в оглавление
назад
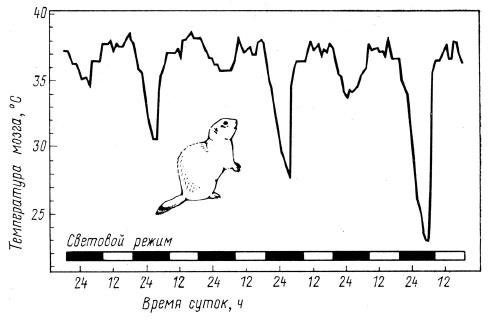
Спячка
Спячка представляет собой состояние временного торможения физиологических процессов с целью безопасно и более или менее комфортно пережить неблагоприятный в кормовом и погодном отношении период. Обычно спячка приурочивается к зиме или времени летней засухи; она практически не встречается в тропических лесах с устойчивыми запасами пищи и в тундре, где продолжительность неблагоприятного периода слишком велика. Выживание в течение многих месяцев спячки обеспечивается за счет крайне экономного использования накопленных заранее энергетических резервов. Это своеобразное приспособление свойственно только определенным группам, обладающим соответствующими экологическими и физиологическими особенностями: однопроходным (Monotremata), сумчатым (Metatheria), броненосцам (Xenarthra), тенрекообразным (Tenrecomorpha), рукокрылым (Chiroptera), мокроносым приматам (Strepsirrhini), грызунам (Rodentia), трубкозубым (Tubulidentata) и хищным (Carnivora). У части млекопитающих (сурков (Marmota), сусликов (Spermophilus), сонь (Gliridae), ежей (Erinaceidae) и некоторых других) спячка превратилась в наследственно закрепленную потребность организма, проявляющуюся в определенное время даже при наличии обстановки, вполне благоприятной для активной жизни. Нет зверей, впадающих в спячку, в тех группах, которым в наибольшей мере свойственны сезонные миграции; таковы ластоногие (Pinnipedia), китообразные (Cetacea) и копытные. Исключение из последнего правила составляют некоторые летучие мыши, у которых спячки чередуются с миграциями.
Зимняя спячка, или гибернация, распространена шире. В зависимости от глубины различается несколько ее типов.
Факультативная спячка, или зимний сон — состояние сравнительно легкого оцепенения, без затруднений прерываемое. У спящих млекопитающих примерно на треть уменьшается интенсивность дыхания и кровообращения, температура тела снижается на 2-7 °С, а общий уровень обмена веществ — на 50-70 %. Потревоженные звери легко и быстро пробуждаются. Такая спячка свойственна бурому медведю (Ursus arctos), барсуку (Meles meles), полосатому скунсу (Mephitis mephitis), еноту-полоскуну (Procyon lotor), енотовидной собаке (Nyctereutes procyonoides). У белого медведя (Ursus maritimus), как правило, в зимний сон залегают только беременные самки и неполовозрелые особи. Самки бурого и белого медведей во время спячки рожают. К данной категории можно также отнести зимнюю дремоту, в которую впадают в сильные морозы и метели белка (Sciurus vulgaris), летяга (Pteromys volans), лесная куница (Martes martes), соболь (Martes zibellina), норка (Mustela lutreola) и некоторые другие млекопитающие, по нескольку суток не покидающие свои убежища.

Свернувшись клубком — в обычной для зимней спячки позе, — суслик находится в состоянии глубокого оцепенения и ничего не чувствует, даже если его взять в руки.
Настоящая спячка, периодически прерываемая — состояние более или менее глубокого оцепенения с сохранением способности пробуждаться и короткое время бодрствовать. Интенсивность дыхания и кровообращения уменьшается более чем на половину, температура тела заметно падает, теряется реагирование на многие раздражители (звуки, прикосновения и т. п.). Такая спячка свойственна хомяку (Cricetus cricetus), бурундуку (Eutamias sibiricus), многим видам летучих мышей (особенно ушанам (Plecotus)). Животные медленно просыпаются лишь при длительных оттепелях и даже пробуют кормиться: летучие мыши ловят в пещерах оживших насекомых, хомяки и бурундуки используют сделанные в теплый период запасы корма.
Настоящая непрерывная спячка — состояние глубокого оцепенения, длящееся весь неблагоприятный сезон без перерывов. Все мышцы расслабленны, у них отсутствует тонус. Интенсивность дыхания и кровообращения уменьшается в 10-20 раз, обмен веществ — в 20-40 раз, температура тела снижается до 5-1 °С и даже ниже. Нервная система пребывает в заторможенном состоянии. Просыпания не вызывают даже самые резкие внешние раздражители (звуки, боль и т. п.) и кратковременные повышения температуры. За период спячки масса тела снижается на 30-50 %, причем в первую очередь расходуются запасы жира. Такая спячка характерна для ежей (Erinaceidae), части летучих мышей и многих грызунов (сурков (Marmota), сусликов (Spermophilus), тушканчиков (Dipodidae), сонь (Gliridae)).
Период подготовки к зиме характеризуется изменением поведения и характера обмена веществ млекопитающих. Многие из них усиленно кормятся, накопляя в организме резервные энергетические вещества (сурки, суслики, ежи). Жир откладывается в подкожной клетчатке и полости тела, аскорбиновая кислота (витамин С) резервируется в тканях, гликоген и витамин Е — в печени. Звери, не привязанные к постоянному жилищу, широко перемещаются, выбирая наиболее богатые кормом места. В средней полосе России бурый медведь (Ursus arctos) посещает ягодники и посевы овса. На хлебные поля также выходят кабаны (Sus scrofa). Повышение упитанности является важным приспособлением для перенесения зимних условий. Запасенный энергетический материал используется на поддержание основного обмена, а у беременных медведиц — еще и на развитие плода во время спячки. Кроме того, сенью происходит сезонная линька.

Суслик (Spermophilus sp.) в норе во время спячки.
Некоторые млекопитающие делают запасы кормов на время зимних пробуждений (хомяки, бурундуки). Очень важное значение имеют выбор и подготовка убежищ для спячки. В пещерах зимуют летучие мыши, в дуплах и на земле в старой листве — сони, в берлогах — некоторые медведи, в норах — различные грызуны и т. п. Как правило, температура воздуха в зимовочных убежищах не должна опускаться ниже 0 °С; однако слишком теплые убежища также не пригодны для спячки (в тепле расходование организмом резервных материалов идет настолько быстро, что они иссякают раньше, чем наступит нормальный срок пробуждения животного). Постоянство температуры и вообще микроклимата обязательно. Большинство зверей перед залеганием в спячку натаскивают в убежище мягкий растительный материал.
Особенно сложную конструкцию могут иметь зимние убежища грызунов. Так, суслики, сурки и хомяки забивают вход в зимнее жилище большой земляной пробкой, чтобы предотвратить проникновение нежданных гостей. Система ходов этих грызунов включает специальные спасательные камеры на случай затопления норы паводковыми или иными водами. Эти камеры устраиваются так, чтобы при затоплении гнездовой камеры у животных была возможность дышать и прокопать ход на поверхность земли. У сусликов в зимовальных норах устраиваются и специальные водоотводные каналы. Ошибка в выборе убежищ приводит к гибели животных. В северных и умеренных широтах нередки, например, случаи замерзания летучих мышей, оставшихся на зимовку в дуплах деревьев. В суровые зимы с необычно глубоким промерзанием почвы погибают ежи, суслики.
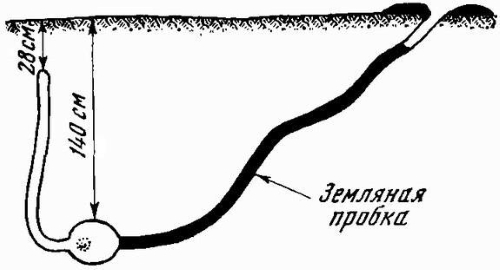
Схема зимовочной норы суслика (Spermophilus sp.) с закрывающей ее земляной пробкой.
Спят млекопитающие либо поодиночке (большинство видов), либо группами. Последние могут состоять из членов одной семьи (сурки) или из неродственных особей, нередко враждующих друг с другом в период активной жизни (сони). Возможно, что такие группы возникают постепенно путем присоединения к заснувшим уже зверькам вновь прибывших (побудительной причиной образования подобного рода групп может служить недостаток подходящих зимних убежищ). Для уменьшения излучения тепла важное значение имеет свертывание животного в клубок, поджимание под себя передних и задних лапок, прикрывание тела пушистым хвостом (сони), окутывание с помощью перепончатых крыльев (летучие мыши).
Механизм спячки обеспечивает предварительную подготовку организма и позволяет приспосабливать ее к конкретным условиям определенной местности в данном году. Он связан с рядом факторов как экзогенного порядка (температура, освещенность, наличие корма и пр.), так и эндогенных (возраст, пол, упитанность, терморегуляция, гормональная деятельность, состояние нервной системы и пр.). В зависимости от всех этих условий сроки начала спячки и ее продолжительность могут сильно колебаться даже у представителей одного и того же вида.
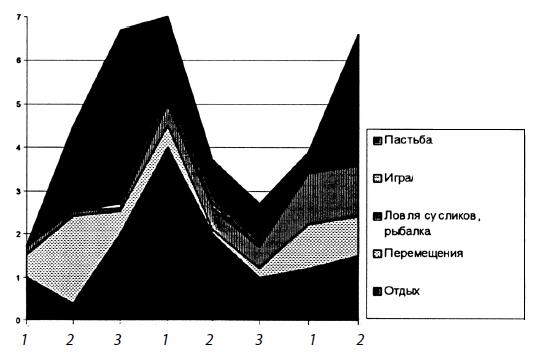
Распределение типов активности бурого медведя (Ursus arctos)в тундровой зоне Северо-Востока Сибири (1, 2, 3 — декады месяца).
В северных частях ареалов и в горах млекопитающие засыпают раньше, чем в южных и равнинных, а в районах с особенно благоприятными зимними климатическими и кормовыми условиями многие животные в спячку почти или вовсе не впадают. Так, бурый медведь (Ursus arctos) на севере Сибири лежит в берлоге с октября по начало мая, в средней полосе сон продолжается с ноября по апрель, на южных склонах Кавказского хребта он длится с декабря по конец февраля. Разбуженные в берлоге медведи могут на протяжении всей оставшейся части зимы вести бродячий образ жизни («шатуны»). В Закавказье в теплые бесснежные зимы медведи вообще не спят. Барсуки (Meles meles) на севере ареала спят с октября по май, а в Западной Европе и на Кавказе в зимний сон обычно не погружаются. Известны многочисленные случаи, когда еноты-полоскуны (Procyon lotor) и енотовидные собаки (Nyctereutes procyonoides) при длительных оттепелях выходят из дупел и нор и ведут активный образ жизни. Иногда из нор на поверхность выбираются хомяки (Cricetus cricetus). Зафиксированы их перемещения к другим норам даже по снегу на расстояния в 100-200 м при температуре до -20 °С.

На территории Воронежской области байбак (Marmota bobak) погружается в спячку во второй половине сентября, а просыпается в апреле. В среднем продолжительность спячки этого грызуна составляет 6,5 месяцев. В более северных регионах (Чувашия) байбак спит на месяц дольше, засыпая в конце августа. Суслики (Spermophilus) на склонах холмов, обращенных к северу, начинают зимовку раньше, чем на склонах южной экспозиции; взрослые самцы, рано успевающие запастись жиром, залегают первыми, за ними следуют взрослые самки и позднее всех — молодняк. Общая продолжительность зимней спячки может составить от нескольких недель до 9 месяцев. На территории России наиболее кратковременная спячка свойственна тарбаганчику (Alactagulus acontion) и некоторым другим южным формам, наиболее продолжительная — длиннохвостому суслику (Urocitellus undulatus) и черношапочному сурку (Marmota camtschatica) в северных районах их распространения.
Зависимость понижения жизнедеятельности от низкой температуры доказывается хотя бы тем, что с помощью искусственного охлаждения любое способное к зимней спячке животное может быть среди лета переведено в состояние оцепенения. Падение температуры воздуха ниже 15 °С вызывает спячку у ежа (Erinaceus europaeus) и т. п. В южных частях своего ареала некоторые звери впадают в спячку не каждый год, а лишь в особенно суровые зимы. Определенное значение имеет также изменение светового режима: сокращение светлой части суток стимулирует подготовку к спячке. Однако основным стимулом спячки, несомненно, служит ухудшение или отсутствие возможности добывания пищи — зеленых частей растений, насекомых и т. д.

Из факторов эндогенного порядка важнейшее значение имеет состояние упитанности к началу спячки; отложению жира у млекопитающих способствует снижение метаболизма, наблюдающееся у них осенью. Жир, накопляемый перед спячкой, отличается особыми физико-химическими свойствами, например низкой температурой застывания (иначе он не мог бы расходоваться при низкой температуре тела спящего животного). Можно также предполагать, что жир имеет антисептическое значение, убивая или ослабляя вредные для спящего животного микроорганизмы. Помимо обыкновенного жира, зачастую запасается т. н. бурый жир. Его главная функция состоит в образовании тепла, а не энергии для обменных процессов, для которых мобилизуются резервы обыкновенного жира. Особенно важную роль бурый жир играет в период пробуждения от спячки, когда температура тела должна быстро повыситься.
Кроме того, ряд млекопитающих обладает выраженным цирканнуальным (окологодовым) ритмом, лежащим в основе их зимней спячки. Так, у золотистого суслика (Spermophilus lateralis) и лесного сурка (Marmota monax) в условиях лабораторной изоляции при постоянной температуре, доступности корма и освещении на протяжении ряда лет сохраняется ритм зимней спячки и связанных с ней изменений живой массы. Более того, переливание сыворотки крови от спящего животного бодрствующему вызывало состояние спячки у последнего. Это говорит о наличии гуморальных факторов гибернации.
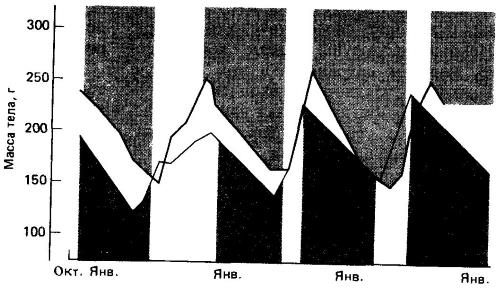
Цирканнуальные ритмы изменения массы тела в периоды зимней спячки (закрашенные области) двух особей золотистого суслика (Spermophilus lateralis), содержавшихся в неизменных лабораторных условиях.
Переселение североамериканского лесного сурка в Австралию в первое время не отразилось на цикличности зимней спячки, но впоследствии доказало наличие внутренних биологических часов, регулирующих сроки наступления зимней спячки. В первый год жизни на новом континенте животные впадали в зимнюю спячку тогда, когда в Австралии начиналась весна. Однако через 2 года сурки перестроили свою цикличность. Сезонная спячка у животных начиналась осенью по местному, а не по североамериканскому календарю. Обращает на себя внимание и тот факт, что гибернация сурков развивалась в отсутствие понижения температуры и при постоянной доступности кормовых ресурсов. Это значит, что у животных имеются врожденные механизмы гибернации на основе нервной циклической активности, запускающей гуморальные регуляторы спячки. В частях света со сменой времен года температурный фактор и фактор доступности пищи выступают своеобразными триггерами ритмической активности нейрогуморального контроля гибернации и эстивации.
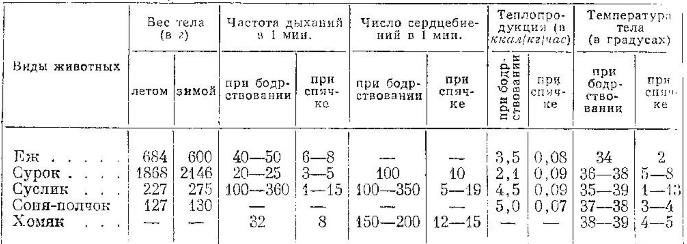
У содержащегося в неволе черного медведя (Ursus americanus), погрузившегося в зимний сон в куче сена, при температуре воздуха -8 °С и сена -7-8 °С температура на поверхности волосяного покрова составила 0 °С, на поверхности кожи +4 °С, в прямой кишке +22 °С, в ротовой полости +35 °С (против 38 °С в период бодрствования). Частота дыхания сократилась до 2-3 в минуту (против 8-14 при бодрствовании). Таким образом, в данном случае физиологические показатели уменьшились незначительно. При погружении млекопитающего в настоящую спячку у него выключается терморегуляция, вследствие чего температура тела резко снижается. Обычно она держится на уровне, лишь немного превышающем температуру окружающего воздуха. У большинства грызунов (сурки, суслики, хомяки, сони), впадающих в настоящую спячку, температура тела в летнее время равна 37-38 °С, а зимой — 0,7-7,6 °С. У ежа (Erinaceus europaeus) температура тела падает с 34-35 °С до 2-4 °С, у зимнеспящих летучих мышей — с 38 °С до 0,1 °С, а в некоторых случаях без летального исхода может опуститься ниже нуля. При постепенном отогревании переохлажденные летучие мыши возвращаются в активное состояние, если в их клетках и тканях не произошла кристаллизация воды; оживление полностью промерзшего зверька невозможно.
Физиологические показатели некоторых млекопитающих в состоянии бодрствования и спячки (по Наумову, 1965)
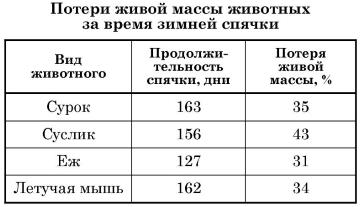
Впадающих в спячку животных называют гетеротермными. Гетеротермность служит защитным приспособлением к длительному голоданию, гибельному при сохранении обычной температуры. Например, еж (Erinaceus europaeus) в состоянии спячки остается без пищи в течение около 240 суток, бодрствующий же зверек не переносит и 30 дней голодания. Байбак (Marmota bobak) при резком снижении температуры тела в период глубокой спячки теряет в среднем около 0,1 % веса тела в сутки, голодовка же во время бодрствования вызывает у него потерю около 4 % веса в сутки.
Частота пульса и дыхания во время настоящей непрерывной спячки резко замедляется. Например, у летучих мышей количество сердцебиений падает с 420 до 15-16, а количество дыхательных движений — с 96 до 5-6 в минуту, у сурка — со 130-90 до 3-15 сердцебиений и с 20 до 3 дыханий в минуту. Вместе с тем дыхание становится нерегулярным, нарушается его ритм. Обычно вслед за серией из 5-8 дыханий наступает пауза в 4-8 или даже в 15 минут, когда животное вовсе не производит дыхательных движений. Интересно также и то, что изменяется соотношение между продолжительностью вдоха и выдоха у животных. На фоне активного дыхания (летом) вдох у животных в процессе зимней спячки значительно короче по сравнению с выдохом.
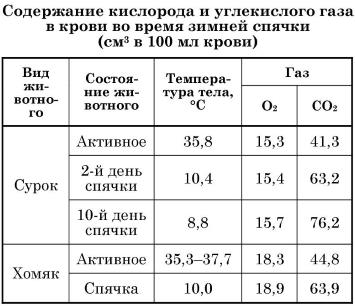
Во время спячки сокращается не только интенсивность газообмена, но и соотношение между потребляемым кислородом и выделяемым углекислым газом — т. н. дыхательный коэффициент. Например, сурки в спячке потребляют в 41 раз меньше кислорода и в выделяют в 75 раз меньше углекислого газа. Для спящего сурка обычным является падение дыхательного коэффициента с 0,8 до 0,5-0,4. Подобное явление описано и для других зимнеспящих млекопитающих. У сони дыхательный коэффициент падает до рекордной отметки 0,23.
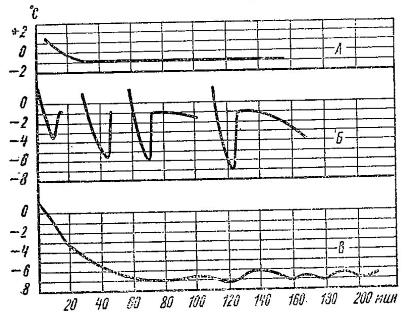
Изменения температуры тела рыжих вечерниц (Nyctalus noctula) при охлаждении ниже 0 °С: А — замерзание без переохлаждения; Б — замерзание после переохлаждения; В — длительное переохлаждение.
Изменения, происходящие в организме животных во время зимней спячки, отражаются на химическом составе и физических свойствах их крови. Снижение температуры тела повышает растворимость газов в крови спящих животных. Если кислородная емкость крови у животных в активном состоянии и во время спячки примерно одинакова, то концентрация углекислого газа в ней существенно возрастает. Накопление углекислого газа в крови животных в спячке относят к изменению функционального состояния легких и снижению активности карбоангидразы, которая катализирует процесс диссоциации растворенной в крови угольной кислоты и последующую диссоциацию гидрокарбоната натрия. Снижению интенсивности газообмена спящих животных способствует и накопление углекислого газа в закупоренных землей норах, где его концентрация за время спячки многократно возрастает. В результате повышенного содержания углекислого газа в дыхательном воздухе возрастает его содержание и в альвеолярном воздухе. А это резко повышает парциальное давление углекислого газа, и основной механизм газообмена между легкими и венозным кровотоком (разница парциального давления) блокируется. Поэтому и возрастает концентрация углекислого газа в крови. Здесь уместно отметить и то, что у спящих животных чувствительность дыхательного центра к углекислому газу чрезвычайно низка. При зимней спячке запредельная для активных животных его концентрация в крови не вызывает одышки.
С другой стороны, потребность животных в кислороде во время зимней спячки существенно понижена. Летучие мыши и ежи в бодрствующем состоянии, будучи опущены в воду, задыхались спустя 2-3 минуты. Спящие летучие мыши пребывали без всякого вреда под водой до 16 минут, а спящий еж ожил после пребывания под водой в течение 29 минут. У убитого во время спячки сурка сердце продолжает сокращаться несколько часов, в то время как у убитого бодрствовавшего зверька оно останавливается через несколько минут.
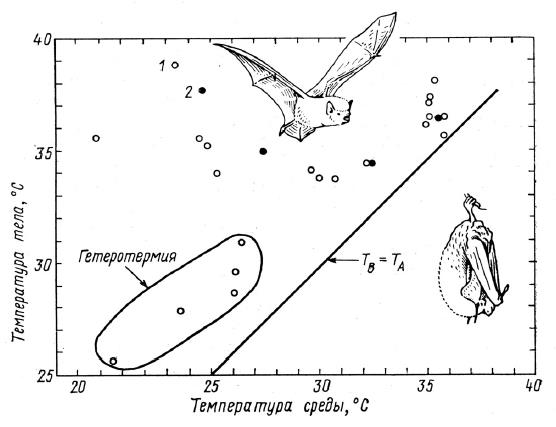
Зависимость температуры тела (Тв)от температуры среды (Та) у активных и находящихся в оцепенении летучих мышей: 1 — белобрюхий трубконосый крылан (Nyctimene albiventer); 2 — Paranyctime rasptor.
Спячка сопровождается снижением концентрации глюкозы в крови животных. Так, при температуре тела сурков на уровне 35-36 °С (активное состояние) концентрация сахара в их крови лежит у разных особей в пределах 117-214 мг%. При понижении температуры тела до 10 °С (спячка) концентрация сахара падает до 10-70 мг%. У суслика во время спячки уровень глюкозы падает до 24-89 мг% против 150-200 мг% у активных животных. У бурундуков перед спячкой сахар присутствует в количестве 150-180 мг%, а в середине спячки (декабрь) этот показатель крови падает до 60-75 мг%, т. е. в 2,5-3 раза. Однако обнаружены и прямо противоположные состояния углеводного обмена у животных во время спячки.
Давно замечено, что в процессе подготовки к спячке и в процессе ее у животных увеличиваются запасы углеводов в органах и тканях. У сурка в летнее время концентрация гликогена (животного крахмала) в печени очень низкая — от 0 до 20 мг%. Но уже на 4-й день спячки она увеличивается многократно и составляет 600 мг%. На 7-й день зимней спячки в печени сурка обнаруживали 800-900 мг% гликогена, а на 10-й — уже 1635 мг%. Этому явлению можно предложить два биохимических объяснения. Спячка включает механизм активизации процесса глюконеогенеза, т. е. образование глюкозы и ее резервной формы гликогена из жира. Возможно и другое. Накопление гликогена является результатом снижения потребности организма в глюкозе во время спячки и переключения метаболизма животных на использование жиров в качестве основных источников энергии. Очевидно, что низкое содержание сахара в крови животных в период спячки достаточно для обеспечения центральной нервной системы.
Энергетические потребности висцеральных органов и опорно-двигательного аппарата обеспечиваются не за счет углеводного, а за счет жирового обмена. На юге России суслики (Spermophilus) за лето прибавляют в весе на 80-100 г за счет жироотложений. У байбака (Marmotа bobak) подкожный и внутренний жир в июне составляет 10-15 г, в июле 250-300 г, в августе 750-800 г. У некоторых особей жир составляет до 25 % общей массы тела. Соня-полчок (Glis glis) к концу лета полнеет настолько, что количество жира равно 20 % ее общего веса, а орешниковая соня (Muscardinus avellanarius) за счет жироотложений увеличивает свою живую массу с 15 до 35 г. Вес ежа (Erinaceus europaeus) за лето увеличивается в 2,5 раза. Тарбаганчик (Pygeretmus pumilio) за счет жироотложений увеличивает массу своего тела на 30-50 %. Енотовидная собака (Nyctereutes procyonoides) летом весит 4-6 кг, зимой — 6-10 кг. Интересно, что довольно часто животные в начале спячки продолжают прибавлять в весе, не питаясь при этом. Так, у сурков (Marmotа) в первые 3-5 дней зимней спячки наблюдают увеличение веса тела на 0,10-0,15 %.
Местом отложения жира у животных перед спячкой является подкожная клетчатка, где толщина его отложений, например, у сурка, достигает нескольких сантиметров. Однако начинается жироотложение с внутренних органов. Летом при хороших кормовых условиях жир откладывается в сальнике и вокруг некоторых органов, образуя капсулу (вокруг почек, яичников, сердца). У впадающих в спячку млекопитающих имеется и специфическое депо жира — это т. н. железа спячки. Данное образование находится в грудной полости у позвоночника. У голодающих активных животных она представлена соединительнотканной губчатой массой, которая по мере приближения осени заполняется жиром.
В процессе спячки млекопитающие ежедневно расходуют за счет жироотложений от 0,1 до 0,4 % живой массы. Однако уровень использования жира зависит от активности обмена веществ. В условиях контролируемого эксперимента сурки при голодании в активном состоянии теряли 3-4% живой массы в сутки. В начале зимней спячки эти потери снизились до 0,24-0,35 %. По данным разных авторов, ежесуточные потери живой массы во время спячки у сусликов составляют 0,18-0,48 %, у ежа — 0,24 %. Из спячки животные, как правило, выходят, имея некоторый запас внутреннего жира. Он позволяет животным избежать резкого перехода на активное питание, поскольку смена источников энергии для обеспечения метаболизма требует некоторого времени. Однако если упитанность животного осенью была недостаточной, то возможно и раннее пробуждение от спячки. Так случается, например, с бурыми медведями (Ursus arctos).
Помимо жировых отложений, организм зимоспящего животного использует ткани других органов. У сурка расход тканей выглядит следующим образом: жировая ткань — 99 %; печень — 59 %, диафрагма — 46 %; легкие — 45 %, мышцы — 30 %, сердце — 27 %, скелет — 12 %. Но не во всех случаях расход бывает таким большим. Как уже говорилось, в печени во время спячки происходит накопление гликогена, что может привести в начале ее к некоторому увеличению веса тела. Прибавление в весе может происходить и от накопления метаболической воды в организме. Конечно, в дальнейшем по мере расходования резервных материалов происходит потеря в весе.
Интересное явление фиксируется у тринадцатиполосого суслика (Ictidomys tridecemlineatus). Как и в других случаях, у погрузившихся в глубокую спячку зверьков дыхание и сердцебиение значительно замедляются, а температура тела падает. Примерно раз в 10 дней она снова повышается до нормальных значений часов на 12, но животное при этом продолжает спать. Масса суслика во время спячки неуклонно падает, и к апрелю он теряет до 40 % своего веса. Примечательно, однако, что примерно за 2 месяца до выхода из спячки мышцы конечностей прекращают дегенерировать, а затем даже увеличивают свой объем. Молекулярные маркеры выявили в этот период повышенную продукцию белков скелетной мускулатуры — на уровне, характерном для летнего времени. Благодаря этому явлению зверек пробуждается готовым активно добывать еду и убегать от хищников.
Наряду с описанными наблюдаются и другие более или менее глубокие изменения физиологического состояния погруженного в спячку животного: например, деятельности эндокринных желез (ослабление их функций вследствие инволюции; ясно выраженная гипофункция щитовидной железы во время спячки наблюдается, например, у ежей, летучих мышей, сурков, которые при введении тиреоидина пробуждаются; с другой стороны, инкреторная функция поджелудочной железы во время сильно возрастает), биохимических процессов, протекающих в мозгу, в состоянии нервной системы вообще (торможение функций коры больших полушарий, снижение тонуса симпатической системы и пр.) и, наконец, в состоянии половой сферы (например, подавление сперматогенеза у ежей и некоторых других спящих зимой животных, в то же время у самцов сурков, сусликов, бурундуков и некоторых других активность половых желез во время спячки полностью не прекращается, и с момента пробуждения эти зверьки способны участвовать в гоне).
Несмотря на резкое общее снижение жизнедеятельности, организм животного во время спячки все же сохраняет способность реагировать на отдельные внутренние и внешние раздражения. Например, чрезмерное накопление углекислоты в крови, переполнение мочевого пузыря, повышение температуры в убежище могут вызвать пробуждение животного. Тот же эффект наблюдается при охлаждении животного ниже свойственной ему минимальной температуры спячки, так как в таком случае включается в действие терморегуляция, а вместе с тем повышаются до предела активность и температура тела. Пробуждение может быть кратковременным, при котором животное освобождается от накопившихся продуктов обмена веществ, согревается и затем снова засыпает, или полным. В ряде случаев оно может иметь гибельные для животного последствия. С другой стороны, у некоторых видов зимнее оцепенение достигает такой глубины, что почти полностью исключает реагирование животного даже на сильные внешние воздействия (у суслика, например, спячка не прекращается при нанесении ему серьезных ранений вплоть до ампутации конечностей).
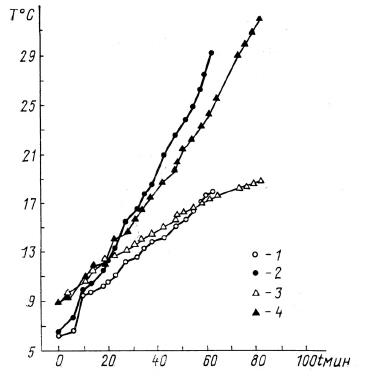
Повышение температуры тела у пробуждающихся из спячки грызунов при температуре воздуха 18-20 °С: 1, 2 — суслик; 3, 4 — хомяк; 2, 4 — температура в пищеводе; 1, 3 — в прямой кишке.
Весеннее пробуждение от спячки во многих случаях вызывается стойким повышением температуры среды. Сроки этого явления изменяются в зависимости от тех же причин, что и сроки залегания, но последовательность явлений, конечно, обратная (на юге пробуждение начинается раньше, чем на севере, и т. д.). В момент пробуждения животное производит конвульсивные движения, температура его тела быстро повышается, находившиеся в угнетенном состоянии физиологические процессы восстанавливаются. Вследствие хотя и крайне замедленного, но не прекращающегося на протяжении всего периода спячки расходования резервных материалов, животные к весне теряют часть веса. Однако основные резервы они используют в голодное весеннее время, у ряда видов совпадающее с брачным периодом. Виды, впадающие в зимний сон, гораздо менее экономно расходуют резервные вещества и, как правило, пробуждаются истощенными. Они усиленно кормятся, восстанавливая подорванные длительной голодовкой силы.
Помимо большого теоретического интереса, явление зимней спячки имеет и практическое значение, поскольку среди впадающих в спячку млекопитающих имеются промысловые виды, вредители сельского хозяйства, переносчики инфекций. Изучение явлений спячки открывает также некоторые перспективы лечебного применения искусственно вызванного (охлаждением) оцепенения. Экспериментально установлено, что на протяжении всего периода спячки у млекопитающих не развиваются инфекционные болезни, даже такие смертоносные как чума; подавляется также рост злокачественных опухолей. Путем охлаждения удавалось полностью освобождать организм некоторых животных от болезнетворных простейших (трипаносом, спирохет). Холодовое оцепенение применяется и в медицинской практике (при сложных операциях и т. п.).
Настоящая летняя спячка, или эстивация, свойственна некоторым пустынным и степным млекопитающим. Ее общей причиной служит высыхание растительности, приводящее к невозможности получить вместе с кормом количество воды, необходимое для нормального функционирования организма. Дополнительно развитию спячки способствует изменение нейрогуморального баланса под влиянием слишком высокой температуры окружающей среды.
«Пробные» понижения температуры тела у сусликов в период, предшествующий впадению в спячку.
Наиболее полно летняя спячка изучена у сусликов (Spermophilus). По мере высыхания растений их роющая деятельность повышается, они устраивают для спячки норы, в которых поддерживается умеренная температура и оптимальный уровень влажности. Зверьки принимают обычную позу сна и впадают в оцепенение, при котором температура тела падает и происходит общее снижение физиологической активности. При этом в годы с влажным дождливым летом переход в спячку происходит значительно позднее, чем в засушливые годы. Желтый суслик (Spermophilus fulvus) залегает в летнюю спячку, как только содержание влаги в траве снижается до 5-18 %. В пустынях Средней Азии это происходит уже в конце мая - начале июня, а в северной части его ареала данный срок отодвигается на июль - август. Продление периода активности наблюдается также у зверьков, поселяющихся возле арыков, где свежая растительность сохраняется значительно дольше. Обычно летняя спячка без перерыва переходит в зимнюю, и желтый суслик становится активными лишь следующей весной, проведя в оцепенении 6-8 месяцев. Малый суслик (Spermophilus pygmaeus) в полупустынных областях впадает в спячку в июле. У такого относительно северного вида, как крапчатый суслик (Spermophilus suslicus), это происходит в августе.
Имеются указания, что в летнюю спячку также впадают некоторые виды сурков (Marmota) и орешниковая соня (Muscardinus avellanarius). С исчезновением насекомых в жаркие месяцы связана непродолжительная летняя спячка у ряда летучих мышей. Белобрюхий еж (Atelerix albiventris) проводит в летней спячке около 3 месяцев, обыкновенный тенрек (Tenrec ecaudatus) — порядка 4 месяцев. Насколько известно, из приматов в спячку погружаются лишь крысиные лемуры (Cheirogaleus). Сигналом для этого является повышение температуры воздуха до 30 °С. При этом лемуры Кроссли (Cheirogaleus crossleyi) и Сибри (Cheirogaleus sibreei) зарываются в мягкую подстилку тропического леса, где поддерживается постоянная температура, а толстохвостый лемур (Cheirogaleus medius) укрывается в расположенном на сквозняке дупле, и температура его тела меняется в соответствии с температурой воздуха. В спячку лемуры проводят 6-7 месяцев, пережидая период засухи и недостатка плодов.
Перед впадением в летнюю спячку в организме животных накапливаются резервные вещества (в том числе и жир). Например, малый суслик (Spermophilus pygmaeus) весной весит 90-120 г, а к середине лета — 400-500 г. У толстохвостого лемура (Cheirogaleus medius) масса тела перед спячкой увеличивается на 40 %. Существенное отличие летней спячки от зимней заключается в том, что она протекает при температуре среды, близкой к температуре бодрствования. Температура тела спящего животного в большинстве случаев также остается относительно высокой, вследствие чего обмен веществ в организме происходит более интенсивно, чем во время зимней спячки. Однако некоторые физиологические процессы все же замедляются. Так, сердечный ритм во время спячки может снижаться до 2-3 ударов в минуту, дыхательный ритм урезается до 1 цикла в 3 мин, почти полностью затухает электрическая активность мозга, хотя восприимчивость к некоторым внешним стимулам может сохраняться. В общем же летняя спячка менее глубока, чем зимняя.
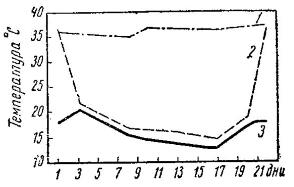
Изменения температуры тела малого суслика (Spermophilus pygmaeus) в летней спячке: 1 — температура тела бодрствующего суслика; 2 — температура тела спящего суслика; 3 — температура воздуха.
Как зимняя, так и летняя спячка может рассматриваться в качестве сложнорефлекторной реакции на изменения условий среды, как биологически целесообразное и наследственно закрепленное свойство, исторически выработавшееся у некоторых видов млекопитающих, дающее им возможность противостоять неблагоприятным сезонным явлениям природы и существовать в областях, где круглогодичная деятельность для них совершенно невозможна.
вперед
в оглавление