2-3-4-5 — дуга образовавшегося условного рефлекса слюноотделения; 6 — корковый рецептор слюноотделения; 7 — слуховые рецепторы; 8 — слуховая проекционная зона; 9 — центр слюноотделения; 10 — слюнная железа.
Более поздние исследования показали, что выработка условных рефлексов возможна и у декортицированных животных (собак, кошек). У низших позвоночных кора не развита и временные связи образуются в других (подкорковых) структурах головного мозга. Так, у птиц и рептилий к механизму установления временных связей имеет отношение полосатое тело. Образование временных связей при выработке условных рефлексов у представителей класса рыб происходит в мозжечке и среднем мозге. Однако даже у низших позвоночных под термином «временная связь» скрывается сложное многоуровневое явление. Фактически при выработке условного рефлекса временная связь устанавливается между корковыми (или мозжечковыми, диэнцефальными высшими структурами) представительствами двух безусловных рефлексов. Условный рефлекс представляет собой результат синтеза двух и более безусловных рефлексов.
Таким образом, рефлекторная дуга безусловного рефлекса представляет собой многоуровневое образование, где нервные элементы коры составляют центральную часть этой дуги. Однако, помимо коры, она включает структуры спинного мозга, продолговатого мозга, ствола, среднего мозга, а также подкорковых образований. Следовательно, и корковая временная связь условного рефлекса — это лишь часть морфофункциональных изменений, возникающих в центральной нервной системе в процессе выработки условного рефлекса. Временные связи возникают и между отдельными центрами подкорковых структур. Поэтому образование условных рефлексов происходит и у собак, лишенных коры. Правда, такие рефлексы достаточно просты. Это условные рефлексы на свет или на звук. Инструментальные условные рефлексы, требующие активизации двигательной коры, выработать у животных с поврежденной корой невозможно.
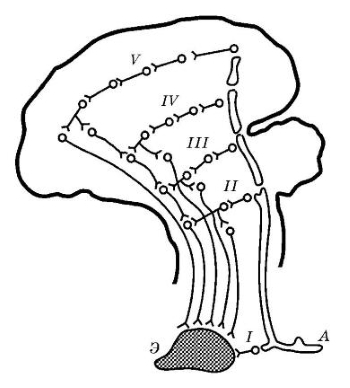
Схема, иллюстрирующая многоуровневый характер дуги рефлекса: Э — эфферентная клетка; А — афферентная клетка; I-II-III-IV-V — центральная многоуровневая часть дуги; V — корковое звено дуги (проекция) безусловного рефлекса.