в оглавление
назад
Пищеварительная система (продолжение)
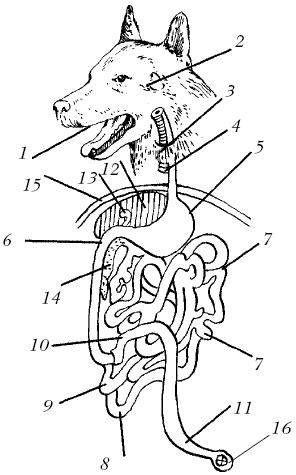
Схема пищеварительного тракта собаки (Canis lupus familiarus): 1 — ротовая полость; 2 — слюнные железы; 3 — глотка; 4 — пищевод; 5 — желудок; 6 — двенадцатиперстная кишка; 7 — тощая кишка; 8 — подвздошная кишка; 9 — слепая кишка; 10 — ободочная кишка; 11 — прямая кишка; 12 — печень; 13 — желчный пузырь; 14 — поджелудочная железа; 15 — диафрагма; 16 — анус.
Желудок (gaster, или ventriculus) представляет собой мускулистое расширение, обособленное от пищевода и кишечника и лежащее в передней части брюшной полости. Желудок служит главным образом для хранения пищи до того, как она поступит в кишечник, здесь же производится ее частичная химическая обработка.
Желудок хорошо развит у всех млекопитающих, но при этом его размеры, форма и строение значительно различаются. Наиболее просто устроен желудок однопроходных — в форме простого мешка; подобно пищеводу он лишен пищеварительных желез и весь выстлан плоским многослойным эпителием. В имеющем более сложную структуру желудке прочих млекопитающих различаются отделы: прилегающий к пищеводу отдел называют пищеводным, ближайший к сердцу — кардиальным (pars cardiaca), расширенный средний — фундальным, или донным (fundus ventricul), суженный конечный, граничащий с кишечником — пилорическим (pars pylorica). Последний отдел охвачен сильной кольцевой мышцей — пилорическим сфинктером, или привратником (pylorus), временно изолирующим желудочную полость от двенадцатиперстной кишки. Как правило, желудок своей длинной осью располагается поперек тела, однако его форма может сильно варьировать у разных групп. Кроме того, она может быть абсолютно разной у мертвого вскрытого животного и у живого, так как работающий желудок время от времени совершает мощные движения, сопровождающиеся заметными изменениями его формы. И все-таки, наиболее характерна форма, присущая, в частности, и желудку человека: сразу за пищеводом он отклоняется влево, расширяясь в большой мешок, затем восходит вперед по правой стороне и оканчивается у привратника, тем самым образуя J-образную фигуру. В некоторых случаях хорошо видны наметившиеся наружные перетяжки желудка (например, у хомяка (Cricetus cricetus), различных копытных).

Продольный разрез через желудок человека: 1 — пищевод; 2 — дно желудка; 3 — большая кривизна; 4 — привратниковая (пилорическая) часть; 5 — отверстие привратника; 6 — сфинктер привратника; 7 — складки слизистой оболочки; 8 — тело желудка; 9 — малая кривизна.
На фоне повсеместно присутствующих слизистых клеток, в зависимости от распространения пищеварительных желез в желудке млекопитающих можно выделить несколько типов эпителия. Начальный, пищеводный отдел желудка в большинстве случаев выстлан безжелезистым многослойным эпителием, напоминающим эпителий пищевода; по существу, эта часть желудка и представляет собой расширение пищеводной трубки. В остальных отделах желудочная выстилка представлена однослойным цилиндрическим эпителием, который образует сложные складки и богат пищеварительными трубчатыми железами, открывающимися в желудочных ямках. В кардиальном отделе имеются ветвистые железы, выделяющие слизистый щелочной секрет. Фундальный отдел характеризуется еще большим развитием слабо ветвящихся желез, которые, вдобавок к секреции слизи в шейках, вырабатывают в своей основной части пепсиногены (предшественники пепсинов, из которых последние отщепляются при низких значениях водородного показателя), прохимозин (предшественник химозина), липазы и соляную кислоту. При этом можно различить два сорта клеток, часто присутствующих вместе в одной и той же трубчатой железе, — главные клетки, вырабатывающие ферменты, и обкладочные клетки, производящие соляную кислоту. В прилегающем к кишечнику пилорическом отделе находятся железы, сходные с таковыми кардиального отдела, но секретирующие пепсиноген и прохимозин. Необходимо также отметить, что распределение различных типов эпителия не обязательно соответствует одноименным отделам желудка, т. е. по расположению эпителия нельзя судить о его гистологическом строении. Например, желудок у домовой мыши (Mus musculus) имеет приблизительно такую же форму, как у человека, но в отличие от него, выстлан в основном эпителием пищеводного типа.
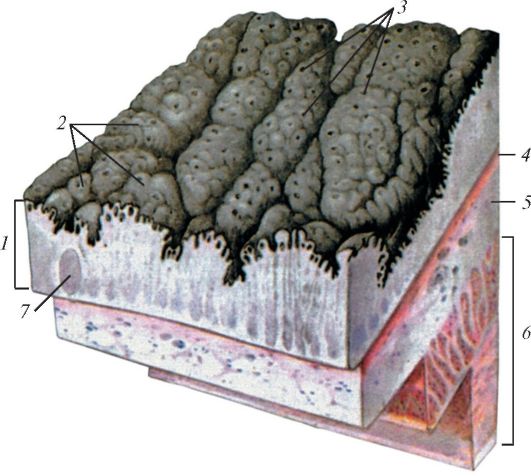
Строение стенки желудка млекопитающего: 1 — слизистая оболочка; 2 — желудочные поля; 3 — желудочные ямки; 4 — мышечная пластинка слизистой оболочки; 5 — подслизистая основа; 6 — гладкомышечная оболочка; 7 — одиночный лимфоидный узелок.
Секреты пищеварительных желез смешиваются и образуют желудочный сок. Соляная кислота (0,4-0,5 %) гидролизует жиры и крахмал, а создаваемая ею кислая среда в желудке способствует обеззараживанию пищи и активирует протеолитические ферменты. Пепсины при этом действуют как эндопептидазы, т. е. направлены на рзрушение пептидных связей внутри полипептидной цепи, подготавливая к дальнейшему расщеплению в кишечнике белки. Липазы заняты расщеплением жиров. Химозин способен створаживать молоко. Желудочный сок китообразных и муравьедов отличается повышенной активностью и разлагает даже такие стойкие вещества, как хитин, другими животными обычно неперевариваемые. Содержащиеся в желудочном соке слизь и бикарбонат защищающие слизистую оболочку желудка от твердых компонентов пищи и действия собственных секретов. Продукты желудочного расщепления питательных соединений имеют слишком крупные молекулы для прямого усвоения, и поэтому в желудке большинства млекопитающих, включая человека, может всасывать только вода.
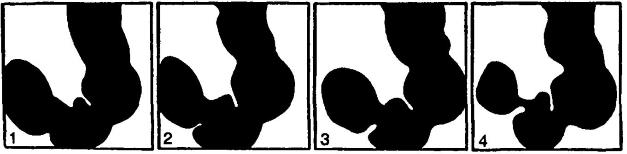
Рентгенограммы желудка человека, сделанные с определенными промежутками времени и демонстрирующие изменение его формы при обработке пищи.
Наиболее просто устроен желудок потребителей высокопитательных кормов. Однокамерный желудок насекомоядных, хищников и большинства приматов напоминает реторту и очень богат пищеварительными железами, подвергающими пищу интенсивной химической обработке. Появление в желудке новых структур и общее его усложнение происходит в результате специализации питания, например поглощения огромной массы грубых и малокалорийных кормов. Так, в пищеводе и желудке млекопитающих, потребляющих особенно жесткую растительную пищу (бобров (Castoridae), некоторых мышевидных грызунов), развивается выстилка из кератизированого эпителия, защищающая их стенки от повреждений. У панголинов (Manidae) зубы отсутствуют, а желудок, как и у птиц, состоит из двух отделов: железистого и пилорического. Второй отдел служит для перетирания пищи, он выстлан кутикулой с очень твердыми складками с роговыми зубцами и снабжен сильно развитой мускулатурой. Сходство с птицами усиливается тем, что в этом отделе обнаруживаются умышленно проглоченные камешки, играющие роль жерновов. С другой стороны, употребление грубой пищи может вести к усложнению не желудка, сохраняющего простое однокамерное строение, а кишечника; таковы непарнопалые, даманы и др. Желудок летучих мышей-вампиров (Desmodontinae) снабжен кишкообразным боковым выростом, в котором резервируется заглатываемая кровь.
Наконец, у растительноядных сумчатых, обезьян-тонкотелов (Colobinae), парнопалых и ряда других млекопитающих, особенно у питающихся грубыми и трудно перевариваемыми растительными кормами, развился сложный многокамерный желудок. У сирен, питающихся грубой водной растительностью, он резко разделен на 2 камеры. Желудок ленивцев является в первую очередь местом накопления пищи и может иметь до 5 камер. Он достигает 20-30 % массы тела и всегда набит едой, медленно поступающей в кишечник и продвигающейся по нему с малой скоростью (иногда до недели). Состоящий из 3-5 камер желудок китообразных, зубы которых не способны размельчать пищу, обеспечивает ее механическую обработку за счет перистальтических движений своих стенок.
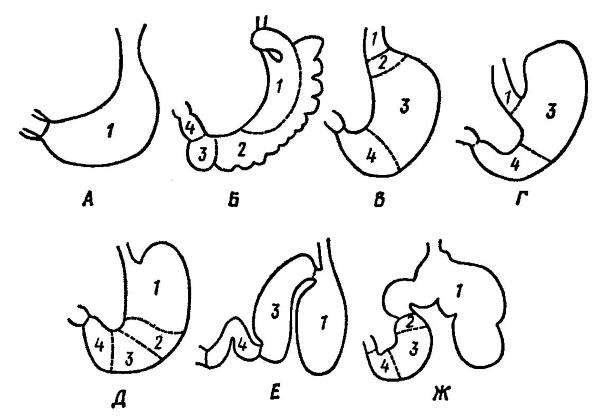
Схема строения желудков различных млекопитающих: А — ехидна (Tachyglossus); Б — кенгуру (Macropus); В — человек (Homo); Г — заяц (Lepus); Д — даман (Hyrax); Е — дельфин (Tursiops); Ж — корова (Bos); 1 — многослойный эпителий пищеводного типа, который может заходить в желудок; 2 — однослойный эпителий кардиального типа, встречающийся только у некоторых млекопитающих; 3 — однослойный эпителий фундального типа; 4 — однослойный эпителий пилорического типа.
Наиболее сложно в морфологическом и физиологическом плане устроен четырехкамерный желудок жвачных (исключение составляют представители примитивного семейства оленьков (Tragulida) с желудком из 3 камер). Слабо пережеванные комки пищи попадают по пищеводу в первую, наибольшую камеру — рубец (rumen), представляющий собой объемистый мешок. Здесь кормовая масса разминается мускулистыми стенками, подвергается мацерации и ферментируется в результате воздействия большого количества симбиотических микроорганизмов. Поддерживать адекватную среду для их деятельности помогает слюна, которая имеет слегка щелочную реакцию, снижающей кислотность сбраживаемой растительной массы. Количество слюны, выделяемой у жвачных в течение суток, составляет до трети от массы тела или до половины общего количества влаги, содержащейся в организме.
Из рубца мягкая пищевая масса небольшими порциями поступает в сетку (reticulum) — гораздо меньшую по размерам вспомогательную камеру, внутренняя поверхность которой образует систему перекрещивающихся складок с глубокими ямками между ними. Из сетки пищевая масса небольшими порциями отправляет обратно в рот. После вторичного, более тщательного пережевывания (у коров на это затрачивается от 7 до 9 часов в сутки), жвачка снова заглатывается. На этот раз она минует первые две камеры желудка и по особому пищеводному желобку в передней стенке сетки стекает непосредственно в третью камеру — книжку (omasum). Желобок этот настолько узкий, что через его просвет может проходить только жидкая пища; именно поэтому при первом заглатывании крупные пищевые комки наполняют рубец, но не книжку. В книжке процесс брожения усиливается. Слизистые оболочки ее внутренних стенок образуют параллельные складки, подобные книжным страницам. Через эти складки пища процеживается в четвертую камеру желудка — сычуг (abomasum). На его стенках присутствуют все три типа эпителия, характерные для различных отделов желудка млекопитающих — кардиального, фундального и пилорического. По сути, только сычуг и служит собственно желудком: именно здесь выделяется кислый желудочный сок, переваривающий пищу окончательно. Предыдущие камеры представляют собой расширения пищевода и образуют т. н. преджелудок, который выстлан многослойным эпителием и лишен пищеварительных желез. У некоторых жвачных пищевые комки отрыгиваются непосредственно из рубца и туда же возвращаются после дополнительного разжевывания во рту; в следующие камеры проходит уже сильно разжиженная пища.
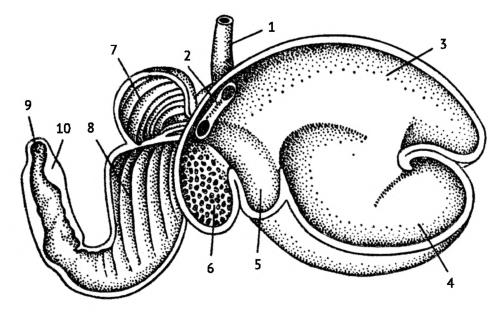
Желудок домашней овцы (Ovis orientalis aries): 1 — пищевод; 2 — пищеводный желоб; 3 — дорсальный мешок рубца; 4 — вентральный мешок рубца; 5 — преддверие рубца; 6 — сетка; 7 — книжка; 8 — сычуг; 9 — пилорус; 10 — поперечный замыкательный валик.
Биологическое значение жвачки состоит в том, что благодаря ей животное может наиболее полно использовать ценные компоненты корма, разрушая клетчатку и древесную паренхиму растительных клеток. Кроме того, жвачка дает возможность сокращать время пастьбы, когда животное в наибольшей степени подвергается опасности нападения хищников: наполнив за короткий срок свой вместительный рубец, оно неторопливо пережевывает и переваривает пищу уже в укромном месте, в тени, где его не беспокоят кровососущие насекомые. Принципиально схожие, но совершенно независимые приспособления выработались и некоторых других млекопитающих — например, у свиней, грызунов и двурезцовых сумчатых; роль бродильного агента в данном случае выполняют массово развивающиеся симбиотические дрожжевые грибки. Что касается хищников, то роль симбионтов в их пищеварении минимальна.
Симбиотические микроорганизмы, обитающие в сложном желудке жвачных, — это анаэробные бактерии и простейшие, среди которых преобладают инфузории. Количество симбионтов в содержимом рубца может превышать несколько сотен тысяч на 1 мл. Они поставляют ферменты-целлюлазы для расщепления клетчатки, и продукты ее гидролиза становятся доступными для усвоения животным-хозяином. Не менее важной является способность симбионтов самостоятельно синтезировать необходимые для хозяина органические вещества. Так, из переваренных углеводов ими синтезируются жирные кислоты, способные всасываться непосредственно в рубце. Синтез аминокислот (включая незаменимые) и белков происходит на основе неорганических азотистых соединений, например мочевины, которая в более типичных случаях удаляется из организма в качестве конечного продукта белкового распада. Экспериментально показано, что добавление мочевины в пищу коров позволяет животным дополнительно получить за счет деятельности симбиотических микроорганизмов до 100 г белка в день. Особенно важную роль играет синтез белка у животных, существующих на обедненной белковой диете, например у баранов (Ovis) и верблюдов (Camelus). У верблюдов мочевина не удаляется из организма с мочой, а заново поступает в желудок либо через его стенку, либо через слюну. Здесь она гидролизуется до углекислоты и аммиака, который затем используется в синтезе белка, что и позволяет верблюду существовать на диете из растений с низким содержанием белков. Синтезированный подобным образом белок выгодно отличается от растительных протеинов по аминокислотному составу. Наконец, практически все витамины группы В (особенно витамин В12) жвачные также получают благодаря их синтезу симбиотическими микроорганизмами.
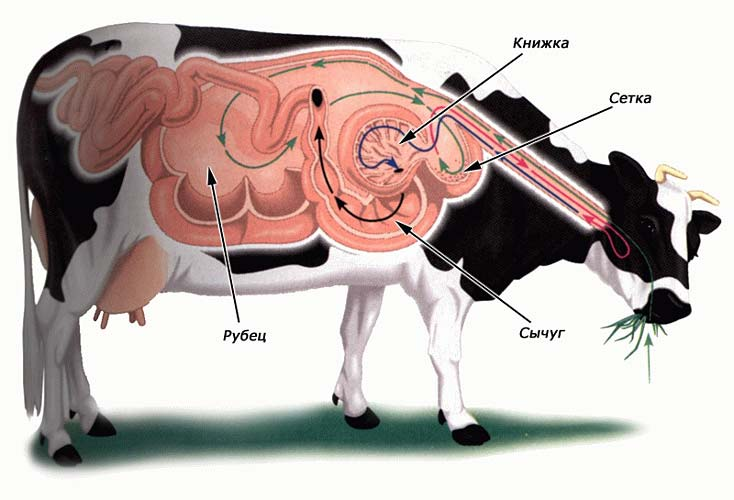
Схема движения пищи по пищеварительному тракту домашней коровы (Bos primigenius taurus).
Большинство млекопитающих способно рефлекторно выбрасывать содержимое желудка наружу, чтобы очистить его от ядовитых веществ. Мыши (Muridae), однако, на это не способны. Во-первых, их желудок отделен от пищевода слишком сильным клапаном, открывающимся и пропускающим пищу только в одну сторону. Во-вторых, их диафрагма не способна сокращаться достаточно сильно, чтобы вытолкнуть содержимое желудка наружу. Неспособность к рвоте мыши и крысы компенсируют превосходно развитыми вкусом и обонянием, благодаря которым они с легкостью распознают ядовитые вещества. Если зверек все же почувствовал себя плохо, он принимается есть глину, которая связывает яды в желудке. Неспособность к рвоте — не единственный недостаток строения пищеварительной системы мышей — отрыжка у них тоже отсутствует. К немногочисленным млекопитающим, неспособным опорожнять желудок через рот, относятся также зайцеобразные.
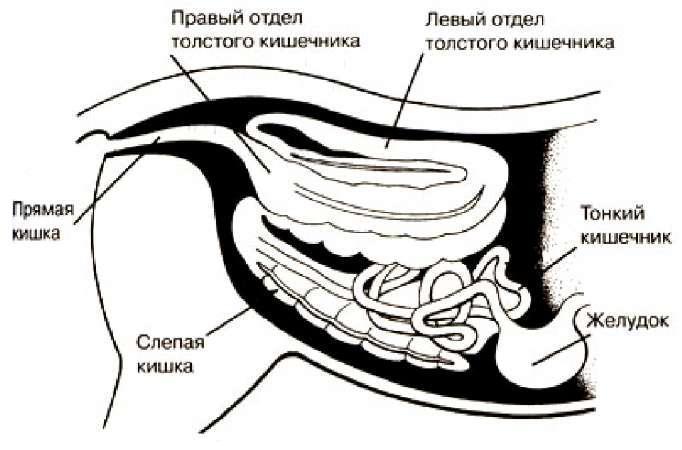
Схема пищеварительного тракта домашней лошади (Equus ferus caballus). Так как пищеварительный тракт непарнопалых короче и устроен проще, чем у жвачных, они не могут получить из растительности столько же питательных веществ, сколько получают жвачные. Об этом, в частности, свидетельствует наличие большого количества непереваренных остатков растений в фекалиях. Непарнопалые компенсируют это, потребляя относительно большее количество пищи и прогоняя ее через желудочно-кишечный тракт с предельной быстротой.
Подобно желудочным, стенки кишечника (intestinum) состоят из нескольких концентрических слоев, или «оболочек». Слизистая оболочка (tunica mucosa) начинается с эпителия. На большей части своей протяженности кишечник выстлан однослойным цилиндрическим эпителием; лишь для прямой кишки характерна выстилка из многослойного плоского эпителия. Эпителиальные клетки имеют различные функции, например секреторную или всасывающую. Редко разбросанные бокаловидные клетки в основном поставляют слизь для смазывания системы, более сложные железистые образования вырабатывают ферменты или иные химически активные вещества. Эпителий подстилает соединительная ткань, часто содержащая гладкие мышечные волокна — тонкую мышечную пластинку слизистой оболочки (lamina muscularis mucosae). Подслизистая основа (tela submucosa) обычно имеет значительную толщину и состоит главным образом из соединительной ткани, но также содержит многочисленные мелкие кровеносные сосуды, нервные клетки и сплетения симпатических нервных волокон (мейснеровы сплетения). Мышечная оболочка (tunica muscularis) состоит из двух мощных слоев гладких мышечных волокон: внутри находится кольцевой слой, способный сужать кишку, а снаружи — продольный слой, способный локально укорачивать ее. Между этими слоями и по их поверхности располагаются диффузные сплетения нейронов и парасимпатических нервных волокон (ауэрбаховы сплетения). Снаружи кишки окружены серозной оболочкой (tunica serosa), состоящей из эпителия и подстилающей его соединительной ткани, обращенной к целомической полости. По всей длине кишки прикреплены складкой брюшины (мезентерием) к спинной стенке брюшной полости.
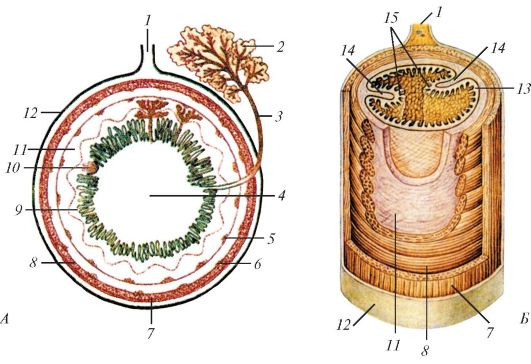
Обобщенная схема строения кишки млекопитающего, поперечное (А) и продольно-поперечное (Б) сечения: 1 — брыжейка; 2 — сложная пищеварительная железа; 3 — проток железы; 4 — просвет органа; 5 — подслизистое нервное сплетение (Мейсснера); 6 — межмышечное нервное сплетение (Ауэрбаха); 7 — продольный слой мышечной оболочки; 8 — круговой слой мышечной оболочки; 9 — собственная пластинка слизистой оболочки; 10 — одиночный лимфоидный узелок; 11 — подслизистая основа; 12 — серозная оболочка; 13 — слизистая оболочка; 14 — складки слизистой оболочки; 15 — ворсинки. В верхней половине рисунка изображена слизистая с железами и ворсинками, в нижней половине — только с ворсинками.
Кишечник млекопитающих подразделяется на два отдела — тонкий и толстый. Тонкий отдел кишечника (intestinum tenue) устроен следующим образом. Сразу за желудком располагается V-образная петля, которая носит название двенадцатиперстной кишки (duodenum). В ее просвет открываются желчный и панкреатический протоки. Двенадцатиперстная кишка без четкой границы переходит в тощую кишку (jejunum) и затем в подвздошную кишку (ileum). Иногда внешние отличия между разными частями тонкого кишечника совсем отсутствуют, так что его разделение бывает довольно условным.
В стенке тонкого отдела расположены мелкие железистые впячивания эпителия — общекишечные железы. В них вырабатывается большая часть ферментов, составляющих кишечный сок. Определенное количество ферментов может образовываться и в поверхностных клетках. Кроме того, эти клетки вырабатывают гормоны, которые уносятся кровью и стимулируют деятельность поджелудочной железы, желчного пузыря и общекишечных желез. Помимо упомянутых, в стенках двенадцатиперстной кишки имеются дуоденальные, или бруннеровы железы, секрет которых близок к таковому пилорических желез желудка.
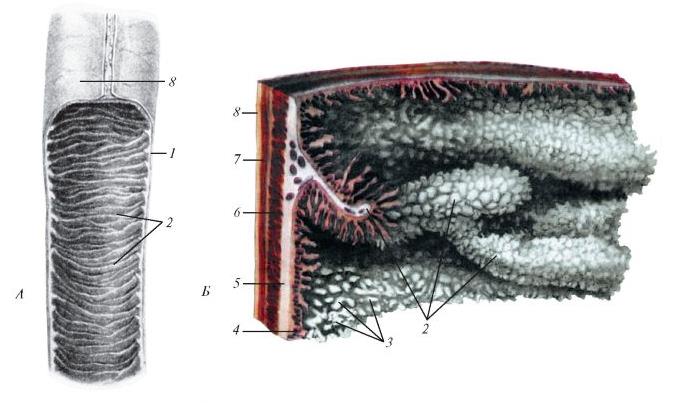
Рельеф слизистой оболочки тонкой кишки млекопитающего: А — участок кишки, вскрытой продольно; Б — поверхность слизистой оболочки и слои стенки кишки; 1 — стенка кишки; 2 — круговые складки; 3 — кишечные ворсинки; 4 — слизистая оболочка; 5 — подслизистая основа; 6 — круговой слой мышечной оболочки; 7 — продольный слой слизистой оболочки; 8 — серозная оболочка.
Таким образом, пищеварение в тонком кишечнике проходит в щелочной среде под влиянием желчи, панкреатического сока и кишечного сока. Все эти выделения очень обильны. Тонкий кишечник заполнен весьма жидким содержимым, т. н. химусом. Здесь происходит окончательное ферментативное расщепление всех видов органической пищи. Одни ферменты подготовляют сахара, другие расщепляют крахмал и жиры, активируют трипсин, разлагают пептиды на аминокислоты и отщепляют от нуклеотидов фосфорную кислоту.
Конечные продукты пищеварения и электролиты подлежат всасыванию в стенки тонкого отдела. Для повышения эффективности данного процесса требуется большая площадь кишечного эпителия. Это особенно актуально для крупных животных, поскольку у них вступают в действие соотношения поверхности и объема, а способная к всасыванию площадь должна быть приблизительно пропорциональна объему нуждающихся в питании тканей. Также должны приниматься во внимание высокая активность и интенсивность обмена веществ гомойотермного организма, из-за чего ему требуется большее количество пищи и соответственно большая всасывающая поверхность. У млекопитающих многократное увеличение площади кишечника обеспечивается на трех размерных уровнях. Прежде всего, для внутренней слизистой оболочки характерно наличие ворсинок (villi) — многочисленных пальцевидных выростов, имеющих микроскопические размеры. К каждой ворсинке подходит сеть кровеносных капилляров и один лимфатический канал. Далее, слизистый и подслизистый слои кишок собраны в циркулярные складки — клапаны Керкринга. Наконец, сами кишки сильно удлинены и сложно извиты.

Строение ворсинок тонкой кишки млекопитающего: 1 — ворсинки; 2 — сеть лимфатических и кровеносных сосудов слизистой оболочки; 3 — одиночный лимфоидный узелок; 4 — центральный лимфатический (млечный) капилляр; 5 — эпителий.
Сахара и аминокислоты всасываются непосредственно в кровь. Жиры в стенках кишечника заново синтезируются из глицерина и жирных кислот, затем поступают в кишечную лимфу, проходят через печень и, наконец, попадают в большой круг кровообращения.
Пищевой материал, который не смог усвоиться в тонком кишечнике, проходит в толстый (intestinum crassum). У ряда млекопитающих оба отдела разделяются илеоцекальной, или подвздошно-слепокишечной, заслонкой (valva ileocaecalis), не позволяющей пищевым частицам забрасываться обратно. Толстый кишечник представляет собой трубку значительного диаметра и неправильной формы, обычно имеющую ряд карманов и продольные мышечные тяжи (отсутствующие, однако, у хищных и жвачных). При виде с вентральной стороны толстый отдел имеет характерную форму перевернутой буквы U или вопросительного знака. Передний отрезок образует восходящую ветвь, идущую вперед вдоль правого бока тела. Тонкий кишечник входит в этот отрезок под углом, так что формируется Т-образное соединение. После перегиба следует нисходящая часть, идущая назад вдоль левой стенки полости тела. У приматов между восходящей и нисходящей частями имеется поперечный отрезок, а у большинства травоядных образуются дополнительные петли. Стенки толстого кишечника богаты трубчатыми железами, выделяющими смазывающую слизь.
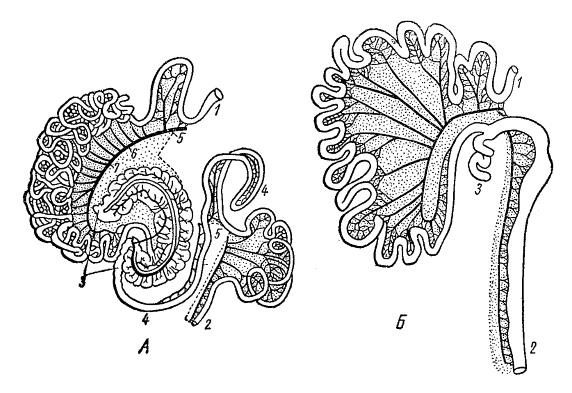
Кишечный тракт морской свинки (Cavia porcellus) (А) и лисицы (Vulpes vulpes) (Б): 1 — начало двенадцатиперстной кишки; 2 — конец прямой кишки; 3 — слепая кишка; 4 — петля ободочной кишки; 5-7 — разрезы кровеносных сосудов.
От начала толстого кишечника у большинства млекопитающих отходит слепая кишка (caecum). Обычно она выглядит как замкнутое на одном конце асимметричное начало восходящей части толстого кишечника, в которое сбоку открывается подвздошная кишка. Поверхность слепой кишки может нести спиральные перехваты, а на ее конце часто помещается более или менее выраженный червеобразный отросток, или аппендикс (appendix vermiformis). Даманы (Procaviidae), подобно птицам, имеют несколько слепых кишок. В стенках слепой кишки у многих млекопитающих находятся многочисленные лимфатические узлы, что делает ее важным органом иммунной системы.
В толстый кишечник попадают преимущественно трудноперевариваемые частицы. Слепая кишка представляет как бы загрузочный бункер для них. Особенно значительных размеров она достигает у нежвачных растительноядных — двурезцовых сумчатых, шерстокрылов, грызунов, зайцеобразных, непарнопалых. У них диаметр слепой кишки часто сравним с диаметром остального толстого кишечника или даже превышает его, а сама кишка бывает длинной, сильно извитой и обладает развитым червеобразным отростком. Это связано с развитием здесь более или менее интенсивных бродильных процессов с участием симбионтов — бактерий, грибков и простейших, аналогично протекающим в сложном желудке жвачных. Данное явление получило название заднекишечной ферментации.
Соответственно, слепая кишка развита тем сильнее, чем больше трудноперевариваемой растительной клетчатки поглощает при кормлении животное. Так, коала (Phascolarctos cinereus) потребляет листья и побеги эвкалиптов, в которых мало белка, но много токсинов. В связи с этим слепая кишка у этого вида сильно расширена, а ее длина достигает почти 2 м. У полевок (Arvicolinae), питающихся в большей мере вегетативными частями растений, она составляет 7-10 % от общей длины всех отделов кишечника, а у мышей (Murinae), питающихся семенами и лишь отчасти вегетативными частями растений, — 18-27 %. У всеядных форм слепая кишка относительно небольшая и особой роли в пищеварении не играет. Если у мокроносых приматов (Strepsirrhini) эта кишка еще хорошо заметна, то у сухоносых (Haplorrhini) она очень коротка — например, у человека от нее сохраняется лишь небольшой выступ с червеобразным отростком. У плотоядных видов слепая кишка почти неразвита или полностью отсутствует. В этой же связи варьирует и длина всего толстого отдела кишечника. У грызунов он составляет 29-53% общей длины кишечного тракта, у насекомоядных и рукокрылых — 26-30%, у хищных — 13-22%.
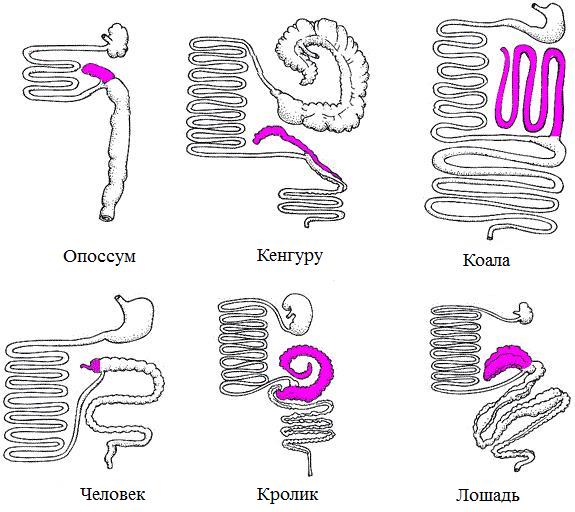
Относительные размеры отделов пищеварительного тракта различных млекопитающих; слепая кишка выделена лиловым.
Слепая кишка переходит в объемистую ободочную кишку (colon). В ней по чисто осмотическим причинам всасывается вода и электролиты, в результате чего относительно жидкая пищевая кашица постепенно превращается в более твердые экскременты. Непосредственного участия в пищеварении эта кишка обычно не принимает, хотя пищеварительные соки, принесенные из вышележащих разделов кишечника, могут продолжать свою работу и здесь. Также ободочная кишка населена бактериями, которые вызывают гниение остатков пищи и разлагают их на ряд веществ, имеющих сомнительное значение для организма или даже ядовитых. Остаточное количество питательных веществ и часть продуктов гниения также всасываются.
За ободочной кишкой следует прямая кишка (rectum), снабженная гладкими и поперечнополосатыми сфинктерами. Здесь формируются и накапливаются каловые массы, которые затем выводятся через анальное отверстие, или анус (anus). Характерный цвет калу придают желчные пигменты.

Каловые массы короткошерстного вомбата (Vombatus ursinus) имеют характерную кубическую форму.
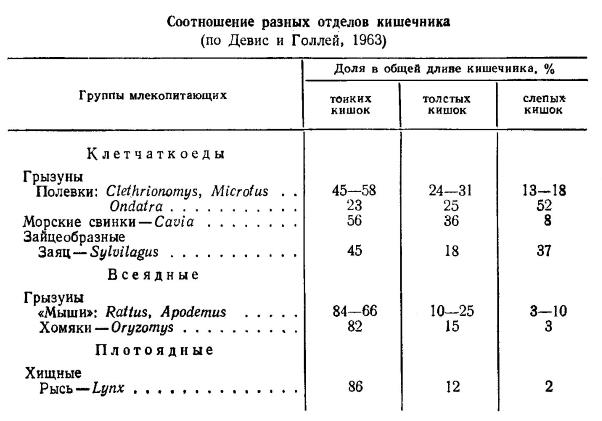
Общая длина кишечника у млекопитающих очень сильно варьирует, отвечая составу их пищи. Как правило, звери, питающиеся грубой растительной пищей, имеют более длинный кишечник, у всеядных видов он относительно короче, и наиболее короткий у плотоядных форм. Так, у летучих мышей длина кишечника превышает длину тела в 1,5-4 раза, у насекомоядных — в 2,5-4,5, у ласки (Mustela nivalis) — в 2,5, у кошки (Felis silvestris catus) — в 4, у собаки (Canis lupus familiaris) — в 6,5, у полуденной песчанки (Meriones meridianus) — в 5, у морской свинки (Cavia porcellus) — в 11,5, у лошади (Equus ferus caballus) — в 12, у свиньи (Sus scrofa domesticus) — в 15, у коровы (Bos primigenius taurus) — в 20, у овцы (Ovis orientalis aries) — в 29, у ла-платского дельфина (Pontoporia blainvillei) — в 32 раза. Таким образом, мелкие плотоядные формы, как правило, имеют самый короткий кишечник, а крупные растительноядные — наиболее длинный. В этой же связи изменяется и общая длина толстого отдела кишечника. У грызунов он составляет 29-53 % общей длины кишечного тракта, у насекомоядных и рукокрылых — 26-30 %, у хищных — 13-22 %. У молодых особей, вследствие высокой потребности в пище при более интенсивном обмене веществ кишечник относительно длиннее, чем у взрослых.
Отметим, что у некоторых млекопитающих внешне видимое разделение кишечника на тонкий и толстый отделы отсутствует. Например, у таких облигатных хищников, как хорек (Mustela putorius) и норка (Mustela lutreola), нет илеоцекального соединения, илеоцекальной заслонки и слепой кишки. По сути, отделы их кишечника различаются между собой только микроанатомически, гистологически и по характеру снабжения стенок сосудами. Собственно толстый кишечник у хорька и норки очень короткий. Нет слепой кишки и четкой границы между отделами при поверхностном осмотре у зубатых китов и большинства насекомоядных.
У части млекопитающих сохраняется клоака (cloaca) — карман, объединяющий выходы из пищеварительной, выделительной и половой систем. Наиболее отчетливо она выражена у однопроходных. По сути, клоака как таковая представлена у них лишь своим задним отделом, приблизительно соответствующим проктодеуму (proctodeum) рептилий. Передний отдел клоаки разделен на крупный дорсальный проход — копродеум (coprodeum), куда открывается прямая кишка, и вентральную часть — уродеум (urodeum), гомологичную мочеполовому синусу. Мочеточники и семяпроводы впадают в передний конец уродеума напротив отверстия мочевого пузыря. Благодаря такому их взаиморасположению моча может поступать в пузырь, не смешиваясь с фекальными массами.
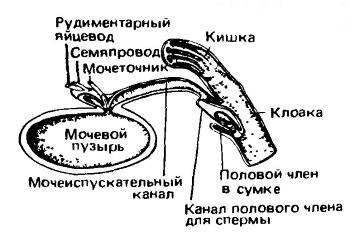
Сагиттальный разрез клоаки самца австралийской ехидны (Tachyglossus aculeatus).
У сумчатых клоака в большей или меньшей степени рудиментарна, представляя собой неглубокий проктодеум. Он снабжен мышцей-сфинктером, эквивалентным мышцам промежности плацентарных. У самцов ряда сумчатых мочеполовое и анальное отверстие разделены перинеальной мембраной, образуя перинеальную клоаку, или "полуклоаку", промежуточную между настоящей клоакой и полностью разделенными промежностью отверстиями плацентарных. Среди последних клоака имеется у тенрековых (Tenrecidae) и златокротовых (Chrysochloridae) из афросцирид, у некоторых грызунов — например, у аплодонтии (Aplodontia rufa), а также у сеноставок (Ochotona) из зайцеобразных. У остальных плацентарных клоака присутствует лишь в начале зародышевого развития, а затем разделяется на мочеполовой синус (sinus urogenitalis) и конечный отдел прямой кишки, которые независимо друг от друга открываются наружу.
В зависимости от месторасположения выделяют различные типы клоак. У однопроходных клоака базикаудальная, т. е. находится в основании хвоста, как и у рептилий. У сумчатых и плацентарных клоака прекаудальная (или ингуинальная). Соневидный опоссум (Dromiciops gliroides) имеет базикаудальную клоаку, как у однопроходных, что может говорить о его ранней дивергенции от остальных сумчатых.
вперед
в оглавление