в оглавление
назад
Сосуды
Сосуды кровеносной системы, как и клетки крови, развиваются из эмбриональной мезенхимы. Как только в теле раннего зародыша обозначаются потоки питающих жидкостей, прилежащие мезенхимные клетки собираются около таких потоков и окружают их, образуя тонкую сплошную стенку. Так идет раннее развитие всех основных кровеносных сосудов. На более поздних стадиях клетки, выстилающие установившееся кровяное русло, образуют наружные разрастания, превращающиеся в мелкие кровеносные сосуды. В течение всей жизни животного в соответствии с изменениями в тканях тела таким способом часто возникают новые сосуды. После ранения или повреждения разрывы сосудов зарастают, и кровообращение восстанавливается. Рост ткани или органа сопровождается появлением дополнительных сосудов.
Внутренняя выстилка сосудов образована эндотелием, состоящим из одного слоя тонких листовидных клеток, соединяющихся друг с другом краями. Морфологические особенности в виде отверстий и пор здесь отсутствуют, и даже промежутки, наблюдаемые между клетками на ультраструктурном уровне, обычно закрыты тончайшей мембраной. В норме циркулирующая жидкость нигде не соприкасается с тканевой жидкостью и клетками, лежащими вне сосудов, хотя большая часть составляющих плазмы может свободно проникать через эндотелиальную мембрану.
К сосудам кровеносной системы относится сердце, артерии (несущие кровь от сердца тканям тела), капилляры и аналогичные структуры (обычно — мелкие сосуды, соединяющие артерии с венами), вены (возвращающие кровь в сердце), дополнительные лимфатические сосуды (сосуды, способствующие оттоку жидкости от тканей).
Капилляры представляют собой мельчайшие сосуды, диаметр которых достаточен для прохождения лишь одного эритроцита. В относительно малоактивной ткани они могут сжиматься и пропускать малое количество крови, а то и вовсе закрываться. Строение капилляров простое. Они берут начало от артериол или вен (в воротных системах) и, извиваясь, проходят между клетками большинства тканей тела таким образом, что любая из них оказывается поблизости от какого-либо капилляра. Степень васкуляризации ткани капиллярами обычно пропорциональна метаболической активности этой ткани. Из дистальных отделов капилляров кровь продолжает свое течение по более крупным сосудам (чаще всего по венам). В большинстве случаев стенки капилляров состоят просто из тонкого сплошного слоя эндотелиальных клеток, которые иногда дополняются соединительнотканными, другими мезенхимоподобными клетками или даже волокнами гладкой мускулатуры.
С функциональной точки зрения капилляры являются важнейшей частью сердечно-сосудистой системы. Здесь кровь совершает свою «работу», тогда как другие отделы она проходит транзитом. В целом эндотелий образует преграду для клеток и крупных молекул, но белые кровяные тельца могут протискиваться наружу между его клетками. Мелкие молекулы и ионы свободно проходят через эндотелий, поэтому через стенки капилляров идет активная диффузия веществ. В типичном случае в своей проксимальной части капилляр отдает наружу кислород и питательные соединения, а в дистальной — получает конечные продукты обмена и двуокись углерода из тканей. Такая жизненно необходимая смена направления диффузии обеспечивается изменением баланса физического и осмотического давлений крови в процессе ее прохождения по капиллярам. Благодаря присутствию в ней специфических белков возникает тенденция к перемещению веществ из тканевой жидкости в кровяное русло. Однако в проксимальной части капилляра гидростатическое давление, создаваемое сердечным насосом, более чем уравновешивает эту тенденцию. В дистальной части гидростатическое давление снижено из-за трения жидкости о стенки капилляров, и осмотическое давление начинает перевешивать физическое.
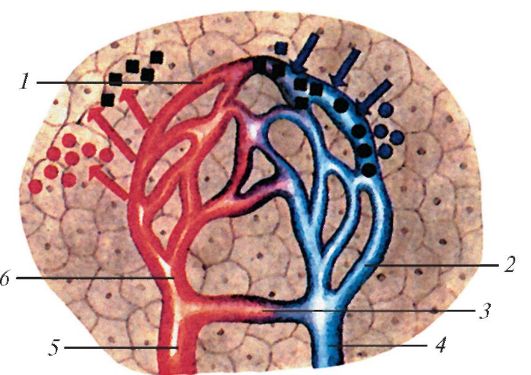
Схема строения микроциркуляторного русла: 1 — капилляры; 2 — посткапиллярная венула; 3 — артериоловенулярный анастомоз; 4 — венула; 5 — артериола; 6 — прекапиллярная артериола. Красными стрелками показано поступление в ткани питательных веществ, синими — выведение из тканей продуктов.
В тканях тела капиллярные сети обычно соединяют артериальные сосуды с венозными. В то же время разветвление на капилляры и обратное слияние последних в более крупные сосуды может местами происходить и по ходу артерий или вен. Возвращающаяся от тканей венозная кровь может также проходить через капилляры воротной системы. Кровь от кишечника проходит через воротную систему вен в капилляры печени. Воротные системы меньшего размера, но также важные функционально, встречаются и в других органах, например в гипофизе.
Несмотря на то, что основной тип артериовенозных соединений составляют капилляры, существуют и другие их типы. Так, между мелкими артериями и венами имеются прямые соединения более крупного калибра — анастомозы, которые способны регулировать поток крови через капилляры данного органа. В некоторых случаях диффузия веществ между тканями и кровью осуществляется в небольших тонкостенных емкостях неправильной формы, называемых синусоидами. Они представляют собой расширенные, часто уплощенные капилляры и встречаются в печени, селезенке и других органах. В селезенке они могут иметь неполные стенки, делая соответствующий участок кровеносной системы «незамкнутым».
Некоторые артерии (или вены) распадаются на большое число параллельно идущих и связанных анастомозами мелких сосудов, из которых кровь вновь собирается в одну артерию (вену). Такое сложное сплетение называется «чудесной сетью» (rete mirabile). Подобные структуры служат для депонирования большого количества крови с целью концентрирования в ней веществ или сохранения тепла по принципу противоточной умножающей системы. Примером «чудесной сети» может служить почечный клубочек. Другая ее разновидность присутствует в ластах многих водных млекопитающих. Лабиринт тончайших артериальных веточек окружает здесь возвращающие кровь вены. От артерий тепло передается венам и коротким путем возвращается в тело, сохраняясь ценой озябших конечностей. Кроме того, при погружении периферическое кровоснабжение часто сокращается, не меняется лишь обеспечение кровью мозга и сердца.
В хвостах многих млекопитающих (например, у кошки, собаки, крысы) находятся небольшие структуры, в состав которых также входят «чудесные сети» — хвостовые клубочки. У человека сохраняется одно из таких образований — копчиковый клубочек. Они могут представлять собой просто массу перепутанных сосудов, и, по-видимому, не используются в качестве противоточных умножающих систем. К этой же категории сосудистых сплетений относят образованные крупными сосудами целомические сети. Они встречаются у значительной части водных млекопитающих, но в то же время известны у ленивцев и некоторых приматов, поэтому трудно представить себе их функциональное значение.
Поверх обязательно присутствующего эндотелия в состав стенок всех сосудов — не только крупных (артерий и вен), но и мелких ответвлений (артериол и венул), а также, по-видимому, главных лимфатических сосудов — входят и другие компоненты. К ним относят соединительнотканные, эластические (эластиновые) и гладкие мышечные волокна в различных сочетаниях и соотношениях. Наблюдаются также нервные волокна, обслуживающие гладкую мускулатуру, а стенки крупных артерий и вен питают специальные мелкие кровеносные сосуды.
Принято считать, что стенки кровеносного сосуда состоят из трех слоев — внутренней (tunica intima), средней (tunica media) и наружной (tunica externa) оболочек. У простейших типов мелких артерий и вен первая может состоять исключительно из эндотелия, вторая — из немногочисленных мышечных волокон (в самых мелких венах последние отсутствуют), а последняя — из небольшого количества соединительной ткани. Однако у толстых оболочек крупных сосудов сложное и разнообразное строение.
В типичной крупной артерии эндотелий внутренней оболочки подстилается тонким слоем волокнистой соединительной ткани, поверх которого сосуд окружен внутренней эластической мембраной (membrana elastica interna). Средняя оболочка представляет собой преимущественно мускульный слой, хотя содержит и немного соединительной ткани; гладкие мышечные волокна иногда встречаются и в двух других оболочках. В случаях сильного развития и продольные, и поперечные мышечные волокна могут располагаться отдельными слоями. Средняя оболочка часто одета наружной эластической мембраной (membrana elastica externa). Поверх всех названных компонентов находится наружная оболочка из соединительной ткани, часто довольно рыхлой, которая прикрепляет сосуд к соседним структурам.
Строение вен приблизительно такое же, но более простое. В целом здесь гораздо ниже отношение мышечных волокон и эластической ткани к соединительной, вследствие чего стенки сосудов не столь упруги. Четкие различия между артериями и венами определяются главным образом тем, что в первых кровь находится под большим давлением. Чтобы противостоять ему, стенки у артерий толще, чем у вен, и сложнее устроены. По той же причине просвет артерии гораздо уже, чем у вены сопоставимой значимости, что связано с большей скоростью потока крови в артериях (так, например, для одинакового числа автомобилей при движении по скоростной магистрали требуется меньше рядов, чем при движении по городской улице, где скорости ниже). Диаметр артерии на всем ее протяжении остается постоянным и уменьшается только при ее раз-ветвлении. С различиями в давлении и скорости кровяного потока связано и то обстоятельство, что в артериальной системе относительно мало индивидуальных различий, тогда как вены чрезвычайно изменчивы. В конечностях артерии расположены глубоко, тогда как часть вен занимает поверхностное положение.

Схема строения стенки артерии (А) и вены (Б) мышечного типа среднего калибра: I — внутренняя оболочка: 1 — эндотелий; 2 — базальная мембрана; 3 — подэндотелиальный слой; 4 — внутренняя эластическая мембрана; II — средняя оболочка: 5 — миоциты; 6 — эластические волокна; 7 — коллагеновые волокна; III — наружная оболочка: 8 — наружная эластическая мембрана; 9 — волокнистая соединительная ткань; 10 — кровеносные сосуды.
Лимфатические сосуды служат для оттока жидкости от тканей к сердцу, т. е. по функциям они сходны с капиллярами и венами. Однако диаметр лимфатических капилляров больше, а их форма не такая правильная, как у капилляров, несущих кровь. Разумеется, лимфатические капилляры никак не соединены с артериями, а берут начало в тканях, куда проникают их слепые веточки. Через стенки капилляров путем диффузии проходит тканевая жидкость. Поскольку артериальное давление в лимфатических сосудах отсутствует, а собственное относительно низко, жидкость попадает в них без особых затруднений. Лимфа движется по сосудам очень медленно, за счет движений тела и сокращений окружающих мышц. Заметим, что у рептилий и нижестоящих классов позвоночных дополнительно имеются т. н. лимфатические сердца, снабженные клапанами и сокращающиеся за счет гладкой мускулатуры в стенках или внешних сжимающих мышц. У млекопитающих в связи с развитием сердца, сосудов и скелетных мышц необходимость в лимфатических сердцах полностью отпадает.
Лимфатические сосуды, хотя и дублируют многие функции вен и часто сопровождают их топографически, во многих отношениях отличаются от них. Стенки даже наиболее крупных сосудов настолько тонки, что сопоставимы только со стенками мельчайших вен. Внутри сосуды снабжены многочисленными клапанами, предотвращающими обратное течение лимфы. В некоторых местах сосуды могут расширяться, образуя тонкостенные резервуары — лимфатические синусы, или цистерны. На пути сосудов располагаются лимфатические узлы.
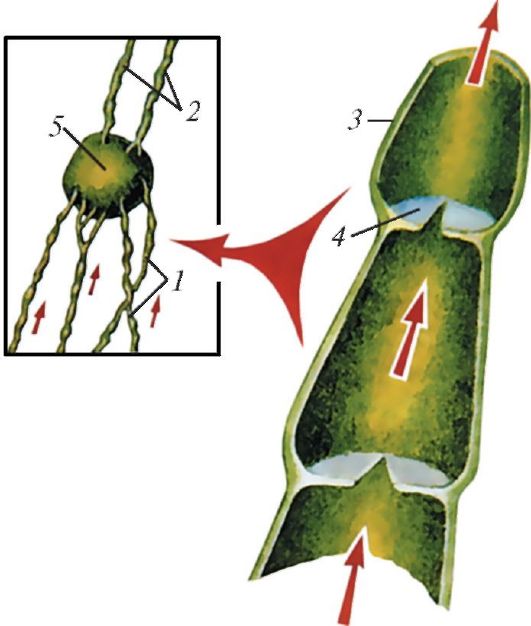
Схема строения лимфатических сосудов: 1 — приносящие лимфатические сосуды; 2 — выносящие лимфатические сосуды; 3 — стенка лимфатического сосуда; 4 — клапан; 5 — лимфатический узел.
Расширение и сужение кровеносных сосудов обеспечивается мышечной и эластической тканями в составе их стенок. Более активная регуляция потоков крови и лимфы часто осуществляется благодаря развитию сфинктеров и клапанов. В узко локализованной области небольших артерий и вен могут сильно развиваться кольцевые мышечные волокна, образующие сфинктер, способный перекрыть поток крови. Обратному течению крови или лимфы по сосуду иногда препятствует наличие внутри него клапанов. Последние представляют собой складки эндотелия и соединительной ткани с карманообразными впячиваниями сзади. Обратный ток жидкости наполняет этот карман, отгибая складку клапана в просвет сосуда. Обычно двух таких клапанов достаточно для перекрытия сосуда, однако иногда их бывает 3 или 4. Они обычны в венах, лимфатических сосудах и, конечно же, сильно развиты в сердце. В артериях клапаны почти никогда не обнаруживаются, поскольку здесь направление перемещения крови однозначно определяется высоким давлением, создаваемым сердцем.
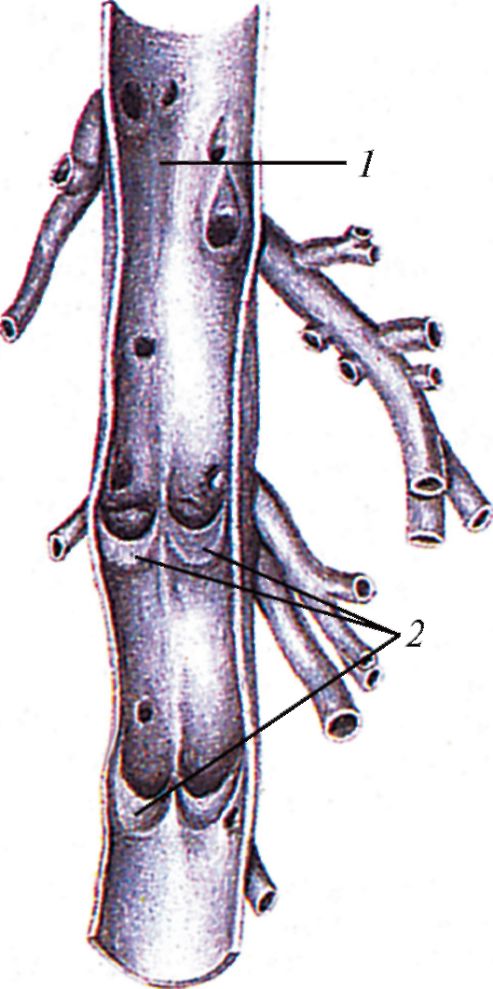
Венозные клапаны; вена разрезана вдоль и развернута: 1 — вены; 2 — створки венозных клапанов.
Артериальная система
Артериальная система млекопитающих имеет очень характерное для этой группы строение. Синапсиды обособились очень рано, и нет никаких оснований предполагать, что у них когда-нибудь имелась система из трех артериальных стволов, подобная наблюдаемой у рептилий. Полагают, что на определенной стадии у предков млекопитающих помимо легочной артерии имелся один-единственный ствол, направляющий кровь из левой части желудочка к двум сонным артериям и двум системным дугам аорты (подобная картина подтверждается особенностями эмбрионального развития). Поначалу обе дуги были вполне равноценными, но со временем правая дуга исчезла (за исключением ее основания, сохраняющегося для подачи крови в правую подключичную артерию). Причины ее утраты понятны. Дело в том, что проводимость пары сосудов хуже, чем одного ствола с суммарной площадью поперечного сечения. В двойном сосуде больше суммарная площадь стенок и потому сильнее выражено трение о них. Небольшое преимущество левой дуги, благодаря которому она сохранилась, возможно, состояло в ее эмбриональной связи через боталлов проток с легочным стволом, который у всех амниот отклонен именно налево. У рептилий левая дуга аорты несет менее оксигенированную кровь и не дает ветвей к голове и передним конечностям. Эта дуга не имеет хорошей эволюционной перспективы, и поэтому неудивительно, что у птиц, обладающих более высоким уровня метаболизма, она исчезла. Следовательно, набор системных дуг у млекопитающих и птиц упростился, но обе эти группы различаются тем, какая дуга пары сохранилась.
Итак, от толстостенного левого желудочка сердца млекопитающих отходит единственная дуга аорты (arcus aortae dorsalis), она заворачивает влево и назад, разнося артериальную кровь по всему телу. Главные артериальные сосуды ответвляются от аорты весьма изменчиво, но все варианты развиваются из одного эмбрионального состояния и функционально равноценны. Сначала от аорты отходят общие сонные артерии (arteriae сarotides), по которым кровь поступает в голову, и подключичные артерии (arteriae subclaviae), несущие кровь в передние конечности. Правая и левая общие сонные артерии могут ответвляться по отдельности, общим основанием, каждая совместно с соседней подключичной артерией и, наконец, все четыре сосуда иногда начинаются единым плечеголовным стволом. Чаще всего от дуги аорты отходит короткая безымянная артерия (arteria innominata), которая разветвляется на правую подключичную артерию (arteria subclavia dextra), правую и левую сонные артерии (arteriae carotis dextra et sinistra), левая же подключичная артерия (arteria subclavia sinistra) ответвляется от дуги самостоятельно. Реже левая сонная артерия ответвляется не от безымянной артерии, а самостоятельно от дуги аорты.
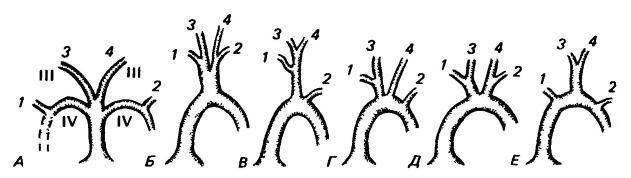
Схема, иллюстрирующая варианты ответвления основных кровеносных сосудов от дуги аорты у млекопитающих (вид снизу). А — Эмбриональное состояние — брюшная аорта с III-й (сонные артерии) и IV-й парами дуг; позднее правая IV-я дуга дальше ответвления подключичной артерии утрачивается. Различия в скорости роста сосудов обусловливают многообразие состояний (Б-Е). Состояние Г характерно для человека. 1,2 — правая и левая подключичные артерии; 3,4 — правая и левая общие сонные артерии.
Каждая общая сонная артерия делится близ головы на внутреннюю и наружную сонные артерии. У низших форм и у всех зародышей млекопитающих артериальная система головы сходна с таковой типичных амфибий и рептилий. У основания сонной артерии, там, где она еще занимает вентральное положение, отделяется небольшая наружная сонная (язычная) артерия (arteria lingualis), которая снабжает кровью ткани языка и глотки. Основной ствол сонной артерии продолжается в качестве внутренней сонной артерии вперед и вверх к той части черепа, где находится среднее ухо. Отсюда внутренняя сонная артерия направляется к отверстию в черепной коробке, но сначала в области стремечка (stapes) отделяет крупную ветвь — стапедиальную артерию (arteria stapediahs), которая снабжает кровью все структуры наружной части головы и большую часть челюстной области. Однако у большинства млекопитающих в данной картине происходят крупные изменения. Наружная сонная артерия разрастается вперед и вверх, перехватывая ветви стапедиальной и частично или полностью принимая на себя ее функции. В результате наружная сонная артерия, как правило, крупная, а стапедиальная мала или отсутствует, т. е. развивается новый, более короткий путь кровоснабжения лицевой области и челюстей. Этот процесс достигает своего наивысшего развития у кошачьих: основание внутренней сонной артерии закрывается, и кровь доставляется к мозгу преимущественно ветвями наружной сонной артерии.
Как и у других позвоночных, кровоснабжение туловища, конечностей и хвоста осуществляется продолжающей левую дугу спинной аортой и ее ветвями. Передняя часть этой аорты представлена единым сосудом, проходящим назад по средней линии тела под позвоночником и над основанием брыжейки. По своему ходу спинная аорта отдает ряд ответвлений к мускульным стенкам, конечностям и внутренним органам. Срединные вентральные (внутренностные) ответвления объединяются в несколько крупных стволов, включающих чревную артерию (arteria coeliaca), направляющуюся к желудку и печени, и переднюю и заднюю брыжеечные артерии (arteriae mesentericae), идущие к кишечнику. Среди парных вентролатеральных ответвлений короткие боковые (висцеральные) ветви снабжают кровью гонады (arteriae genitales) и почки (arteriae renales). Также имеется серия парных сосудов, отходящих от аорты вбок и снабжающих кровью осевую мускулатуру, кожу и спинной мозг. Позади спинная аорта продолжается в виде срединной хвостовой артерии (arteria caudalis).
Парные латеральные ответвления спинной аорты направляются к наружным областям тела и конечностям. Основной канал, соединяющий аорту с передней конечностью, в соответствии с его положением обычно называют подключичной артерией (arteria subclavia). Этот сосуд отдает ветви к плечевой и грудной областям, входит в конечность под названием подкрыльцовой, или подмышечной, артерии (arteria axillaris), а в конечности продолжается как плечевая артерия (arteria brachialis). Как уже говорилось, основание правой системной дуги сохраняется у млекопитающих в качестве проксимальной части правой подключичной артерии. Главным стволом задней конечности становится общая подвздошная артерия (arteria iliaca). Этот сосуд отходит спереди тазового пояса, дает ветви к некоторым органам брюшной полости, к ее стенке и затем направляется в конечность, где именуется: в бедре — бедренной артерией (arteria femoralis), в коленной области — подколенной артерией (arteria poplitea), в голени — малоберцовой артерией (arteria регоnеа).
Отдавшая кислород кровь поступает в общую легочную артерию (pulmonalis communis), отходящую, как и у птиц, от правого желудочка. Затем этот сосуд переходит на левую сторону, косо пересекая снизу основание дуги аорты, и делится на две легочные артерии (arteriae pulmonales), направляющиеся в правое и левое легкое.
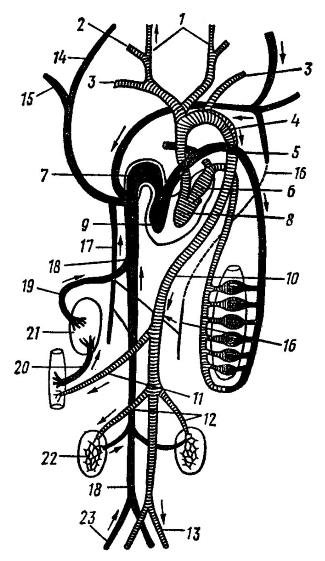
Схема кровеносной системы млекопитающих: 1 — внешняя сонная артерия (a. carotis externa); 2 — внутренняя сонная артерия (а. с. interna); 3 — подключичная артерия (a. subclavia); 4 — дуга аорты; 5 — легочная артерия (a. pulmonalis); 6 — левое предсердие; 7 — правое предсердие; 8 — левый желудочек; 9 — правый желудочек; 10 — спинная аорта (ао. dorsalis); 11 — внутренностная артерия (а. coeliaca); 12 — почечная артерия (а. геnalis); 13 — подвздошная aртерия (a. iliaca); 14 — яремная вена (v. jugularis); 15 — подключичная вена (v. subclavia); 16 — девая непарная вена (v. hemiazygos); 17 — правая непарная вена (v. azygos); 18 — задняя полая вена (v. cava posterior); 19 — печеночная вена (v. hepatica); 20 — воротная вена печени (v. portae hepatis); 21 — печень; 22 — почка; 23 — подвздошная вена (v. iliaca).
Венозная система
Расположение вен, т. е. сосудов, несущих кровь из капиллярных систем в сердце, у взрослых млекопитающих сложно и разнообразно, однако все же возможно выделить ограниченное число их систем. Прежде всего, это сложная система вен, несущих кровь к сердцу от головы, верхних частей туловища и парных конечностей. Воротная система печени направляется от желудочно-кишечного тракта к печени, а печеночные вены идут от печени к сердцу. Воротная система почек, присущая другим челюстноротым, у млекопитающих ввиду особенностей протекания их выделительных процессов полностью редуцирована. Наконец, легочные вены идут к сердцу от легких.
В увеличенной полости черепа млекопитающих развивается система сообщающихся венозных синусов. Получив кровь от мозга, венозные сосуды выходят с обеих сторон в качестве внутренних яремных вен (venae jugulares internes). Следуя назад, они сливаются с наружными яремными венами (venae jugulares externes), собирающими кровь от поверхностных частей головы и шеи. Формируются общие яремные вены (venae jugulares communes), которые сливаются с подключичными венами (venaе subclavia), отводящими кровь из передних конечностей. Их продолжения, начиная от места соединения с подключичными венами и до впадения в правое предсердие, именуются передними, или верхними, полыми венами (venae cavae anterior, или venae cavae superior). У некоторых млекопитающих (однопроходных, сумчатых, некоторых грызунов) передние полые вены открываются в сердце самостоятельно, однако чаще левая вена редуцируется и сливается с правой. В таких случаях кровь левой яремной и подключичной вен оттекает в правую переднюю полую вену по поперечному анастомозу, развившемуся впереди от сердца, — левой безымянной вене. Также характерна асимметрия остатков кардиального ствола, несущего кровь из грудной полости — непарных вен, располагающихся внутри грудной клетки по бокам от позвоночника. Как правило, только правая непарная вена (vena azygos) впадает непосредственно в правую переднюю полую вену, а левая непарная вена (vena hemiazygos) утрачивает с полой веной связь и через поперечную вену впадает в правую непарную вену.
Из задних конечностей кровь собирается в подвздошные вены (venae iliacae), которые сливаются с хвостовой веной (vena caudalis) и образуют заднюю полую вену (vena cava posterior). Обычно она представляет собой единый крупный сосуд, следующий без перерывов от области крестца и впадающий в правое предсердие. По пути задняя полая вена принимает ряд венозных сосудов от дорсальной области туловища, например, почечных вен (venae renаlis) и др., и проходит через печень, причем кровь из нее в сосуды печени не попадает. Расположение задней полой вены приурочено к средней спинной области (за исключением ехидн (Tachyglossidae), у которых она сдвинута влево). Древо ветвления этого сосуда весьма точно повторяет систему ветвей спинной аорты, особый путь имеют лишь ветви воротной системы печени.
Воротная система печени представлена венозным стволом, который собирает кровь от желудка, кишечника, селезенки и сливается в воротную вену (vena portae hepatis). Впадая в печень, воротная вена образует в ее синусоидах обширную сеть капилляров. Функциональное значение воротной системы огромно: капилляры тонкой кишки всасывают все продукты переваривания пищи (за исключением некоторых жиров), и наличие такой воротной системы гарантирует печени «право первого отбора» питательных веществ для хранения и переработки, прежде чем они поступят в общее кровяное русло. Кроме того, в печени кровь освобождается от токсинов. Пройдя через печень, кровь собирается в мощные печеночные вены (venae hepaticae), впадающие в заднюю полую вену.
Легочные вены (venaе pulmonales) несут обогащенную кислородом кровь из легких в левое предсердие. Обычно они представлены несколькими короткими сосудами (например, у крысы — тремя), впадающими в сердце со спинной стороны.
Лимфатическая система
Лимфатическая система выступает в качестве дополнения к кровеносной системе и у млекопитающих хорошо развита. Основная ее функция — возвращение из тканей в кровь белков, воды, солей, токсинов и метаболитов. Помимо этого, данная система участвует в создании иммунитета, в защите организма от болезнетворных микробов и вирусов.
Лимфа лишена цвета и обладает вязкой консистенцией. По происхождению — это межтканевая жидкость, образованная просочившейся через стенки капилляров плазмой крови и затем собранная в лимфатические сосуды. По своему химическому составу лимфа напоминает плазму, однако беднее белками. В лимфе отсутствуют эритроциты, но из примыкающих полостей в тканях сюда могут проникать лейкоциты. Обильно представлены лишь лимфоциты — фагоцитирующие клетки, очищающие лимфу от микроорганизмов и токсинов. Из лимфы они могут переходить в кровь и обратно. Количество лимфы в организме меньше количества крови в несколько раз. Ее ток происходит снизу вверх, от кончиков пальцев до грудного лимфатического протока.
Лимфатические сосуды пронизывают большинство тканей организма, за исключением центральной нервной системы, печени, хряща, зубов, кости и костного мозга. Особенно сильно они развиты в области кишечника. Сосуды переходят в протоки и стволы: слева — в грудной проток (самый большой), левый яремный и левый подключичный стволы, справа — в правый лимфатический проток, правый яремный и правый подключичный стволы. Протоки и стволы открываются в общие яремные вены, впадающие в верхнюю полую вену. Гораздо реже встречаются соединения по ходу задней полой вены. Здесь, рядом с сердцем, давление в венах самое низкое, и поступление в них дополнительной жидкости может происходить практически беспрепятственно.
От тонкой кишки по расположенным в брыжейке млечным сосудам оттекает насыщенная лимфа, называемая млечным соком, или хилусом. Эти сосуды отдают хилус в продольные, или туловищные, протоки, тянущиеся вдоль дорсальной поверхности полости тела параллельно задней полой вене и затем впадающие в нее. У некоторых млекопитающих они сохраняются в парном числе, но у большинства слиты в единый асимметричный проток. По его ходу или в его заднем конце, лежащем в поясничной области, может развиваться крупное расширение — хилезная цистерна (cisterna chyli). Значение лимфатических сосудов, контактирующих с пищеварительным трактом, огромно, ибо, если аминокислоты и углеводы поступают в воротную вену печени, то жиры транспортируются преимущественно лимфатической системой. Вероятно, это обстоятельство связано с относительно крупными размерами молекул жиров и с трудностями их проникновения в кровеносные капилляры тонкой кишки навстречу давлению.
У млекопитающих отсутствуют «лимфатические сердца» — пульсирующие участки сосудов, имеющиеся у амфибий и рептилий, поскольку при характерном для млекопитающих подвижном образе жизни движение лимфы по сосудам обеспечивается сокращениями скелетных мышц.
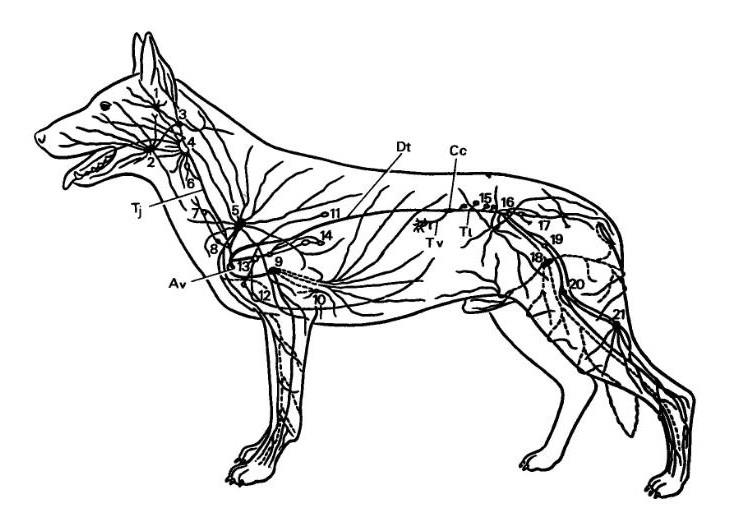
Общая схема лимфатических узлов и коллекторных лимфатических сосудов собаки (Canis lupus familiaris), без лимфатических узлов внутренних органов брюшной полости. Av — место впадения лимфы в левый венозный угол, angulus venosus; Сс — cisterna chyli; Dt — ductus thoracicus; Tj — truncus jugularis; Tl — truncus lumbalis; Tv — truncus visceralis. 1 — In. parotideus; 2 — Inn. mandibulares; 3 — In. retropharyngeus lateralis; 4 — In. retropharyngeus medialis; 5 — Inn. cervicales superficiales; 6 — In. cervicalis profundus cranialis; 7 — In. cervicalis profundus medius; 8 — In. cervicalis profundus caudalis; 9 — In. axillaris proprius; 10 — In. axillaris accessorius; 11 — In. intercostalis; 12 — In. sternalis cranialis; 13 — Inn. mediastinales craniales; 14 — Inn. bifurcationis et pulmonales; 15 — Inn. lumbales aortici; 16 — In. iliacus medialis; 17 — Inn. sacrales; 18 — Inn. inguinales superficiales; 19 — In. iliofemoralis; 20 — In. femoralis; 21 — In. popliteus superficialis.