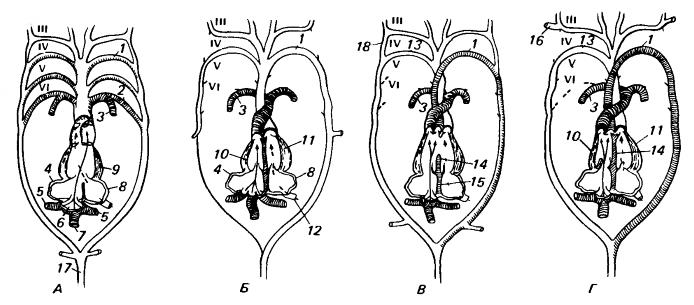
Схематическое изображение сердца и артериальных дуг у амфибии (А), млекопитающего (Б) и типичной современной рептилии (В) и крокодила (Г), вид снизу. Разрез сердца изображен таким образом, как если бы все камеры располагались в одной плоскости. Верхние концы дуг искусственно разведены в стороны Сплошные стрелки соответствуют основным потокам венозной крови, штриховые стрелки — потокам крови, приходящей от легких. Сосуды, несущие богатую кислородом кровь, оставлены незаштрихованными, а сосуды, проводящие, по-видимому, венозную кровь, заштрихованы (по сравнению с реальным положением дел, схема значительно упрощена). 2 парных сосуда в самой верхней части каждого рисунка представляют собой внутреннюю сонную артерию (латеральнее) и наружную сонную артерию (медиальнее). У амфибий, не обладающих перегородкой между желудочками, 2 потока крови до некоторой степени смешиваются, подразделение артериального конуса обусловливает тенденцию к частичному разобщению потоков, но некоторое количество венозной крови возвращается в спинную аорту. У млекопитающих (и птиц) желудочек подразделен полностью, артериальный конус преобразован в 2 отдельных сосуда, а из набора артериальных дуг сохранились лишь системная дуга и легочная артерия. Характерное для млекопитающих состояние, очевидно, сложилось непосредственно на основе примитивного, сохранившегося у амфибий, поскольку у ныне живущих рептилий наблюдается подразделение артериального конуса не на 2 сосуда, а на 3. Строение сердца на приводимой схеме упрощено. У крокодилов имеется почти полная межжелудочковая перегородка, устранение из такой системы левой IV-й дуги даст состояние, характерное для птиц. 1 — левая системная дуга; 2 — артериальный (боталлов) проток; 3 — легочная артерия; 4 — правое предсердие; 5 — передняя полая вена; 6 — венозная пазуха; 7 — задняя полая вена; 8 — левое предсердие; 9 — желудочек; 10 — правый желудочек; 11 — левый желудочек; 12 — легочная вена; 13 — правая системная дуга; 14 — межжелудочковая перегородка; 15 — перегородка предсердий; 16 — подключичная артерия; 17 — спинная аорта; 18 — часть латеральной аорты (корня спинной аорты), сохраняющаяся у некоторых рептилий.
Сердце
Сердце (cardium, или cor) является центральным органом системы крово- и лимфообращения. Оно расположено в грудной полости и снаружи заключено в околосердечную сумку — перикард (pericardium), состоящий из 2 листков, между которыми имеется щелевидная полость с небольшим количеством перикардиальной жидкости. Сердце прикрепляется к стенкам перикарда только в местах входа и выхода из него кровеносных сосудов, поэтому может легко изменять свою форму в процессе мощных сокращений. Подобно птицам, но независимо от них, сердце млекопитающих полностью разделилось на 4 камеры — относительно тонкостенные правое и левое предсердия, и толстостенные правый и левый желудочки. Каждое предсердие соединено с желудочком своей стороны атриовентрикулярным отверстием. Деятельность сердечных отделов происходит в следующей последовательности: 1) наполнение кровью и систола (сокращение) обоих предсердий, благодаря чему кровь поступает в желудочки, 2) диастола (расслабление) предсердий и систола желудочков — проталкивание крови в отходящие от сердца сосуды, 3) диастола желудочков и общая пауза сердца.
В целом сердце представляет собой усложнившееся расширение основного кровеносного ствола. Его гистологическая структура, хотя и претерпела крупные изменения, в общих чертах сравнима с наблюдаемой у других кровеносных сосудов. Здесь присутствует тонкая внутренняя выстилка — эндокард, а наружная оболочка представляет собой тонкий мезодермальный эпителий — эпикард, сходный с выстилкой полости тела. Основная масса сердца — миокард — состоит из соединительной и особой мышечной ткани. Соединительная ткань может быть плотной (особенно вокруг желудочков) и служить своеобразным скелетом. У ряда копытных в сердечной перегородке развиваются небольшие окостенения (cordis), представляющие собой опору для напряженно работающей мышцы и клапанов сердца. Сердечная мышца построена преимущественно специальными сократительными клетками — кардиомиоцитами. В полостях желудочков, а иногда и предсердий, можно обнаружить тяжи соединительной или мышечной ткани, соединяющие противоположные стенки сердца и предохраняющие его от чрезмерного расширения под действием давления.
Отверстия между сердечными камерами снабжены мощными клапанами с перепончатыми створками, дающими крови свободно проходить в одном направлении, но предотвращающими ее обратный ток. Левое предсердие отделено от левого желудочка двухстворчатым клапаном, а правое предсердие от правого желудочка — трехстворчатым. Свободные края этих клапанов прикреплены к мускульным выступам внутренних стенок желудочков посредством сухожильных тяжей, ограничивающих вдавливание клапана в закрываемое отверстие. Кроме того, клапаны обслуживаются собственными маленькими мышцами, которые сокращаются вместе со всей сердечной мышцей и обеспечивают сохранение необходимой длины соединительнотканных тяжей при изменении формы желудочка в ходе сокращения. Карманообразные полулунные клапаны имеются также в начале выходящих из сердца аорты и легочной артерии.
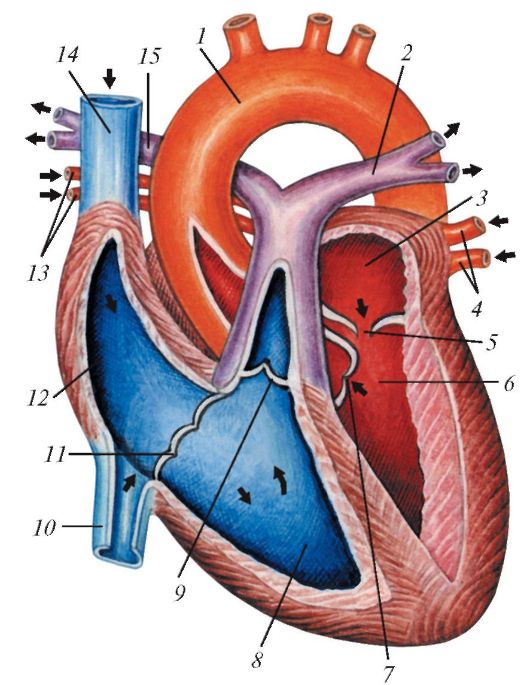
Схема строения сердца млекопитающих на примере человека, продольный (фронтальный) разрез: 1 — аорта; 2 — левая легочная артерия; 3 — левое предсердие; 4 — левые легочные вены; 5 — левое предсердно-желудочковое отверстие; 6 — левый желудочек; 7 — клапан аорты; 8 — правый желудочек; 9 — клапан легочного ствола; 10 — нижняя полая вена; 11 — правое предсердно-желудочковое отверстие; 12 — правое предсердие; 13 — правые легочные вены; 14 — верхняя полая вена; 15 — правая легочная артерия. Стрелками показано направление тока крови.
Разделение сердца на две половины исключает смешивание венозной и артериальной крови. Однако от изоляции потоков крови в сердце будет мало проку, если, покидая его, она не будет направляться в надлежащие артериальные сосуды: оксигенированная — к голове и органам тела, венозная — к легким. Большой круг кровообращения начинается артериальным стволом в левом желудочке сердца и заканчивается полыми венами, впадающими в правое предсердие. Само сердце снабжается кровью через коронарные сосуды, разветвляющиеся в его мышечных стенках. Значение этих сосудов, несмотря на их небольшие размеры, огромно. Закрытие коронарных артерий может привести к внезапной смерти от остановки сердца. Малый, или легочный, круг кровообращения начинается легочной артерией, выходящей из более тонкостенного правого желудочка, и заканчивается легочными венами, впадающими в левое предсердие.
Пока легкие не функционируют, кислород поступает в организм зародыша не через малый, а через большой круг кровообращения — через пупочную вену, идущую от плаценты к нижней полой вене, впадающей в правое предсердье. Так кровь оказывается в правой половине сердца, связанной с малым кругом. Однако находящееся на другом конце малого круга левое предсердие не получает достаточного количества крови, поскольку сюда впадают лишь легочные вены, по которым она практически не течет. В связи с этим возникает необходимость в перераспределении крови между кругами кровообращения: иначе органы, связанные с большим кругом, не получат достаточно кислорода. На помощь приходит то обстоятельство, что предсердия у зародыша соединяются овальным окном, прикрываемым специальным клапаном. Благодаря этому окну кровь из правого предсердия может поступать в левое в количестве, достаточном для нормальной работы системы. Кроме того, легочная артерия связывается с аортой через уже упомянутый артериальный, или боталлов, проток (ductus arteriosus, или ductus Botalli), который впадает в нее ниже ответвления сонных и подключичных артерий. В эти артерии смешанная кровь попадает соответственно из правого предсердия через овальное отверстие в межпредсердной перегородке, левое предсердие и левый желудочек. В момент рождения происходит резкое изменение легочного кровообращения. При первом вдохе боталлов проток рефлекторно зажимается гладкими кольцевыми мускульными волокнами и вся кровь легочной артерии проталкивается в легкие. Возвращаясь из них по легочной вене, она наполняет левое предсердие, создавая в нем высокое давление, благодаря чему прекращается поток крови из правого предсердия через овальное отверстие, оно закрывается специальным клапаном межпредсердной перегородки и в дальнейшем зарастает. Мгновенно устанавливается характерная для взрослого животного схема кровотока, но венозный проток, несущий часть крови от кишечника в обход воротной системы печени, закрывается лишь со временем.
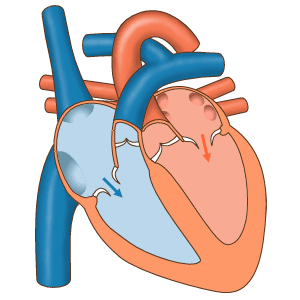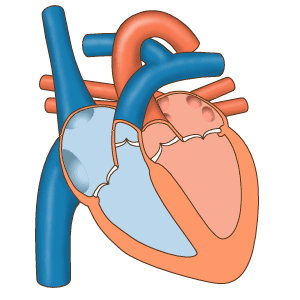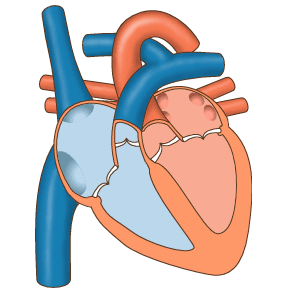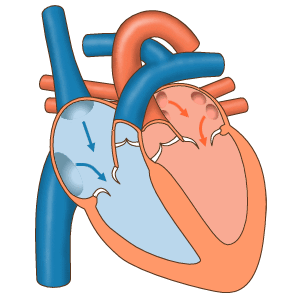
Последовательность сокращений предсердий и желудочков.
Предсердия иннервируются волокнами вегетативной нервной системы, которые способны регулировать ритм сокращений. Однако так как сердце продолжает сокращаться даже тогда, когда его жизнедеятельность поддерживается вне организма, в целом можно считать, что оно регулирует свою работу самостоятельно. Поскольку непрерывный переход мышечных волокон от предсердий к желудочкам отсутствует, для управления сердечной деятельностью развилась уникальная проводящая, или синовентрикулярная, система — фактически локальный аналог нервной. Она состоит из тяжей и скоплений особых мышечных волокон, неспособных сокращаться, но способных к генерации электрических импульсов и проведению их к кардиомиоцитам. При этом между элементами проводящей системы и кардиомиоциами имеются щелевидные контакты (нексусы), а между самими кардиомиоцитами — особые вставочные пластинки. Низкое электрическое сопротивление нексусов и вставочных пластинок обеспечивает распространение возбуждения с проводящей системы на кардиомиоциты и далее по всему миокарду. Именно способность клеток проводящей системы (водителей ритма, или пейсмейкеров) к самостоятельной генерации возбуждения и обусловливает автоматизм миокарда.
Важнейшими элементами проводящей системы являются синоатриальный узел, расположенный между местом впадения правой полой вены и ушком правого предсердия, и атриовентрикулярный узел, локализованный в нижней части межпредсердной перегородки. От атриовентрикулярного узла отходит пучок Гиса, по мере приближения к верхушке сердца распадающийся на волокна Пуркинье. Синоатриальный узел запускает сокращение, стимул передается преимущественно атриовентрикулярному узлу, откуда по пучку волокон нисходит в мускулатуру желудочков, обусловливая их сокращение.
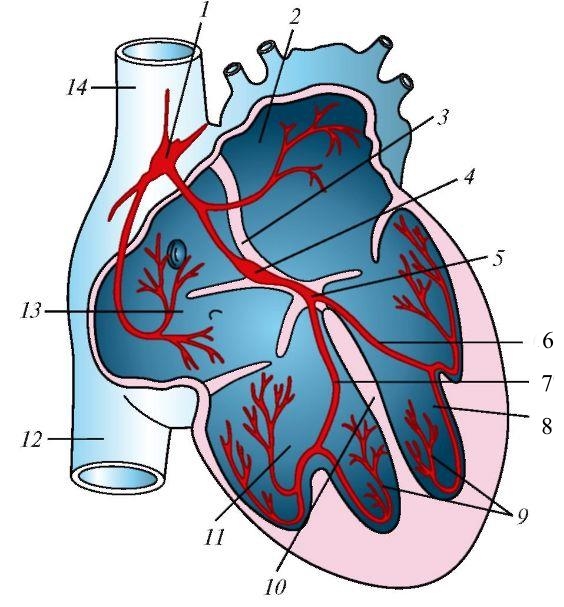
Схема проводящей системы сердца млекопитающих на примере человека: 1 — синусно-предсердный узел; 2 — левое предсердие; 3 — межпредсердная перегородка; 4 — предсердно-желудочковый узел; 5 — предсердно-желудочковый пучок; 6 — левая ножка предсердно-желудочкового пучка; 7 — правая ножка предсердно-желудочкового пучка; 8 — левый желудочек; 9 — проводящие мышечные волокна; 10 — межжелудочковая перегородка; 11 — правый желудочек; 12 — нижняя полая вена; 13 — правое предсердие; 14 — верхняя полая вена.
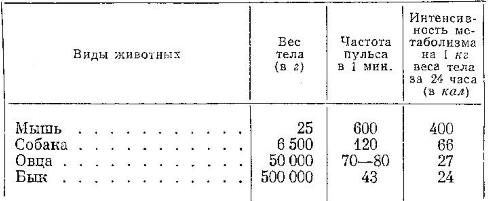
Частота сердечных сокращений (пульс) сильно варьирует, имея тенденцию к понижению у крупных и медлительных животных и повышению у мелких и подвижных. Также она зависит от возраста, физиологического состояния и характера активности животного. Так, у слона в состоянии покоя сердце сокращается приблизительно 35 раз в минуту, у коровы — 45, у овцы — 75, у собаки — 120, у кролика — 200, у ежа — 250, у крысы — 350, у мыши — 600, у летучих мышей — до 100 в покое и до 970 в полете, у мелких землероек при энергичной двигательной активности — до 1500 раз в минуту.
В последнем приведенном случае продолжительность одного сердечного цикла чрезвычайно мала и составляет всего 40-50 мс. Быстрее кровь не может проходить камеры сердца из-за своей высокой вязкости. Поэтому дальнейшее уменьшение массы тела потребует усиление кровотока для обеспечения роста интенсивности метаболизма. Есть 2 различных способа увеличить кровоток: за счет повышения частоты сердечных сокращений (а это уже невозможно) и за счет увеличения ударного (систолического) объема. Второй вариант невозможен без увеличения размеров самого сердца. А увеличение массы сердца возможно лишь при условии пропорционального изменения массы всего тела. Таким образом, у мелких млекопитающих оказываются исчерпаны все возможные способы насыщения организма кислородом. Для них характерны чрезвычайно высокая концентрация гемоглобина в крови и кислородная емкость крови. У землеройки уровень гемоглобина в крови составляет 170 г/л. У мелких летучих мышей еще выше — около 250 мг/л.
Частота пульса и интенсивность метаболизма у некоторых млекопитающих (по Наумову, 1965)
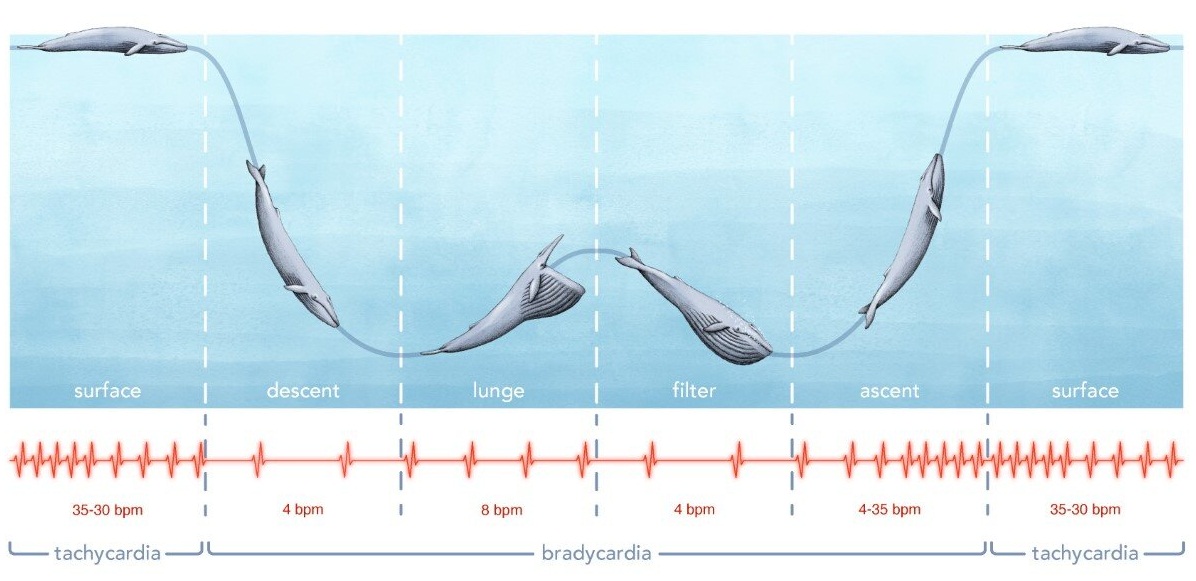
Интенсивность частоты сердцебиения имеет и иное приспособительное значение. Хорошо известно учащение пульса (тахикардия) при усилении работы мышечной системы. С другой стороны, у водных и полуводных млекопитающих при погружении в воду число сердцебиений уменьшается, возникает брадикардия, что приводит к замедлению тока крови и более экономному использованию запасов кислорода в легких и воздухоносных полостях. Например, у обыкновенного тюленя (Phoca vitulina), находящегося в непогруженном состоянии, число сокращений сердца в 1 мин равно 150. Через 11 сек после погружения оно уменьшается до 60, через 27 сек — до 35 и далее устанавливается на уровне 30 в течение всего времени нахождения зверя под водой. Одновременно изменение частоты сердцебиения облегчает выравнивание кровяного давления, которое при быстром погружении китообразных (100-140 м/мин) сильно меняется в короткие промежутки времени.
Как и в случае с легкими, относительные размеры сердца млекопитающих находятся в прямом соответствии с их образом жизни и интенсивностью обмена веществ, которые в свою очередь обусловливаются общими размерами тела, двигательной активностью и положением местообитаний над уровнем моря. Сердечный индекс (относительная масса сердца, выраженная в процентах к общей массе тела) у американского ламантина (Trichechus manatus) составляет, трехпалого ленивца (Bradypus tridactylus) и кашалота (Physeter macrocephalus) — 0,3, у саванного слона (Loxodonta africana) — 0,4, у обыкновенной полевки (Microtus arvalis) — 0,55, у обыкновенного ушана (Plecotus auritus) — 1,2-1,4, у обыкновенной бурозубки (Sorex araneus) — 1,4. В двух последних случаях масса сердца превышает ожидаемую расчетную массу в 3-4 раза, что связано с предельно интенсивным метаболизмом.
Связь между размерами тела и величиной сердца хорошо подтверждается и сопоставлением сердечного индекса систематически и экологически близких видов животных. Так, у крупного сурка-байбака (Marmota bobak) этот показатель равен 0,47, у большого суслика (Spermophilus major) — 0,61, а у малого суслика (Spermophilus pygmaeus) — 0,82. Сравним также сердечный индекс у сходных по величине мышей, перемещающихся много и быстро, и серых полевок, относительно малоподвижных: полевая мышь (Apodemus agrarius) — 0,89, европейская лесная мышь (Apodemus sylvaticus) — 0,85, полевка-экономка (Microtus oeconomus) — 0,47. У очень подвижного песца (Vulpes lagopus) отношение веса сердца к весу тела более чем вдвое превосходит таковое у относительно медлительной енотовидной собаки (Nyctereutes procyonoides). К сказанному можно добавить, что у домашнего кролика относительные размеры сердца в 3 раза меньше, чем у дикого зайца. Примерно та же закономерность выявляется и при сличении этого показателя у комнатной и охотничьей гончей собак.
Необычно сложно устроена кровеносная система жирафа (Giraffa camelopardalis) — иначе сердце попросту не смогло бы протолкнуть кровь по длинной шее животного к его голове и обеспечить кислородом и питательными веществами головной мозг. Сердце жирафа в 2,5 раза крупнее сердца животных сравнимых размеров, а давление, создаваемое им при сокращении, составляет 300 мм ртутного столба. Но когда жираф резко опускает голову к земле, давление крови в головном мозге поднимается незначительно: чтобы предотвратить резкий прилив крови к мозгу и предохранить его от возможного повреждения, ток крови в шейных артериях прерывают особые клапаны. Длинные ноги жирафа туго обтянуты плотной кожей — эти «компрессионные чулки» облегчают отток крови вверх, к сердцу животного. Кроме того, содержание эритроцитов, осуществляющих транспорт кислорода, в крови жирафа вдвое больше, чем, например, у человека.
вперед
в оглавление