в оглавление
назад
Опыт и научение (продолжение)
Научение и расширение личного опыта становится возможным благодаря уникальному свойству нервной системы — памяти. Нейроны мозга, имеющие отношение к научению, обладают свойством определенным образом изменяться под влиянием поступающей афферентации. Эти изменения относятся к электрическому состоянию клеток, биохимическим процессам и даже к морфологии нейронов, т. е. в результате процесса научения животного афферентный поток изменяет морфофункциональное состояние нейронов мозга. В конечном счете эти изменения предопределяют усвоение, хранение и возможность воспроизведения информации. Таким образом, процесс научения и явление памяти имеют единую морфофизиологическую основу.
Память — это способность животных воспринимать, сохранять и воспроизводить информацию о ранее воздействовавших на них стимулах. Во временном формате память подразделяют на кратковременную, промежуточную (лабильную) и долговременную.
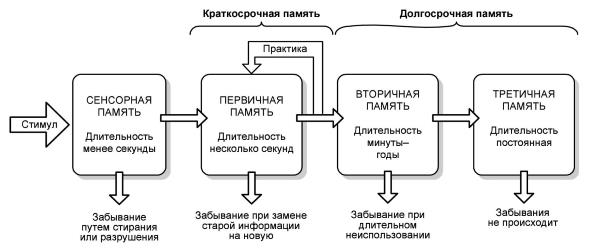
Виды и формы памяти.
Некоторые исследователи на этапе, предваряющем кратковременную память, дополнительно выделяют сенсорную, или сверхкороткую, память. При этом имеют в виду след возбуждения, который остается в сенсорной системе. Продолжительность существования этого следа всего 50-500 мс. Поскольку данное явление не в полной мере отвечает классическим характеристикам явления памяти (восприятие, хранение и воспроизведение информации), то многие ученые небезосновательно предпочитают называть это явление «перцептивным или сенсорным следом».
Сенсорный след невоспроизводим. Он представляет собой начальный этап переработки информации, поступающей из внешней и внутренней среды организма. Сенсорный след предельно субъективен. Его характер зависит от индивидуальных особенностей животного (физиологического состояния, эмоционального состояния, возраста). Сенсорная память избыточна; она представляет собой комбинацию афферентных потоков от зрительного, обонятельного, слухового, тактильного и других анализаторов. На этом начальном этапе адаптаций животный организм выбирает наиболее значимую информацию и после своеобразной ее фильтрации отправляет в кратковременную память.
Центральное место в формировании сенсорной памяти занимает импринтинг, в основе которого лежит образование энграммы — следа от афферентного потока на уровне коры и подкорковых образований мозга. Причем субстратом для энграммы служат нейроны, межнейронные синапсы и глиальные клетки. В соответствии с современными представлениями, энграмма памяти есть ансамбль нейронов и глиальных структур, объединенных специфическими синаптическими механизмами. Под влиянием афферентного потока, поступающего от периферической части сенсорных систем, клетки головного мозга запускают процесс экспрессии специфических белков — коннектинов, которые встраиваются в мембрану контактирующих клеток. В этих участках и образуются новые синапсы или шипики, представляющие собой шлюзы, обеспечивающие беспрепятственное движение электрического потенциала между клетками, объединенными в реберверирующий круг. В процессе научения и формирования сенсорного следа наблюдается гиперполяризация нейронов вследствие возросшей калиевой проводимости их мембранного комплекса. Далее происходит активизация натриевого насоса, что ускоряет синаптическое продвижение нервного импульса.
Устойчивость энграмм особенно велика у животных в раннем постнатальном периоде. Известны чувствительные периоды раннего онтогенеза, когда происходит быстрое и устойчивое образование сенсорных следов, которые на системном уровне проявляются в форме импринтинга. В эти чувствительные периоды сенсорные следы от зрительного, слухового, обонятельного анализаторов очень быстро переадресовываются в блоки долговременной памяти животных.
После фильтрации афферентации в процессе сенсорной памяти отобранная информация поступает в систему кратковременной памяти, где подвергается дальнейшей сортировке и фильтрации. Этот процесс занимает, как правило, несколько минут, после чего наиболее значимая информация закладывается в систему долговременной памяти.
В ходе исследований были выявлены замкнутые цепи нейронов, по которым происходит многократное движение электрических импульсов при формировании кратковременной памяти. При этом данные нейронные цепи локализованы на уровне корково-подкорковых структур. Например, они существуют между корой и таламическими ядрами, между корой и гиппокампом. Это явление электрической циркуляции по замкнутым цепям нервных клеток получило название реверберации. Обнаружены реверберирующие цепи нейронов между правым и левым полушариями мозга. Убедительным доказательством наличия реверберирующих цепей служит и лимбический круг, включающий поясную извилину коры больших полушарий, гиппокамп, миндалину, гипоталамус.
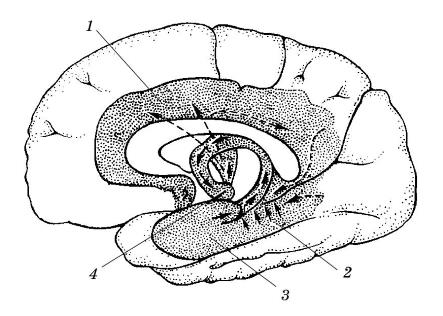
Ревербирующий лимбический круг: 1 — поясная извилина; 2 — гиппокамп; 3 — миндалина; 4 — гипоталамус. Стрелки отражают направление циркуляции потенциалов между ними.
В лобных долях больших полушарий обнаружены нейронные круги типа ловушек. Внутрикорковую реверберацию в пределах лобной коры рассматривают как морфофункциональную основу кратковременной памяти и проявления внимания и ориентировочной реакции. Реверберация между симметричными отделами коры правого и левого полушарий через мозолистое тело рассматривается как основа для системной организации кратковременной памяти.
После консолидации памяти энграмма проходит дополнительную фильтрацию, и отобранная информация оказывается в хранилищах долговременной памяти.
Кратковременная память сохраняет информационный след в течение нескольких минут. Свойства кратковременной памяти изучают методом экспериментальной ретроградной амнезии. Этот метод позволяет разобщить систему кратковременной и долговременной памяти. Ретроградная (обращенная в прошлое) амнезия (потеря памяти) наступает в результате травмы, электрошока, гипоксии или под влиянием наркотических средств. В экспериментах ретроградную амнезию моделируют электрошоком, который наносится сразу после обучения животного. Это приводит к потере или нарушению воспроизведения следов памяти. Электрошок не влияет на долговременную память. Если воздействовать электрошоком спустя час и более после обучения, то результаты научения не стираются. Это позволяет считать, что процесс перехода информации из системы кратковременной памяти в систему долговременной памяти (процесс консолидации энграммы) завершается за время, исчисляемое часами.
Поскольку не существует экспериментального (электрофизиологического, биохимического или гистологического) подтверждения того, что при ретроградной амнезии элиминируется энграмма памяти, высказывается предположение, что электрошок или травма влияют на механизм воспроизведения энграммы, а не на сам сенсорный след. В доказательство приводятся результаты опытов на человеке с подсказками (напоминанием), после которых «забытая» пациентом информация воспроизводится частично или в полном объеме. С позиции такого подхода к проблеме кратковременной памяти процесс консолидации памяти представляется как процесс формирования механизма воспроизведения ее энграммы.
Наблюдение за больными с потерей памяти, а также специальные эксперименты на животных позволяют определить круг структур головного мозга, которые имеют отношение к формированию кратковременной памяти. Научение на базе пространственной ориентации нарушается при повреждении маммилярных тел гипоталамуса. Отмечены амнестические явления на уровне кратковременной памяти у больных с повреждениями ретикулярной формации. У обезьян нарушение кратковременной памяти вызывали путем повреждения вентрального гиппокампа. Еще более глубокие нарушения кратковременной памяти связывают с функцией миндалин. Из кортикальных образований в связи с этим следует назвать поясничную извилину, имеющую связи с гиппокампом и передним таламусом, а также височные доли, медиальную часть префронтальной коры и ассоциативные области новой коры. Гиппокамп имеет особое значение для формирования энграммы памяти, поскольку осуществляет фильтрацию афферентного потока, прежде чем он достигает корковых структур. Гиппокамп исключает жесткую фиксацию сенсорных следов, имеющих случайный характер или несущих малозначимую для данного мотивационного состояния характеристику стимула.
В долговременную память поступает только информация большой значимости. Долговременная память формируется под влиянием систем подкрепления, т. е. имеет условно-рефлекторную составляющую.

Молодые западные серые кенгуру (Macropus fuliginosus), для которых схватка — это и игра и обучение одновременно, учатся драться с раннего возраста. Наблюдения показывают, что молодые самцы меняют интенсивность игровых схваток в зависимости от возраста своих противников. Когда в столкновения вовлекают молодежь, старшие особи сами уравнивают условия, проявляя меньшую агрессию, чем они это делают с более взрослым соперником. Таким образом, они стараются продлить схватку, сохраняя при этом интерес у всех участников.
Интересен вопрос о вместительности памяти. Количественную оценку кратковременной и долговременной памяти дать очень сложно. Очевидно одно — емкость системы долговременной памяти на несколько порядков выше. Психологические эксперименты на добровольцах показывают, что при необходимости быстрого запоминания (активизируются элементы кратковременной памяти) человек безошибочно воспроизводит (вспоминает) 7 ± 2 предъявленных для запоминания предметов. В других опытах добровольцам демонстрировали слайды с комментариями через небольшие временные интервалы. Спустя 2-3 дня испытуемым предъявляли по два слайда, один из которых был им показан накануне, и просили определить, какой слайд им знаком. В данном эксперименте ставилась задача выяснить возможности долговременной памяти пациентов. Оказалось, что испытуемые узнавали до 10 000 ранее увиденных слайдов. На основании полученных результатов было сделано заключение о том, что объем долговременной памяти человека на узнавание ранее увиденных объектов не ограничен.
Информация в долговременной памяти от одних и тех же стимулов внешней среды у разных индивидуумов будет иметь существенные отличия.
Искажения восприятия действительности происходят вследствие того, что сенсорный след фильтруется гиппокампом и изменяется под влиянием других структур лимбической системы, отвечающих за эмоциональную окраску потребностей животного организма. Поэтому у разных котят из одного помета формируется различное отношение, скажем, к петуху. У котенка, который испытывал по каким-то причинам страх и впервые при этом увидел петуха, впоследствии петух будет вызывать страх. Другой котенок впервые увидел петуха в процессе увлекательной игры со своими сверстниками. Последующие встречи этого котенка с петухом никакого страха у него вызывать не будут. Разница в восприятии одного и того же объекта у этих котят связана с тем, что процесс консолидации памяти при первой встрече с петухом у них проходил на разном эмоциональном фоне.
Гиппокамп осуществляет двойную функцию в процессе формирования энграммы памяти. С одной стороны, гиппокамп служит избирательным фильтрующим звеном на пути информации к корковым структурам. С другой стороны, гиппокамп участвует и в процессе селективного извлечения информации под влиянием мотивационного состояния организма.
Механизм долговременной памяти пока до конца не ясен. В связи с этим ученые предлагают несколько концепций для объяснения явления долгосрочного хранения информации.

Подобно человеческим детям, малыши японских макак (Macaca fuscata) аккуратно лепят руками маленькие снежки, катают их по снегу, чтобы сделать больше, и кидаются ими.
Теория структурных изменений. Достоверно известно, что в процессе обучения и перехода информации в долговременную память в головном мозге происходят некоторые структурные изменения. Гистологические исследования показывают, что в процессе формирования долговременной памяти в некоторых отделах мозга возрастает количество синаптических связей. Отмечается, кроме того, увеличение размеров существовавших до начала научения синапсов. Под влиянием процесса научения происходит увеличение количества шипиков в синапсах. Шипики представляют собой специализированные постсинаптические образования, облегчающие прохождение возбуждения через синаптическую щель.
Этологические исследования показывают, что у крыс, выращенных в условиях обогащенной среды, кора большого мозга имеет большую толщину по сравнению с аналогичными крысами, выращенными в условиях обедненной среды. Опыты на лабораторных мышах свидетельствуют, что у животных, выращенных в полной темноте, под влиянием последующего освещения происходит разрастание дендритов пирамидных клеток. На субклеточном уровне в процессе формирования памяти обнаруживают увеличение числа микротрубочек и рибосом в составе цитоплазмы нейронов.
Ряд ученых считают, что к процессу формирования следов долговременной памяти имеют отношение не только нейроны, но и окружающие их глиальные клетки. Нейроглия способствует повышению проводимости нервного импульса в нейронах реверберирующих цепей. Под влиянием экспериментальной электропотенциации нейронов происходит дополнительная миелинизация отростков нейронов, что повышает скорость проведения импульса. В процессе формирования долговременной памяти происходит накопление РНК в глиальных клетках.

Афалина (Tursiops truncatus) играет осьминогом перед тем, как его съесть.
Теория биохимических изменений. Установлено, что в процессе формирования долговременной памяти происходят заметные изменения белкового метаболизма в нейронах и клетках нейроглии. Из мозга выделили ряд белков, получивших название «белков памяти». Сюда относятся белки S-100, ацетилхолинэстераза, холинорецептивный белок, белок 14-3-2. Всего, судя по литературным сообщениям, насчитывается несколько десятков веществ белкового происхождения, которые каким-то образом связаны с обучением и памятью. Однако безоговорочно признается прямая причастность к формированию энграммы памяти лишь для нескольких белков. Это нейронный белок S-100 и белок S-110, имеющие цитозольное происхождение, а также ряд белков синаптической локализации. Кроме того, доказано, что в процессе образования долгосрочной памяти происходит накопление белковых веществ в нейроглии (белки 10В, NS-1, NS-2, GFAP). Белок S-100 накапливается в гиппокампе при переучивании крыс.
Процесс формирования долговременной памяти сопровождается усилением синтеза РНК в нервных клетках. Если процесс синтеза РНК блокировать (например, введением Д-актиномицина или пуромицина), то затрудняется процесс научения животных. Скорость транскрипции РНК с молекулы ДНК особенно высока в тех структурах мозга, которые связаны с научением и памятью. Выделенная из лобных долей РНК отличается уникальной способностью к гибридизации. Однако РНК сама по себе не может быть носителем энграммы памяти, поскольку она не специфична для стимулов разной модальности. Повышение синтеза РНК при формировании долговременной памяти следует рассматривать как следствие усиления белкового анаболизма вообще и синтеза белков памяти в частности.
Критики теории белков памяти в качестве сильного контраргумента ссылаются на то, что период жизни белка не может превышать нескольких месяцев (чаще период их жизни значительно короче). При этом они указывают, что 90% белков головного мозга обновляются в течение 2 недель.

Койоты (Canis latrans) — территориальные животные, поддерживающие строгую иерархию внутри своих стай. В то же время с самого рождения они охотно играют друг с другом, принимая участие в безвредных схватках. Приглашая друг друга к игре, койоты используют едва различимые сигналы, показывающие, что их намерения не агрессивны. Сама игра содержит много элементов агрессии (укусы, быстрые движения головой из стороны в сторону и т. д). При этом, совершая регулярные «поклоны» (задние ноги стоят, а голова и передние ноги прижаты к земле), койот демонстрирует, что он только выглядит грубым и резким, хотя на самом деле хочет играть. Для продолжения совместной игры с более слабым или молодым партнером сильный койот будет воздерживаться от сильных укусов и бегать не столь быстро, как может на самом деле.
С памятью связаны и многие нейрогормоны. Например, окситоцин затормаживает образование следов памяти. Эндогенные опиаты, такие как эндорфины и энкефалины, также препятствуют процессу научения. Однако они затормаживают процесс угашения условных связей. Известно, что на консолидацию энграмм положительно влияет вазопрессин. Процессу формирования памяти способствует глютаминовая кислота и у-аминомасляная кислота.
Весьма интересными для понимания механизма памяти были экспериментальные попытки переноса памяти от обученных животных к их необученным соплеменникам. В ходе одного эксперимента у лабораторных крыс вырабатывали страх перед темнотой, а затем выделяли из их мозга особый пептид, получивший название скотофобина. При введении этого пептида в организм непуганых крыс последние демонстрировали боязнь темноты. Подобным образом у крыс стимулировали образование нейропептида, названного амелитином. Этот пептид в нарастающем количестве накапливался в мозге крыс, которых приучали не бояться звука резкого звонка. Внутрибрюшинное введение этого пептида делало контрольных крыс менее пугливыми по отношению к звонку.
Большинство исследователей памяти не находят биохимические исследования достаточно убедительными, чтобы признавать идею молекулярного кодирования следов памяти. Все же было показано, что в процессе обучения морфофункциональное состояние синапсов клеток, участвующих в реверберирующей импульсной рециркуляции, подвержено существенным изменениям и что эти изменения сохраняются достаточно длительное время. Также удалось доказать, что в процессе формирования кратковременной памяти происходят структурные изменения каналов цитоплазматического ретикулума, которые облегчают выход ионизированного кальция. Это влечет за собой увеличение выброса нейромедиатора и, следовательно, облегчение прохождения возбуждения через данный синапс.

Молодые койоты (Canis latrans) учатся выть, подражая матери. Хотя койоты живут небольшими стаями, но пищу обычно ищут поодиночке. Члены стаи периодически стараются поддерживать звуковой контакт. Голосовые сигналы почти всегда включают вой и некоторое количество высокочастотного лая. Вой состоит из серий высокочастотных взвизгиваний, за которыми следует длинный, как сирена, вой, обычно звучащий на рассвете, в вечерних сумерках или ночью. Длинные, чистого тона завывания слышны дальше и передают более сложную информацию, тогда как короткий лай необходим, чтобы оценить расстояние между особями. Сочетание двух сигналов позволяет койотам координировать свои перемещения и согласовывать действия при защите участка. Последнее очень важно, поскольку вторжение других койотов может привести к гибели детенышей размножающейся пары. Голоса горных койотов сильнее, чем у их равнинных собратьев, а их тон обычно ниже.
Синаптическая передача имеет отношение и к долговременной памяти. При ее формировании усиливается белковый обмен в нейроне, что приводит к резкому нарастанию концентрации специфических пептидов в структуре синапса. Внешне (при помощи микроскопа) исследователи наблюдали своеобразное «разбухание» синапса вследствие повышения его функциональной активности и концентрации в нем протеинов.
В научном сообществе большой популярностью в силу своей широкой доказательной базы пользуется гипотеза многонейронной системы хранения следов долговременной памяти. Она не противоречит предыдущей теории. Молекулярные трансформации синапса в процессе научения объясняют механизм стойкой реверберации потенциалов в нейронных цепях.
Таким образом, процесс формирования следа долговременной памяти и ее хранение все еще остаются загадочными явлениями. Однако очевидно, что в основе как кратковременной, так и долговременной памяти лежат пластические процессы нервных клеток и их синаптических связей, обеспечивающих образование межклеточных комплексов многоуровнего формата.
Еще более непонятен в нейрофизиологическом отношении процесс извлечения хранимой информации, т. е. процесс воспоминания.
Эксперименты, выполненные на добровольцах, свидетельствуют, что воспоминание прежнего опыта у человека связано с деятельностью височных отделов коры больших полушарий мозга. Хирургические операции на височных областях коры вызывали у пациентов воспоминания давно забытых событий. При электрическом раздражении височных долей пациенты сообщали о том, что у них возникали картины жизненных событий различной давности в мельчайших подробностях.

Молодой обыкновенный шимпанзе (Pan troglodytes) пробует на вкус все, постепенно понимая и запоминая, что съедобно, а что нет. Шимпанзе живут семейными группами. Взрослые особи знают, что можно есть, а что нельзя, где и как добывается корм. Молодой шимпанзе может не использовать малоэффективный метод проб и ошибок, а копировать поведение старших сородичей. В своей группе он обучается сложным навыкам, которые вырабатывались и оттачивались много веков и передавались из поколения в поколение.
Нельзя категорично утверждать, что именно височные доли отвечают за извлечение информации из структур долговременной памяти. Эта область коры морфофункционально связана с множеством подкорковых образований, включая такие, как гиппокамп, миндалины, маммиллярные тела переднего гипоталамуса.
Процесс извлечения информации из долговременной памяти сильно зависит от эмоционального состояния. Возбуждение структур лимбической системы (гиппокамп, миндалины) облегчает этот процесс. Клиницисты часто сообщают об уникальных способностях людей в состоянии аффекта, т. е. эмоционального перенапряжения, воспроизводить с мельчайшими подробностями давно забытые события.
Процесс утраты информационного следа в структурах центральной нервной системы происходит на всех этапах научения и формирования памяти. На уровне кратковременной памяти забывание происходит вследствие замещения одного сенсорного следа другим или его разрушения в результате фильтрации, например, в гиппокампе. В блоке долговременной памяти информация «стирается» в случае ее длительной невостребованности. Когда информация многократно извлекается из долговременной памяти (повторение, «зубрежка»), происходит ее закрепление в форме обновления электрохимических и морфологических носителей информации.
Утверждение о том, что на уровне третичной памяти забывания не происходит, следует оценивать критически. По ходу онтогенеза происходит накопление ошибок в механизмах репликации ДНК и транскрипции РНК. Поэтому развиваются возрастные изменения в процессах структурного обновления организма, приводящие, в свою очередь, к соответствующим функциональным изменениям. Возрастные явления не могут не затронуть и состояние центральной нервной системы и, в частности, механизмы памяти. Поэтому забывание на уровне третичной памяти будет происходить, по крайней мере, в форме искажения хранящейся информации. Убедительным доказательством правомерности данных рассуждений можно считать, хотя бы небезызвестную болезнь Альцгеймера — старческое слабоумие. Часть долговременной информации утрачивается у людей, больных алкоголизмом, при полиневрите вследствие недостатка витамина В1. Во всех этих случаях имеет место забывание долговременной информации из-за морфологических изменений мозга.

Детеныш саванного слона (Loxodonta africana) пьет воду. Поскольку для сосания материнского молока маленький слоненок пользуется ртом, он не знает, как использовать хобот для питья. Взрослые набирают воду хоботом и пускают струю в рот, но малыши ложатся и пьют с помощью губ. Хобот же для них является скорее помехой. Примерно до 5 месяцев слоненок может спотыкаться об хобот, с трудом представляя, как его можно использовать. Этот подвижный орган содержит порядка 40 000 мышц, и слоненку необходимо наблюдать за сородичами и практиковаться, чтобы научиться его контролировать.