назад
Подготовка к размножению, гон, спаривание (продолжение)
Наиболее ярким и внешне заметным компонентом полового поведения является вспрыгивание самца на самку — т. н. садка. Ложные (пробные) садки обычны в процедуре ухаживания самца за самкой. Однако садка при ухаживании не сопровождается обнимательным рефлексом, т. е. сжиманием тела самки с боков при помощи передних конечностей. При ложной садке у самца не происходит эрекции полового члена в полном объеме и он не выдвигается из препуциального мешка. У некоторых видов также имеет место взаимные садки, т. е. садки самца на самку, самки на самца и одной самки на другую самку. Анализ ложной и настоящей садки показывает, что сама садка и собственно коитус имеют раздельные нервные механизмы регуляции, т. е. являются самостоятельными рефлексами. При спаривании кошек (Felidae) самец не только совершает обнимательные действия передними лапами, но и захватывает зубами кожу самки в области холки и шеи. Не исключено, что при этом он наносит самке болевые раздражения, способствующее наступлению овуляции.

Копуляция у тигров (Panthera tigris). Тигры являются одиночными, не общественными животными. Готовая к спариванию самка оповещает об этом самцов громким рычанием и выделением пахучих меток. Прежде чем заслужить взаимное доверие, пара вначале рычит друг на друга и расходится несколько раз. Затем самка «соблазняет» самца: подталкивает его мордой, вылизывает его и катается по земле, размахивая в воздухе лапами, прежде чем лечь на землю и начать спариваться. Самец делает садку на самку в положении с согнутыми коленями, чтобы не раздавить ее собственным весом, рычит и кусает ее за шею. Самка отпрыгивает, ворчит и бросается на него, если спаривание не удается.
В случае половой рецептивности самки и успешной садки у самца рефлекторно происходит кровенаполнение пещеристых тел полового члена — эрекция, и его введение в влагалище или клоаку самки — интромиссия. При этом головка пениса соприкасается с половыми губами самки, что приводит к раздражению чувствительных нервных окончаний пениса. В результате кровенаполнение пещеристых тел полового члена самца еще больше усиливается и облегчается его введение во влагалище. В свою очередь, раздражение чувствительных нервных окончаний влагалища со стороны пениса усиливает реакцию замирания самки, повышает тонус гладких мышц влагалища и матки, приводит к развитию волны перистальтических движений половых путей, что обеспечивает эффект насасывания препуциальной жидкости и спермы после семяизвержения в матку, ее рога и яйцеводы.
Интромиссия может длиться от нескольких секунд (даманы (Procaviidae), полорогие (Bovidae)) до нескольких часов (носороги (Rhinocerotidae), некоторые куницы (Mustelidae)). В первом случае она завершается мощным толчком с эякуляцией, но у большинства видов самец для того, чтобы достигнуть извержения семени совершает ритмичные движения тазом и соответственно половым членом — фрикции. Половой член хряка в это время выполняет еще и вращательные движения относительно продольной оси. У домашних собак (Canis lupus familiaris) после эякуляции развивается «замок» — половой член самца защемляется мощными сокращениями кольцевых мышц влагалища самки, так что животные на протяжении некоторого времени не могут расцепиться (однако в случае опасности они легко разъединяются). Подобное явление также имеет место у волка (Canis lupus), лисицы (Vulpes vulpes) и многих куниц (Mustelidae).

«Замок» у собак (Canis lupus familiaris).
Продолжительность коитуса соответствует экологическим особенностям вида. Так, среди свинкоподобных грызунов (Hystricomorpha) она самая короткая у видов, обитающих в открытых ландшафтах и потому доступных для хищников. У самцов таких видов половой член зачастую снабжен дополнительными придатками, которые обеспечивают надежное раздражение рецепторного аппарата влагалища и, как следствие, сильнейшее половое возбуждение самки. Львице во время течки необходимо спариваться каждые полчаса в течение 5 дней и ночей. Самец способен совершать столь многочисленные акты в значительной мере потому, что его половой член дополнительно поддерживается бакулюмом, а также несет стимулирующие роговые шипики, что придает ему форму своеобразного ершика.
После успешного спаривания партнеры не торопятся расходиться. При этом они демонстрируют еще один распространенный стереотип полового поведения — обоюдное стремление оставаться рядом друг с другом. Это важное для воспроизводства поведение самца и самки способствует повторному спариванию и повышает вероятность оплодотворения. У копытных самец после спаривания остается рядом с самкой, зачастую касаясь ее тела своим. Если животные находятся на пастбище, то самец щиплет траву в непосредственной близости от самки. Самцы при этом периодически кладут голову на круп самки, что особенно выражено в отношениях жвачных (Ruminantia).
Спаривание как физиологический процесс регулируется нервной и гуморальной системами. Нервная регуляция коитального процесса имеет сложную природу. В регуляции этого явления принимают участие как структуры головного мозга (включая кору), так и центры спинного мозга. Как уже говорилось, половое желание самца находится под влиянием и внутренних (гормональных), и внешних условно-рефлекторных факторов. Ведущую роль в его формировании играет передний гипоталамус и преоптическая зона гипоталамуса. После их экспериментального разрушения половое поведение пропадает. В то же время имплантирование кастратам в преоптическую область гипоталамуса таблеток тестостерона восстанавливало весь комплекс полового поведения самцов, включая коитальный этап. Имплантирование в преоптическую зону гипоталамуса стерилизованных самок эстрогенов приводило к активизации стереотипов поведения, характерных для самок в состоянии эструса. .

Самец белого носорога (Ceratotherium simum) делает садку.
Однако не менее важная роль в регуляции полового поведения отведена и центрам спинного мозга, особенно на завершающем этапе половых отношений животных. Эрекция полового члена возникает под влиянием возбуждения полового центра крестцово-поясничной зоны спинного мозга. Эфферентное стимулирующее влияние половой член получает по парасимпатическим нервам от спинальных крестцовых корешков. При этом запускается сосудистая реакция, превращающая внутрипещеристое пространство из системы с небольшим объемом и низким давлением в систему большого объема с высоким давлением крови (до 60-100 мм рт. ст.).
В результате сосудодвигательных реакций, происходящих в пенисе, половой орган самца увеличивается в объеме, становится жестким, что существенно облегчает его проникновение через отекшие и увлажненные густой слизью наружные половые губы самки. Увеличившаяся в размерах головка пениса с невысоким давлением внутри нее действует в качестве функционального амортизатора, предотвращающего болевые ощущения и повреждения в принимающих пенис половых путях самки. Введение во влагалище полового члена и его фрикции вызывают раздражение механорецепторов головки пениса, что приводит к дальнейшему нарастанию возбуждения половых центров и — в завершении коитуса — к эякуляции и оргазму.
Эякуляция происходит под влиянием симпатического отдела вегетативной нервной системы с центрами в поясничном отделе спинного мозга. Эфферентный путь рефлекса семяизвержения проходит через симпатический узел по нервам тазового сплетения ко всем органам таза, включая придаточные половые железы и гладкомышечные образования в составе придатка семенника, ампулообразных расширений семявыносящих протоков.
В эякуляции выделяют два этапа. На первом этапе происходит сокращение гладких мышц добавочных органов — выносящих сосудов семенников, придатков семенников, семенного протока, семенных пузырьков и предстательной железы. В результате мочеиспускательный канал пениса самца заполняют секреты придаточных половых желез. На этом фоне запирается внутренний сфинктер мочевого пузыря, что исключает попадание мочи в состав спермы и спермы в мочевой пузырь.
На втором этапе отмечается расслабление наружного сфинктера мочевого пузыря и ритмичные перистальтические сокращения гладкой мускулатуры семявыносящих протоков. В результате давление спермы возрастает настолько, что она выбрасывается из полового члена самца на расстояние 30-60 см. После выброса спермы из мочеполового канала возбуждение половых центров самца резко падает. Эфферентный поток к половым органам самца как со стороны парасимпатических, так и симпатических волокон резко сокращается и развивается фаза детумесцепции. Детумесцепция — процесс быстрого развития ригидности полового члена с уменьшением его объема и медленного восстановления объема и давления крови в пещеристых телах.

Бонобо (Pan paniscus) спариваются лицом к лицу. Это считалось уникальным, но горилл (Gorilla) также наблюдали в подобной позиции. В целом же бонобо сексуально очень активны и используют для спаривания множество различных позиций. Также эти приматы являются единственными помимо людей существами, пользующимися во время поцелуев языком.
При интромиссии происходит выброс больших количеств гормона окситоцина из задней доли гипофиза. Концентрация этого гормона в крови домашних быков (Bos primigenius taurus) остается высокой начиная с момента эрекции полового члена и заканчивая семяизвержением. У быков после пятикратной эякуляции в крови окситоцин не обнаруживается. Чрезвычайно низка его концентрация и у самцов в отсутствие полового желания.
В момент эякуляции у быков регистрируют пиковые значения психо-гальванического кожного рефлекса. Этот рефлекс используется в качестве объективного критерия психоэмоционального возбуждения животных. Психо-гальванический кожный рефлекс животных и соответствующее психическое состояние предельного утомления после коитуса служат основанием для того, чтобы констатировать у животных при коитусе с эякуляцией состояние, близкое к оргазму человека. Эмоциональное сопровождение коитуса у таких животных, как высшие приматы и свиньи, не оставляет сомнений в наличии оргазма у животных и других видов. Именно оргазм является средством удовлетворения сексуальной потребности животных. В момент оргазма частота сердечных сокращений и частота дыхания как у самца, так и у самки увеличиваются в 2-3 раза. При этом на 50-80 мм рт. ст. повышается артериальное давление. После результативного коитуса наступает разрешающая фаза полового возбуждения — электрическая активность половых центров резко падает.
Оргазм самцов ограничивается несколькими секундами. Биологическое значение оргазма самок остается неясным. Овуляция и оплодотворение не имеют прямой связи с оргазмом. Овуляция и оргазм самок не синхронизированы. Оргазм самок выглядит как многократно повторяющийся процесс. Как эмоциональное состояние он развивается под влиянием раздражения пенисом рецепторов передней трети влагалища у самок, относящихся к группе животных с влагалищным типом осеменения, и под влиянием раздражения шейки матки у животных с маточным типом осеменения (кобыла, свинья). При полноценном коитусе в момент эякуляции у самок регистрируются судорожные сокращения мышц влагалища, которые, вероятно, совпадают с оргазменными ощущениями самки. В процессе одного полового акта оргазм у самки может наступать многократно. Исследования показали, что половое возбуждение и коитальная восприимчивость самок разобщены во времени. Только после того, как половое возбуждение достигает определенного уровня, а именно становится непреодолимым, самка демонстрирует коитальную рецептивность. Оргазм, клиническим выражением которого являются сладострастные судорожные сокращения мышц влагалища, возникает вследствие того, что гипоталамус достигает некоего критического уровня возбуждения и приводит к выбросу через гипофиз в кровь большой дозы гормона окситоцина. Однако остается неясным, что у самок первично — эмоция сладострастия или сокращения влагалища.
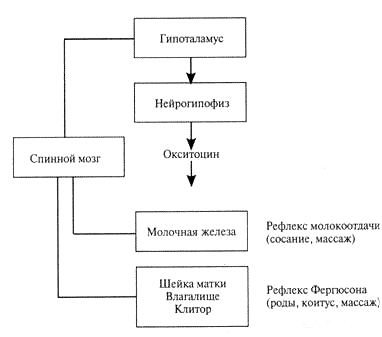
Нейрорефлекторные пути высвобождения окситоцина гипоталамо-гипофизарной системой самок.
Оргазм у самки не вызывает такого сильного торможения полового возбуждения, как у самцов. Доказательством этому служат практические наблюдения за поведением самок при спаривании. Так, овцематка в состоянии эструса проявляет коитальную восприимчмвость по отношению к нескольким самцам подряд на протяжении очень короткого времени. После последовательного спаривания с несколькими самцами самка не выглядит такой опустошенной, как каждый из этих самцов при однократном коитусе.
Определенно, часть самок вообще не испытывает оргазм. Очевидно и то, что молодые самки испытывают болезненные ощущения при первом половом акте, о чем свидетельствует соответствующая вокализация. Оргазменные ощущения у самок появляются в более зрелом возрасте. Отсутствие оргазма у самок не исключает успешного оплодотворения. Это наглядно подтверждает практика искусственного осеменения, получившая широкое распространение в современном животноводстве. Процедура искусственного осеменения, как правило, представляет для самки стрессовую ситуацию, поскольку предполагает поимку животного, физическое ограничение движений, а зачастую и болезненные манипуляции в половых путях. Искусственное осеменение, как минимум, неприятно, а то и вовсе болезненно, т. е. процедура искусственного введения спермы в половые пути коров, свиноматок, кобыл и других животных определенно сопровождается торможением полового возбуждения, не способствует перевозбуждению гипоталамуса, выбросу окситоцина и развитию оргазменных сокращений влагалища. Однако результативность искусственного осеменения достаточно высока. Следовательно, овуляция у самок не имеет прямолинейной связи с половым возбуждением и оргазмом.
Таким образом, половое возбуждение самки, ее коитальная восприимчивость, оргазменные ощущения и овуляция являются самостоятельными процессами с собственными нейрогуморальными регуляторными механизмами. Поэтому они могут быть разобщены во времени. Синхронизация этих процессов имеет место при естественном половом поведении здоровых животных. Половое влечение с положительным подкреплением в форме оргазменных ощущений, характерное для зрелых самок, лежит в основе случаев инициации коитуса со стороны самки. Разобщенность оргазма самки и ее коитальной рецептивности выступает одной из причин промискуитетности полового поведения самок.
У большинства видов коитальная восприимчивость самки развивается за некоторое время до овуляции. Опережающее овуляцию поступление сперматозоидов в половую систему самки физиологически оправдано, поскольку продвижение сперматозоидов в яйцеводы требует определенного времени. Собственно оплодотворение как слияние мужской и женской половых клеток происходит только в верхней части яйцевода. Поэтому предовуляторный коитус идеально синхронизирует движение сперматозоидов и яйцеклеток к месту оплодотворения.

Самец большего рыжего кенгуру (Macropus rufus) делает садку. Очевидна разница в размерах и окрасе самца и самки.
Первые копуляции половых партнеров приходятся на начало эстрального цикла самки с несколькими повторениями по мере его развития. Частота необходимых коитальных контактов различна у разных видов животных и, кроме того, зависит от индивидуальных особенностей партнеров. Судя по всему, частота спариваний больше зависит от полового желания индивидуумов, чем от видовой или породной принадлежности половых партнеров. По мере развития эстрального цикла частота спариваний уменьшается. Отмечена и суточная динамика частоты спариваний. В начале светового дня она выше, чем в середине и в конце дня. Эта цикличность объясняется, прежде всего, потребностями самца во времени, которое ему необходимо на накопление запасов половых продуктов и восстановление либидозного состояния.
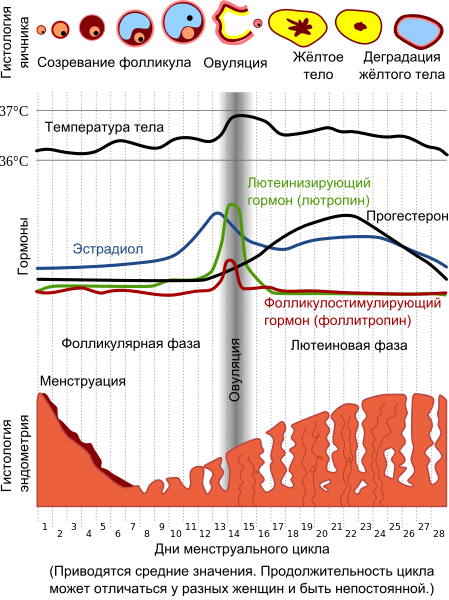
По окончании брачного периода возбуждение самцов и самок спадает, в половом цикле самок наступает стадия торможения (лютеиновая стадия, или стадия желтого тела), характеризующаяся угасанием полового тонуса, уменьшением яичников, ослаблением гиперемии гениталий. Для первой фазы стадии торможения — послетечкового восстановления, или метэструса (metaoestrus), характерны резкое снижение активности вагинального эпителия, отторжение базального слоя клеток, а также формирование желтых тел. Уровень эстрогена падает, начинает секретироваться прогестерон.
У хорьков овуляция индуцированная, т. е. наступает лишь после совершения полового акта. Если спаривания так и не произошло, самка одомашненной формы лесного хорька (Mustela putorius furo) выходит из течки с большим затруднением. Высокий уровень эстрогена через некоторое время начинает мешать костному мозгу вырабатывать эритроциты, вследствие чего зверек может погибнуть от апластической анемии. Самки диких хорьков, по-видимому, способны самостоятельно выходить из течки без каких-либо существенных последствий, но и у них без спаривания течка иногда растягивается до 6 недель. Покрытие стерилизованным самцом также способствует выходу самки из течки, как и спаривание с интактным. Это иногда используется вместо стерилизации или гормональных препаратов, если в будущем от самки планируется получать потомство, но в этом случае может развиваться ложная беременность.
Во второй фазе — послетечкового покоя, или диэструса (dioestrus), — половые органы постепенно возвращаются к состоянию покоя, а вагинальный эпителий теряет свою активность. Этому соответствует низкий уровень секреции всех гормонов, производимых яичниками. На межтечковый покой приходится около половины продолжительности всего эстрального цикла, в это время происходит общее восстановление сил. У полициклических видов относительно короткий диэструс сменяется новым проэструсом практически без наступления анэструса; у монциклических видов стадя покоя длится до следующего года. Сигналом к началу нового эстрального цикла служат лизис желтого тела и переход яичника в фолликулярную фазу овариального цикла.