назад
Размножение и связанные с ним явления
Размножение — важнейший биологический процесс, обеспечивающий поддержание и увеличение численности вида, возможность его расселения и, в конечном итоге, успех борьбы за существование. Если животное созрело для размножения и если внутренние и внешние стимулы благоприятствуют этому процессу, то у животного формируется мотивация полового поведения с очень высоким уровнем приоритетности, подавляющая все прочие виды поведения. Половая доминанта формируется под влиянием разных процессов и явлений, включая особое состояние нервной и эндокринной систем, высокую чувствительность к феромонам и внешним стимулам другой модальности.
У млекопитающих процесс размножения имеет как некоторые общие для высших позвоночных черты (внутреннее оплодотворение, забота о потомстве), так и специфические особенности (живорождение, выкармливание детенышей молоком). Особенности размножения у млекопитающих имеют ясно выраженный приспособительный характер и связанны с особенностями жизненных условий. Скорость размножения каждого конкретного вида зависит от времени полового созревания, длительности промежутка между рождениями и числа детенышей в выводке. Благодаря развитости и разнообразию форм заботы о потомстве на всех этапах его развития общая плодовитость млекопитающих относительно невелика.
Пребывая внутри материнского организма, зародыш растет за счет веществ, получаемых через специальный орган — плаценту. Из современных представителей класса лишь однопроходные откладывают крупные, богатые желтком яйца. У всех млекопитающих после рождения молодого животного его вещественная связь с матерью не прекращается, так как детеныш питается молоком. Однако и после завершения молочного кормления связи между родителями и потомством, как правило, какое-то время сохраняются. Это обеспечивает возможность обучения молодняка — передачи накопленного родителями индивидуального опыта потомству. Такая связь поколений, позволяющая накапливать и суммировать популяционно-видовой опыт, получила название сигнальной наследственности, или сигнальной передачи информации. Она хорошо развита у птиц и особенно характерна для млекопитающих. Качественно иной уровень это явление приобретает у обладающего разумом и второй сигнальной системой человека, основой деятельности которого служат производственно-общественные отношения.
Хотя размножающаяся пара обычно дает начало более чем двум потомкам, популяции в большинстве случаев не растут (если вообще растут) с большой скоростью. Поскольку популяция не может расти бесконечно, многие молодые животные должны погибнуть, а различия между ними обеспечивают избирательную смертность, позволяя выжить более приспособленным. Признаки, которые обеспечивают адаптивные преимущества, будут сохранены, а если они наследственны, то их передадут потомкам и они постепенно станут преобладать в популяции. Таким образом, естественный отбор приводит к тому, что в последующих поколениях возрастает приспособленность к определенным условиям среды. Видовые особенности представляют собой совокупный результат действия естественного отбора на отдельных представителей данного вида.
Однако было бы неверно утверждать, что животные стремятся «к пользе вида», — на самом деле они должны увеличивать свою собственную приспособленность, которая часто эквивалентна количеству потомков, произведенных родителями на свет и доживших до репродуктивного возраста. Отбор воздействует на генетический материал, лежащий в основе признаков каждой особи, так что задача последней — обеспечить сохранение и передачу генов, для которых она служит просто временным вместилищем. Иногда животное помогает своим сородичам, причем как будто вопреки собственным интересам, но такое поведение приносит выгоду его собственным генам и способствует улучшению общей адаптированности.
Ocoби ведут себя так, чтобы их успех в размножении был наибольшим, а поскольку стратегия размножения составляет основу организации сообщества, приспособления к достижению конечного результата в сообществах млекопитающих чрезвычайно разнообразны. Самцы и самки в этом отношении радикально различаются: производство сперматозоидов не так затратно, как яйцеклеток, при этом только самка несет все энергетические «расходы» на вынашивание и выкармливание потомства. Самцы могут легко увеличить свой репродуктивный успех за счет спаривания со многими самками. Самки же могут выносить сравнительно небольшое количество детенышей, поэтому увеличить свой вклад в будущее поколение они могут, только если повысят «качество» потомства, в том числе за счет спаривания с наиболее успешным (т. е. эволюционно приспособленным) самцом.
Самки — ресурс, за доступ к которому соревнуются самцы. Напряженный естественный отбор между соревнующимися самцами называется половым. Он объясняет, почему у многих млекопитающих самцы спариваются с несколькими самками и почему первые часто крупнее вторых. Рослый самец может устранить больше соперников, обеспечив себе возможность спаривания с большим количеством самок, и благодаря этому оставить больше потомков. Если его размеры и мощь перейдут к его сыновьям, те, в свою очередь, станут успешными доминирующими самцами. Точно так же и самки, поведение которых способствует спариванию с наиболее приспособленными самцами, имеют больше шансов произвести на свет более успешных потомков.
Иная ситуация складывается у видов, у которых репродуктивный успех самцов определяется участием в заботе о потомстве. Например, у псовых выживание молодняка зависит от обеспеченности пищей, поэтому самец едва ли способен прокормить более одной, максимум двух семей. В данном случае отбор благоприятствует моногамии и половой диморфизм не столь значителен. Однако не очень понятно, почему половые различия преобладают в основном среди крупных животных. Возможно, что у них меньше энергетические потребности, и они могут позволить себе больше вкладывать в наращивание мышц и вооружений.
Половая зрелость
Для того чтобы участвовать в размножении, индивидуум должен достичь определенного возраста — возраста половой и физиологической зрелости. Сроки ее наступления у млекопитающих очень сильно варьируют, но, как правило, у мелких форм с небольшой продолжительностью жизни способность к воспроизводству наступает раньше. Рыжая полевка (Myodes glareolus) способна приносить потомство уже в 1 месяц, степная пеструшка (Lagurus lagurus) — в 1,5 месяца, домовая мышь (Mus musculus) — в 2-3 месяца, серая крыса (Rattus norvegicus) — в 3 месяца, ондатра (Ondatra zibethicus) — в 5 месяцев, заяц-русак (Lepus europaeus) — в 1 год. Более крупные звери начинают размножаться позже: рыжая лисица (Vulpes vulpes) — в 1-2 года, волк (Canis lupus) и лось (Alces alces) — в 2-3 года, бурый медведь (Ursus arctos) — в 3-4 года, коала (Phascolarctos cinereus) — в 4 года, синий кит (Balaenoptera musculus) — в 8 лет (до 20 лет), азиатский слон (Elephas maximus) — в 15-20 лет. Многие млекопитающие начинают размножаться, еще не достигнув взрослых размеров. Самцы и самки многих видов становятся половозрелыми не одновременно. Например, у индийского носорога (Rhinoceros unicornis) самка впервые участвует в размножении в 3-4-летнем возрасте, а самец — в 7-9 лет. Интересный случай представляют самки горностая (Mustela erminea). Они становятся способными к оплодотворению в возрасте всего нескольких дней или недель и иногда могут оплодотворяться взрослым самцом еще в гнезде, однако из-за задержки развития зародышей потомство появляется только на следующий год.
Половые циклы
Размножение не является постоянной формой активности и отнюдь не хаотично. Оно ритмично, т. е. упорядочено во времени. Многие млекопитающие спариваются в четко определенное время года, дабы приурочить роды к самому благоприятному для выкармливания молодых периоду — как правило, к весне. Появление на свет детенышей в весеннее время наиболее целесообразно, поскольку весной отсутствуют губительные для новорожденного холода и расширяется за счет пробуждения растительности рацион матери.
Готовность к размножению обеспечивается сложным регуляторным механизмом, действующим по стимулам как внутренней (взаимодействие нервной и гуморальной систем), так и внешней среды (фотопериодизм, температура, характер пищи). В умеренных и высоких широтах внешним сигналом служит изменение длины светового дня. Для видов с весенним размножением сигнальное значение имеет увеличение длины дня (большинство млекопитающих), для спаривающихся осенью копытных — ее сокращение, а для волков (Canis lupus) — короткий зимний день. В более низких широтах значение сигнального фактора могут принимать другие периодические природные явления, предшествующие приближению благоприятного для размножения времени: в пустынях и саваннах — осадки, в тропических лесах — созревание основных кормовых растений и т. п. Даже в районах с устойчиво теплым и влажным климатом размножение млекопитающих обычно сезонно, хотя у разных видов может приходиться на разные времена года. Помимо годовых, некоторые виды животных демонстрируют суточные колебания половой активности.
Однако половую цикличность самок и изменчивость половой активности самцов не следует рассматривать как прямую реакцию на изменение факторов среды. Внешние факторы лишь создают благоприятные (или неблагоприятные) условия для работы внутренних механизмов ритмичности физиологических процессов, обслуживающих размножение. Это справедливо прежде всего в отношении сезонной цикличности половой активности. Неблагоприятные факторы среды сдерживают физиологические механизмы или внутренние биологические часы. В благоприятных условиях среды эти механизмы работают постоянно без торможения со стороны внешних факторов. Поэтому самки домашних собак при совместном проживании с человеком в теплом помещении приходят в состояние половой охоты дважды в год, коровы — ежемесячно. В дикой природе половая активность всех этих животных имеет другой уровень напряжения.
Если у самцов гаметы образуются постоянно после наступления полового созревания, то у самок они созревают циклически. Параллельно этому процессу изменяется уровнь женских половых гормонов и происходит ряд изменений в репродуктивной системе. В центре механизма регуляции половых циклов находится гипофиз, получающий сигналы из различных частей тела и дающий на них соответствующие ответы. Общее стимулирующее влияние на гипофиз оказывают попадающие в организм с пищей и образующиеся в тканях витамины и гормоны. С другой стороны, его работа корректируется корой головного мозга, в которой в результате восприятия внешних раздражений возникают соответствующие импульсы. Так, изменение светового режима при посредничестве коры побуждает переднюю долю гипофиза к выделению гонадотропного гормона фолликулина, распадающегося на две фракции — пролан А и пролан Б. Первая из них обусловливает созревание фолликулов и овуляцию, а вторая стимулирует развитие желтого тела. Другой важный влияющий на гонады гормон — лютеостерон — вырабатывается уже желтым телом.
Последовательный ряд физиологических процессов созревания яйцеклетки и фолликула получил название овариального, или яичникового, цикла. Он состоит из нескольких стадий, которые в свою очередь можно разделить на отдельные фазы.
Стадии: Фазы (соответствуют фазам эстрального цикла):
I. Уравновешивания, или покоя 1. Предтечкового покоя
II. Возбуждения, или фолликулярная 2. Подготовки к течке, или предтечки
III. Овуляторная 3. Течки
IV. Торможения, или лютеиновая (желтого тела) 4. Послетечкового восстановления
5. Послетечкового покоя
В случае оплодотворения наступают еще стадии:
V. Беременности
VI. Лактации
Циклическим процессам, происходящим в яичниках, яйцеводах и матке у самок сумчатых и плацентарных млекопитающих соответствуют эстральные циклы — периодически повторяющиеся изменения в их влагалище. Эстральный цикл напрямую зависит от эндокринных функций яичников и состоит из нескольких стадий, каждая из которых характеризуется определенным функциональным и морфологическим состоянием слизистой влагалища и его гладкомышечного слоя. У разных млекопитающих длительность цикла значительно варьирует, причем она обычно возрастает с увеличением размеров животных. Так, у сирийского хомячка (Mesocricetus auratus) цикл продолжается 4 сутки, у черной крысы (Rattus rattus) и домовой мыши (Mus musculus) она составляет 4-6 суток, у морской свинки (Cavia porcellus) — 16-18 суток, у домашней коровы (Bos primigenius taurus) — 21 сутку, у лошади (Equus ferus) — 19-25 суток, у саванного слона (Loxodonta africana) — 16 недель и т. д. Соотношение между длительностью отдельных стадий эстрального цикла также видоспецифично. Например, у серой крысы (Rattus norvegicus) проэструс и эструс длятся по 12 ч, метэструс — 21 ч, диэструс — 57 ч; у домашней свиньи (Sus scrofa domestica) соответственно — 3, 2-3, 3 и 11-13 суток. Продолжительность цикла может меняться под действием различных внешних факторов (стресс, нагрузки и пр.). Физиологические изменения организма самки при смене стадий эстрального цикла происходят параллельно с изменениями ее психики. Иногда эти два явления расходятся во времени.
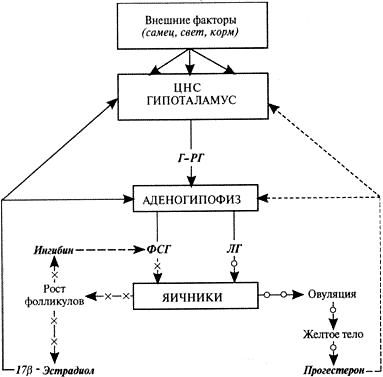
Схема нейроэндокринной регуляции половых процессов у самок. Сплошными линиями показана прямая и положительная обратная связь (стимуляция), прерывистыми — отрицательная (блокирование). Вырабатываемый в гипоталамусе гонадотропин-рилизинг-гормон стимулирует выделение из аденогипофиза фолликулостимулирующего и лютеинизирующего гормонов. Фолликулостимулирующий гормон активирует рост и развитие фолликулов и выработку ими эстрогенов и ингибина — ингибин избирательно блокирует секрецию фолликулостимулирующего гормона. Эстрогены, воздействуя на органы-мишени (центральную нервную систему, вторичные половые органы), индуцируют проявление течки, полового возбуждения и охоты. В конце проэструса и в начале эструса, когда концентрация прогестерона низкая, преовуляторный пик 17β-эстрадиола инициирует циклический выброс гонадотропин-рилизинг-гормона, фолликулостимулирующего и лютеинизирующего гормонов. Преовуляторный пик лютеинизирующего гормона индуцирует созревание преовуляторных фолликулов, их овуляцию, базальная секреция этого гормона — формирование желтых тел и выработку ими прогестерона. Прогестерон через механизмы отрицательной обратной связи контролирует секрецию гонадотропин-рилизинг-гормона, фолликулостимулирующего и лютеинизирующего гормонов: высокие его концентрации блокируют, а низкие стимулируют выделение этих гормонов.
В период размножения овуляция и течка повторяются вплоть до наступления оплодотворения. В зависимости от количества половых циклов различают два типа млекопитающих. У самок полициклических видов циклы при отсутствии беременности повторяются много раз на протяжении года. Таковы, в частности, обезьяны (Simiiformes), лев (Panthera leo), гиены (Hyaenidae), большинство копытных, грызунов и др. Самки моноциклических видов имеют в году только один цикл, приуроченный к определенному сезону. Таковы однопроходные, некоторые сумчатые, насекомоядные (кроты (Talpa)), грызуны (бобры (Castor), сурки (Marmota)), копытные (косули (Capreolus))), многие хищные (куницы (Mustelidae), собаки (Canidae), медведи (Ursidae), ластоногие (Pinnipedia)), китообразные и т. д. Самки большинства млекопитающих спариваются только в период течки; лишь у относительно немногих видов они рецептивны (коитально восприимчивы) большую часть времени. Само собой разумеется, и в том и в другом случае оплодотворение яйцеклетки становится возможным только после овуляции.
Половые циклы следуют друг за другом до тех пор, пока самка может размножаться. По достижении определенного возраста эта способность утрачивается, наступает т. н. климактерический период (climax). У самцов половая цикличность выражена слабее и заключается в основном в изменениях интенсивности сперматогенеза (и вместе с тем размера семенников) и в изменении поведения животных. Таким образом, как незрелые в морфофункциональном отношении животные, так и старые особи в размножении не участвуют. В связи с этим применяется особый термин — «репродуктивный период жизни». Он обозначает возрастные рамки индивидуума, в которых животные могут проявлять результативную половую активность.
Весь круг явлений, связанный с процессом размножения млекопитающих, складывается из нескольких этапов: подготовки к размножению; совокупления; беременности; родов; подсосного периода и воспитания потомства; распада семей и расселения молодых.
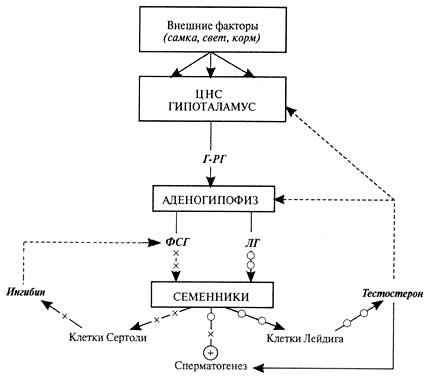
Схема нейроэндокринной регуляции половых процессов у самцов. Сплошными линиями показана прямая связь (стимуляция), прерывистыми — отрицательная обратная связь (блокирование). Вырабатываемый в гипоталамусе гонадотропин-рилизинг-гормон стимулирует выделение из аденогипофиза фолликулостимулирующего и лютеинизирующего гормонов. Лютеинизирующий гормон стимулирует выработку клетками Лейдига гормона тестостерона. Тестостерон поддерживает сперматогенез, половое влечение и через механизмы отрицательной обратной связи контролирует секрецию гонадотропин-рилизинг-гормона, фолликулостимулирующего и лютеинизирующего гормонов: высокие его концентрации блокируют, а низкие — стимулируют выделение этих гормонов. Фолликулостимулирующий гормон стимулирует выработку клетками Сертоли гормона ингибина. Ингибин через механизмы обратной связи подавляет секрецию фолликулостимулирующего гормона.
вперед
в оглавление