в оглавление
назад
Суточная активность
Днем и ночью большинство животных сталкивается с разными условиями. Приспосабливаясь к ним, животные производят суточное распределение активности, представленной многими видами поведения, которые в совокупности образуют определенный распорядок, повторяющийся с небольшими отклонениями изо дня в день. Можно легко заметить, что каждый вид активности имеет тенденцию следовать типичному циркадному, или суточному, ритму.
Самые важные суточные изменения во внешней среде — это изменения температуры и освещенности. В холодном климате мелким млекопитающим может быть выгодна ночная активность при низкой температуре среды. Период наибольшей теплопродукции придется у них в таком случае на самую холодную часть 24-часового цикла, когда активность способствует терморегуляции. В то же время подавляющее большинство мелких птиц холодными ночами сберегают энергию, становясь неактивными и давая температуре тела снизиться. В жарком климате мелким млекопитающим также выгодно быть ночными животными, избегая дневной жары.
По отношению к свету животные могут быть разделены на ночных (включая сумеречных), дневных и нейтральных — т. е. активных как днем, так и ночью. Виды, специализированные к дневному зрению, ночью могут оказаться в неблагоприятном положении, потому что легко станут жертвой хищников и не смогут эффективно добывать себе пищу. Впрочем, даже у отдельно взятой особи отношение к свету существенно изменяется под влиянием различных внешних и внутренних причин (голода, степени преследования, кормовых условий и др.). Несмотря на врожденную природу характера активности, она может терять свою четкую временную приуроченность в различные сезоны года, в разные периоды жизненного цикла животных, в частности в сезон размножения.
В зависимости от количества сменяемых друг друга периодов деятельного состояния и покоя суточную активность разделяют на несколько типов. Монофазная активность, которую графически можно изобразить в виде одновершинной кривой, свойственна, например, барсуку (Meles meles), хохлатому дикобразу (Hystrix cristata) и шимпанзе (Pan troglodytes). Двухфазная активность (выглядит как двухвершинная кривая) хорошо выражена у некоторых видов летучих мышей (Microchiroptera), сусликов (Spermophilus), сурков (Marmota). Полифазная активность (в виде многовершинной кривой) обычна для землероек (Soricidae), кошек (Felidae), енотовидной собаки (Nyctereutes procyonoides) и полевок (Arvicolinae).
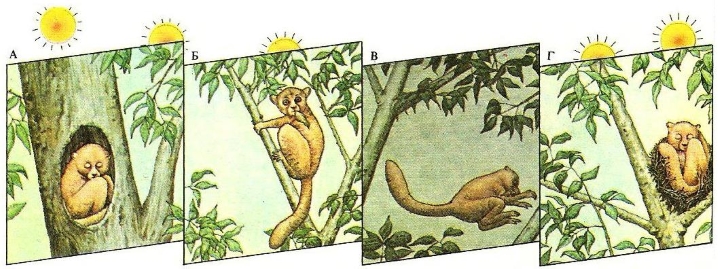
Монофазная суточная активность ласковидного лемура (Lepilemur mustelinus). Днем зверек спит в древесном дупле или гнезде из листьев и веток (А), а в сумерках выходит кормиться корой и древесной листвой (Б). Ночью он активно передвигается по своему индивидуальному участку (В) в радиусе 50 м от гнезда. С рассветом возвращается «домой» (Г).
Неудивительно, что ритмы покоя и активности распространены в животном мире так широко. Если ночная активность невыгодна, лучше всего сидеть неподвижно в надежном месте и сохранять как можно больше энергии. Было высказано предположение, что это одна из главных функций сна. Ночные виды могут затаиваться днем, если им грозит нападение. Если же они сами являются ночными хищниками, то днем могут прятаться и бездействовать, чтобы не спугнуть добычу. Таким образом, суточные ритмы физической внешней среды делают некоторые виды активности выгодными в одно время и невыгодными в другое.
В зависимости от своей экологии, животные придерживаются такого суточного распорядка, который максимально увеличивает ценность разных видов его активности для выживания. В дополнение к типичному для вида суточному распределению активности отдельные особи могут приобретать свои собственные привычки. Например, хищник, обнаруживший добычу в определенное время и на определенном месте, стремится повторить на следующий день тот же охотничий маршрут. Такая стратегия годится в тех случаях, когда виды-жертвы тоже имеют свой собственный типичный суточный распорядок.
Ритмика активности у млекопитающих подвержена географическим изменениям. В полярных странах, где день длится на протяжении почти всего лета, а ночь — в течение всей зимы, преобладают нейтральные формы с полифазным типом активности (ночные виды здесь легко приспособляются к полярному дню, дневные — к полярной ночи, что до некоторой степени сглаживает разницу между ними). В остальной части зоны арктических пустынь и тундровой и в умеренном поясе повышается число дневных форм, а в субтропиках и тропиках преобладают ночные формы.
Имеются доказательства межвидового социального влияния на ритмику поведения. Так, кабан (Sus scrofa) в природных безопасных условиях является дневным животным. В непосредственной близости от жилья человека кабан имеет пик активности в темное время. Подобным образом меняется активность лисицы (Vulpes vulpes). В московском парке Сокольники лисицы активизируются (появляются около мусорных ящиков) в сумерках и в ночное время, когда отдыхающие покидают парк. Близость человека делает ночными животными и таких грызунов, как мыши (Mus) и крысы (Rattus).
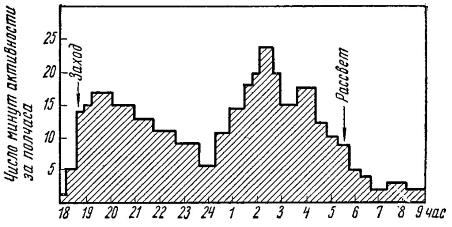
Двухфазная суточная активность лесной мыши (Apodemus silvaticus).
На активность многих зверей оказывает влияние яркость освещения. В пасмурную погоду, например, независимо от температуры, утренний выход сурков (Marmota), сусликов (Spermophilus) и других дневных норников на поверхность значительно запаздывает, общая активность их снижается, а вечерний уход в норы ускоряется по сравнению с солнечными днями.
Ночные млекопитающие реагируют на свет Луны. Причем их ответная реакция может быть прямо противоположной. Так, у полевок и летучих мышей яркий лунный свет угнетает активность. Мирикины (Aotus), домашняя кошка (Felis silvestris catus), наоборот, при яркой Луне усиливают свою активность.
Некоторые виды связывают свою ночную активность не с яркостью лунного света, а с фазами луны. Например, знамехвостый (Dipodomys spectabilis) и фресновский (Dipodomys nitratoides) кенгуровые прыгуны проявляют повышенную активность в новолуние, а наименьшую — в полнолуние.
Привязанность к фазам Луны обнаружили у ряда рукокрылых. Так, ямайский листонос (Artibeus jamaicensis) демонстрирует повышенную пищевую активность в начале ночи в определенный период лунного месяца, а именно, в интервале от последней до первой четверти лунного цикла. В другую половину лунного месяца (первая - последняя четверти) животное вылетает на кормежку перед рассветом. Крыланы (Pteropodidae) синхронизируют свою активность с фазами лунного месяца даже тогда, когда небо покрыто плотным слоем облаков и Луны на небе не видно. Это говорит о том, что для распределения фуражировки во времени эти животные пользуются внутренними «лунными часами».
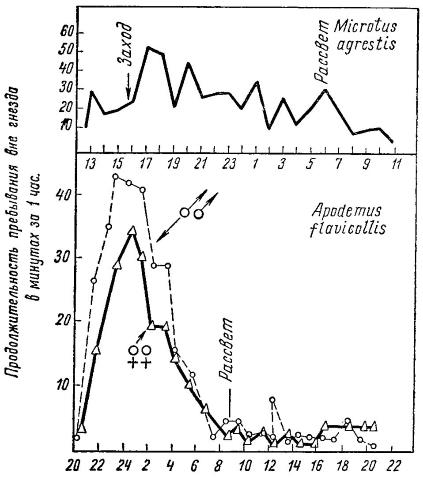
Полифазная суточная активность темной полевки (Microtus agrestis) и монофазная — желтогорлой мыши (Apodemus flavicollis).
Не меньшее влияние на ритм активности млекопитающих может оказывать температура. Летом, в часы наиболее высокой температуры наблюдается резкое снижение деятельности большинства млекопитающих; они укрываются в норах и т. п. убежищах, что предохраняет их от перегрева. Весной, осенью и зимой эти же животные могут оставаться деятельными всю светлую часть суток. Хорошо выражена эта закономерность у джейрана (Gazella subgutturosa), в летний период пасущегося два раза в сутки (утром и вечером), а зимой — на протяжении целого дня (конечно, с перерывами на отдых). Кривая суточной ритмики сурков (Marmota), сусликов (Spermophilus), большой песчанки (Rhombomys opimus) летом двухвершинна, весной и осенью — одновершинна. Такое же соотношение летней и зимней активности отмечено для полевки Брандта (Microtus brandti) и ряда других видов. Выпадение осадков, сильные ветры обычно снижают активность млекопитающих. Во всех перечисленных явлениях ясно выражена зависимость суточной активности от метеорологических условий.
Связь ритмики активности с типом питания отчетливей всего выражена у ночных и круглосуточно активных видов. Многофазность суточной активности ряда мышевидных грызунов, землероек и некоторых других млекопитающих обусловлена тем, что они вынуждены несколько раз наполнять желудок пищей; обеспечить себе суточную кормовую норму за один прием они не могут. Европейская лесная (Apodemus sylvaticus), желтогорлая (Apodemus flavicollis), японская лесная (Apodemus speciosus) и полевая (Apodemus agrarius) мыши деятельны преимущественно ночью и могут в дневное время не выходить из гнезда в течение 2-8 часов. Такие продолжительные перерывы поиска и поедания корма среди мелких грызунов свойственны только видам, питающимся высококалорийной пищей — семенами, орехами, насекомыми, насыщающими зверьков на достаточно значительный срок.

Многим млекопитающим свойственна большая пластичность и лабильность ритма активности, что позволяет им приспосабливаться к меняющимся условиям среды. Синантропные формы, например, легко «подгоняют» свойственную им суточную ритмику к многообразию режимов, существующих в поселениях человека. При усиленном преследовании охотниками дневные промысловые звери иногда переходят на ночной образ жизни. Так, например, дневной по своей природе кролик (Oryctolagus cuniculus) во многих местах, где на него интенсивно охотятся, превратился уже в ночное животное. Наоборот, отсутствие опасности, например в малолюдных районах или заповедниках, приводит иногда к восстановлению дневной активности у очень осторожных зверей — бобра (Castor fiber), барсука (Meles meles), рыси (Lynx lynx). У лисицы (Vulpes vulpes), енотовидной собаки (Nyctereutes procyonoides), соболя (Martes zibellina) в зависимости от обилия корма и интенсивности преследования преобладает то ночной, то дневной тип активности.
Высокая плотность популяции также может вызвать изменение характера активности. Например, сумеречные летучие мыши при недостатке пищи частично переходят на ночное питание, а ночные — на утреннее и дневное (такие зверьки иногда летают даже при ярком солнечном свете). Мало времени на отдых животные тратят в период миграции. У копытных наблюдаются нарушения ритма суточной активности при массовом появлении кровососущих двукрылых (гнуса). Как правило, в зонах с неблагоприятными условиями питания суточная активность млекопитающих повышается, в оптимальных — понижается.
Помимо внешних факторов, суточная активность существенно корректируется физиологическим состоянием самих млекопитающих. Так, животные, испытывающие жажду или голод, имеют повышенную мотивировку поискового поведения. Резко повышается активность самцов и самок ряда млекопитающих в период течки и, наоборот; у беременных самок и больных животных она резко падает. В разгар гона ночные и дневные формы часто становятся деятельными круглосуточно. Наоборот, роль отдыха особенно велика в начале и конце жизни. Новорожденные (прежде всего у незрелорождаемых видов) спят свыше 20 часов в сутки. У старых животных время отдыха также увеличено.
Поведенческая динамика фиксируется по двигательной (мышечной) активности, которую обслуживают вегетативные процессы. Поэтому можно ожидать, что циркадные ритмы активности характерны не только для опорно-двигательного аппарата, но и для работы вегетативных органов. Среди вегетативных реакций на свет выделяется температура тела. У человека суточные колебания температуры тела составляют 1° С, у обезьян — 5° С, а у летучих мышей — до 10 °С. У человека и животных описана и суточная динамика изменений в работе сердца, артериального давления крови, частоты дыхания, интенсивности диуреза, а также суточная динамика в изменениях состава мочи и крови. Однако у многих видов животных, ведущих преимущественно дневной образ жизни, смена дня и ночи не оказывает заметного влияния на поведение и на вегетативные функции (собака, лошадь). У грызунов (крысы, мыши) на фоне очевидной циркадности поведения (двигательной активности) цикличность в деятельности вегетативных органов не обнаружена.
Очевидно, что цикличность поведения имеет адаптивную природу и задается самыми разными факторами среды от космических тел до колебаний численности кормовых организмов и неизбежной цикличности биохимических и электрических явлений внутри самого организма. Циркадные ритмы обслуживаются разными рецепторными аппаратами животного организма.
Сон
Сон являет собой очень сложное и до конца не изученное физиологическое состояние. Он имеет ряд составляющих — соматические и вегетативные реакции, электрофизиологические явления в мозге, специфические биохимические процессы, которые усложняют понимание механизмов возникновения сонливого состояния, развития глубокого сна и механизмов пробуждения. В любом случае, регулярный сон является абсолютно необходимым для всех млекопитающих. Его депривация на протяжении продолжительного времени приводит к развитию глубоких расстройств функций центральной нервной системы.
Признано, что в процессе сна в структурах нервной системы происходят, с одной стороны, восстановление электрического состояния и ресинтез истраченных химических веществ (энергетический материал, электролитный состав, нейромедиаторы и их предшественники и др.); с другой стороны — катаболизм и удаление продуктов обмена, которые аккумулируются в нервной ткани во время активных действий животного.
Кроме того, складывается впечатление, что во время сна происходит своеобразная «архивация» следов нервного возбуждения. Центральная нервная система как бы замыкается в себе и определенным образом систематизирует, анализирует и архивирует накопленное за день возбуждение. Некоторые структуры головного мозга (кора больших полушарий, лимбическая система) не только отстраняются от восприятия афферентного потока (при глубоком сне животные не реагируют даже на сильные стимулы из внешней среды), но и функционально изолируются от структур, находящихся в их подчинении (продолговатого и спинного мозга).
Инструментальные исследования показали, что в период сна мозг находится в особом активном состоянии. Электрофизиологические исследования свидетельствуют о том, что во сне метаболическая активность переднего мозга заметно возрастает. Причем одновременно с повышением электрической активности и потребления кислорода передним мозгом фиксируют своеобразное разъединение этой части центральной нервной системы с нижележащими структурами.
Торможение нервных процессов имеет место лишь в определенные периоды сна. Оно чередуется с нарастанием возбуждения в коре. Активизация функции головного мозга сопровождается особыми клиническими признаками. В частности, при глубоком сне (человек не реагирует на внешние раздражители и его удается разбудить, только применяя раздражители большой силы) у людей наблюдали быстрые движения глазных яблок и легкие подергивания конечностей.
Похожие явления зафиксировали и у животных. Так, у собаки (Canis lupus familiaris) при глубоком сне на фоне нулевой реактивности нервной системы наблюдали те же движения глазных яблок и подергивания мышц конечностей. Дополнительно у собак фиксировали клонические сокращения поверхностных мышц, подергивания хвоста, ушных раковин и вибрисс. Глубокий сон у собак зачастую сопровождается специфической вокализацией — писклявым тявканьем и оскалом на морде.
Как у человека, так и у животных во время сна отмечали изменения в состоянии вегетативных функций: изменение частоты дыхания и сокращения сердца, повышение температуры тела, изменения артериального давления, у представителей мужского пола — эрекцию полового члена. У человека дополнительно наблюдали эмоциональную активность — появление улыбки или грусти на лице, а также несвязную речь.
Таким образом, на основании полученных при помощи инструментальных методов и клинических наблюдений данных приходится заключить, что при глубоком сне не происходит полного торможения нервной деятельности. Наоборот, отмечается специфическая активизация функций мозга, скелетной мускулатуры и вегетативных органов.
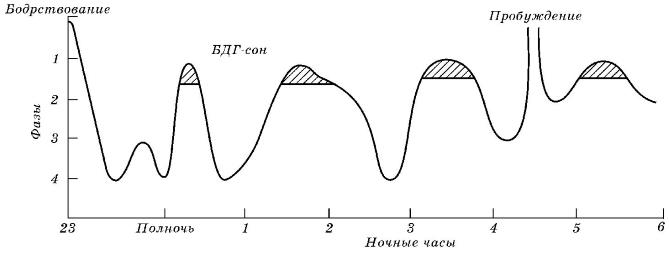
Результаты экспериментов позволяют говорить о сне как о неоднородном физиологическом состоянии животного организма. Та фаза сна, которая сопровождается повышением нервной активности, именуется «парадоксальным» или «быстрым» сном. Ей противопоставлена фаза «медленного» сна. Физиологическое значение парадоксального сна тем больше, чем сложнее поведение и богаче психика животного. У млекопитающих с высокоорганизованной центральной нервной системой парадоксальный сон достигает по продолжительности 1/3 времени сна в целом, у остальных его временная доля в структуре общего сна меньше.
Установлено, что за наступление и регуляцию сна отвечают группы нервных клеток, расположенные в различных мозговых структурах. При усилении возбуждения органов чувств ретикулярная формация повышает возбудимость коры больших полушарий. При этом из лобных зон коры к заднему гипоталамусу устремляется поток, который вызывает торможение центров сна в этой части промежуточного мозга и повышает активность ядер ретикулярной формации среднего мозга. В результате активность коры возрастает еще в большей мере, и животное пробуждается окончательно. Снижение общего афферентного потока, поступающего в таламус и в ретикулярную формацию, снижает восходящее активирующее влияние ретикулярной формации на кору больших полушарий. Параллельно угасает возбудимость лобной коры и ее тормозящее влияние на центры сна заднего гипоталамуса. Эти события сопровождаются развитием дремоты и медленного сна.
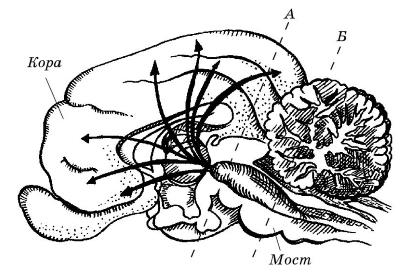
Критические зоны мозга кошки (Felis silvestris catus), регулирующие сон и бодрствование: сечение А (выше среднего мозга) — кошка спит и не в состоянии пробудиться; сечение Б (ниже моста) — кошка постоянно бодрствует и не в состоянии заснуть.
В свою очередь, расторможенные центры сна гипоталамуса за счет своих многочисленных синаптических связей со структурами лимбической системы стимулируют электрическую активность последней. Активизация лимбической системы в отсутствие стимулирующего влияния со стороны ретикулярной формации приводит к тому, что отдельные нейроны коры приводятся в активное состояние. Таким образом развивается парадоксальная фаза сна со специфическими психосоматическими проявлениями и сновидениями. В механизме регуляции сна также участвует мост и продолговатый мозг.
Наличие корково-гипоталамо-ретикулярных взаимодействий делает понятными явления сна у животных с удаленной корой больших полушарий. Новорожденные находятся в состоянии сна большую часть суток по причине неразвитости коры и отсутствия тормозного влияния со стороны лобных зон коры по отношению к центрам сна заднего гипоталамуса. Пробуждение новорожденных связано с тем, что в силу анатомической близости центров голода и жажды в пределах гипоталамуса возбуждение последних иррадиируется на центры сна, т. е. деблокирующее влияние лобных долей коры у новорожденных замещается аналогичным влиянием центров голода и жажды, возбудимость которых очень высока. Поэтому пробуждение новорожденных инициируется исключительно необходимостью удовлетворения их витальных потребностей (пищевых, температурных, болевых). Афферентный поток с хемо-, баро-, термо-, механо — и болевых рецепторов является причиной прерывания постоянного сна новорожденных.
Таким образом, можно заключить, что сон как состояние, при котором происходит временная потеря психических связей животного с внешним миром, протекает на фоне специфической активности головного мозга. Сон развивается в результате возбуждения одних и торможения других мозговых структур. Установлено, что в качестве нейромедиаторов передачи нервного импульса в механизме регуляции сна участвуют такие вещества, как глутатион, серотонин, гистидин. При этом серотонин участвует в формировании медленного сна, а норадреналин является медиатором пробуждения. Также изучена роль мелатонина (продукта превращения серотонина в эпифизе) в чередовании сна и бодрствования.
При монофазном сне 72 % времени приходится на первую половину ночи (по крайней мере, у человека). А весь сон складывается из 4-5 циклов-чередований легкого (медленного) и глубокого (быстрого) сна. При этом продолжительность одного цикла оценивается в среднем в 60-90 минут при соотношении медленный - быстрый сон 5:1. В течение ночи продолжительность отдельных циклов быстрого сна изменяется в сторону увеличения. У животных с полифазным сном и количество циклов, и их продолжительность иные. Причем существенное влияние на структуру сна оказывает этологический фактор. В зависимости от того, какую пищевую нишу занимает животное и консументом какого порядка оно является, количество циклов отдыха может различаться в несколько раз. Так, у кошки (Felis silvestris catus) (типичного хищника) в течение 5-часового периода исследователи насчитывают около 10 циклов, у крысы (Rattus norvegicus) — 22, а у мыши (Mus musculus) еще больше.
У кошки отдых быстро переходит в дремоту и медленный сон. На его фоне в спокойной обстановке развивается парадоксальный сон. В среднем у кошки медленный сон занимает в отдельной фазе сна 20-30 минут, а парадоксальный сон — 3-8 минут. Критерием наступления медленного сна и его отличия от парадоксального служат определенные внешние проявления активности (или ее отсутствия). Во время медленного сна кошка реагирует на сильные и биологически значимые средние по силе звуковые раздражители поворотом ушей в сторону источника звука. В парадоксальном сне кошка выключает центральные механизмы слухового анализатора и не реагирует на звуки даже большой силы (не поворачивает уши в сторону источника звука). У кошки в глубоком сне можно наблюдать быстрые движения глазных яблок, подергивание вибрисс, а также пальцев передних конечностей, ушей и хвоста. Движения глаз кошки цикличны. Они повторяются с частотой 60-70 движений в минуту. Количество движений глаз в одном цикле непостоянно и колеблется от 5 до 50. Очевидно, что парадоксальный (быстрый) сон неоднороден не только по клинической картине, но и по характеру активности нервных процессов, лежащих в его основе.
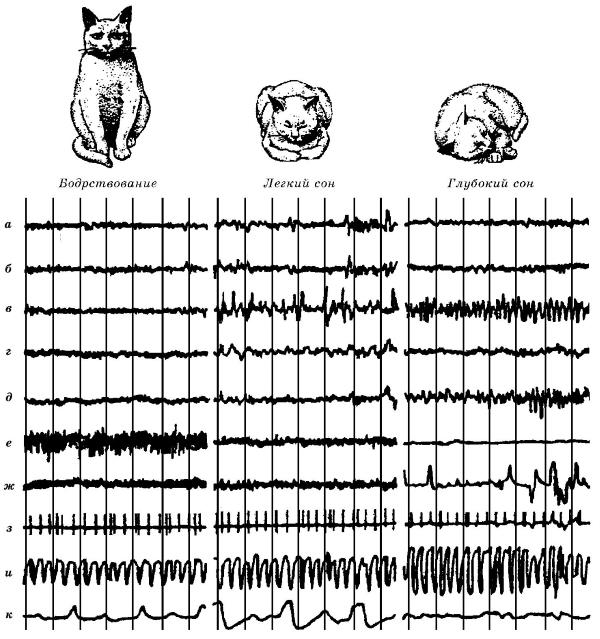
Электрическая активность разных структур кошки (Felis silvestris catus) при бодрствовании, легком и глубоком сне: а — сенсорная кора; б — эктосильвиева кора; в — вентральный гиппокамп; г — ретикулярная формация среднего мозга; д — мост; е — мышцы шеи; ж — движения глаз; з — сердце; и — плетизмограмма; к — пневмограмма.
Чередование периодов циклов сна почти никогда не бывает распределено по суточной шкале равномерно. Так, самцы европейской рыжей полевки (Clethrionomys glareolus), вида с ночной активностью, наиболее подвижными бывают в самом начале ночи и перед рассветом. Животные с дневной активностью отнюдь не спят в течение всего темного времени суток. Например, продолжительность сна у лошади (Equus ferus caballus) составляет всего около 2,5 часов, у косули (Capreolus capreolus) — 1,5-4 часa, у азиатского слона (Elephas maximus) — не более 4-5 часов в сутки. Большинство жвачных (Ruminantia) даже ночью редко спят с закрытыми глазами, по-видимому, их сознание никогда не отключается. Напротив, у оленей (Cervidae) и горных баранов (Ovis) периоды неглубокого сна чередуются с короткими (около 10 минут) моментами настоящей летаргии, когда животные не реагируют ни на какие раздражители, даже на прикосновения. У мелких насекомоядных видов (например, землероек (Soricidae), кротов (Talpidae)), обладающих высоким уровнем обмена веществ и не способных длительно голодать, короткие периоды активности и покоя почти равномерно чередуются в течение всех суток, хотя в принципе землеройки — ночные животные. У кожанов (Eptesicus) на сон отводится примерно 20 часов в сутки.
Хищники в большинстве случаев спят гораздо дольше, чем растительноядные. Причина очевидна: у животных, потребляющих малопитательный растительный корм, на сон попросту не остается времени. Чтобы обеспечить организм необходимым для жизни количеством энергии, они вынуждены подолгу кормиться. Но из этого правила есть исключения. Так, например, коала (Phascolarctos cinereus) спит 18 часов в сутки. Связано это с тем, что поедаемые им листья эвкалиптов трудно перевариваются и содержат много ядовитых веществ, чрезмерное потребление которых может привести к отравлению организма. По этой причине коала вынужден экономить энергию, и тут ему как раз и помогает продолжительный сон, а также малоактивный образ жизни: во время бодрствования он либо занят кормежкой, либо отдыхает. Еще больше спят ленивцы (Folivora). Они также питаются древесной листвой, в которой много волокон и мало питательных веществ.
Однако сон возникает не сразу даже у животных с полифазным характером сна. Возбужденное животное, тем более животное стрессированное, заснуть не сможет (например, кошка, увидев вблизи собаку). Сну предшествует некая подготовка. Животные убеждаются в безопасности окружения, подыскивают для сна подходящее место, принимают видотипичную для сна позу. Мелкие млекопитающие обычно сворачиваются в клубок. Вомбаты (Vombatidae) спят на спине с задранными вверх ногами. Многие копытные благодаря особому строению коленного сустава (своеобразный замок) способны погрузиться в сонное состояние в положении стоя. Так, обычно стоя спит жираф (Giraffa camelopardalis). Он настолько медленно и неуклюже поднимается с земли, что легко может стать в это время добычей хищников. Стоя также дремлют лошади (Equidae), однако глубокий сон у них наступает только в положении лежа на боку. В этой же позе спят и свиньи (Suidae).
Изменение глубины сна и продолжительности циклов быстрого сна у человека в течение ночи.
В жару при наличии большой лужи свиньи могут расположиться на отдых в грязи. Последняя их слабость связана с проблемами терморегуляции. Обладая толстым слой подкожного жира, свиньи чувствуют себя крайне некомфортно при высоких температурах. Грязевые ванны за счет дополнительной теплоотдачи позволяют животным справиться с этой проблемой и поддерживать тепловой баланс во время отдыха. Социальные животные (например, некоторые собаки (Canidae), ластоногие (Pinnipedia) и грызуны) часто спят вповалку, наваливаясь друг на друга. Эта этологическая черта особенно характерна для детенышей-одноплеменников. В больших пометах некоторые особи оказываются с головой придавленными своими братьями и сестрами, однако терпят это неудобство продолжительное время.
Итак, глубокий сон становится возможным только в отсутствие сильных внешних раздражителей при расслабленных скелетных мышцах. Другими словами, главным внешним признаком сна является обособленность животного от внешнего мира, а иногда и оцепенение. У человека на период сна отключается сознание. Многие млекопитающие (вомбаты, медведи, собаки, гориллы и др.) во сне громко храпят.
Дельфины спят наплаву у самой поверхности воды, изредка закрывая глаза на 15-30 секунд. Каждые 30 секунд они делают взмах хвостом, чтобы высунуть из воды верхнюю часть головы и сделать вдох. Во время сна у дельфинов попеременно бодрствует то правая, то левая половина мозга, в то время как другая отдыхает (при этом один глаз животные оставляют открытым, а второй закрывают). Такой «полусон» позволяет дельфинам совершать вдохи и выдохи, которые являются у них произвольными, а также сохранять бдительность на случай появления хищника. Примерно каждые 2 часа половины мозга меняются ролями. В общей сложности сон занимает у дельфинов третью часть суток. Полагают, что в полусне этим водным животным легче контролировать состояние дыхала и узнавать, когда нужно подниматься к поверхности воды за глотком свежего воздуха.

Электроэнцефалограмма левой и правой половины мозга дельфина во время сна.
Исследования, выполненные на человеке и других млекопитающих, свидетельствуют, что в процессе онтогенеза соотношение медленный сон - быстрый сон меняется. Абсолютное время, затрачиваемое на парадоксальный сон, максимально у новорожденных — около 50 % времени всего сна. У недоношенных детей его доля еще больше и составляет 75 %. К концу жизни время, затрачиваемое на парадоксальный сон, сокращается в 4-5 раз и не превышает 10-20 % суммарного времени суточного сна. Имеется в виду, что вместе с характерными изменениями электрической активности мозга с возрастом сокращается и время локомоторных и вегетативных проявлений парадоксального сна.
Интересно и то, что в преклонном возрасте фазовые свойства сна животных и человека закономерно меняются. У человека монофазный сон превращается в полифазный. Старики просыпаются среди ночи по несколько раз, однако днем периодически впадают в сонливое состояние. Общее время сна при этом сокращается. У других млекопитающих по мере старения сокращается количество циклов сна и бодрствования на протяжении суток, и их сон начинает походить на монофазный сон человека. При этом продолжительность одной фазы существенно увеличивается. Скажем, старая собака может проспать, не просыпаясь, всю ночь. Светлое время суток при отсутствии раздражителей большой силы она также может проспать, просыпаясь лишь для отправления своих физиологических нужд (питания, дефекации, мочеиспускания, груминга).
Парадоксальный сон у человека сопровождается сновидениями. С большой степенью уверенности это можно утверждать и о других млекопитающих. Клинические признаки сна со сновидениями у них практически такие же, что и у человека. Однако метод опроса (основное доказательство наличия сновидений) по понятным причинам в случае с животными неприемлем.
Исследования свидетельствуют о том, что обыкновенно людям снятся зрительные образы, их собственные действия и действия других фигурантов, а также события из личного опыта. Реже человеку снятся звуки и запахи. Условия, в которых человек спит, влияют на характер картины сновидений. Так, в жарком помещении может присниться летняя картина. В холодном помещении вероятны сновидения из зимнего опыта человека. Звуковые раздражители влияют на сценарий сна. Не менее сильное влияние на образное содержание снов оказывают и раздражения со стороны внутренних органов спящего (переполнение мочевого пузыря, прямой кишки, физические боли, половое возбуждение, жажда). В этих случаях сны содержат сцены, связанные с удовлетворением возникших физиологических потребностей или снятия дискомфортного состояния.

Лев (Panthera leo) проводит во сне большую часть своей жизни — до 20 часов в сутки. Во-первых, потому, что ему требуется полный покой для переваривания мяса, во-вторых, ему необходимо копить силы для активной охоты и выживания. Потому во время сна лев, как правило, старается полностью расслабиться. Если он ложится спать в спокойном месте, то чаще всего спит на боку или на спине, задрав кверху лапы. Если кто-то может потревожить льва (слон или носорог), то он забирается на дерево и ложится, свесив лапы, полностью расслабляя тело.
Большую ценность для понимания механизма формирования сновидений у животных и человека представляют исследования сна и сновидений у слепых и от рождения слепоглухонемых людей. Оказывается, что у слепых от рождения людей сны лишены зрительных образов. Их сны представляют комбинацию слуховых, обонятельных, вкусовых и тактильных ощущений. Если человек потерял зрение в возрасте старше 6 лет, то он до конца жизни продолжает видеть сновидения со зрительными образами своего детства.
Можно с большой уверенностью утверждать, что основу сновидений у животных составляют зрительные образы с элементами слуховых, обонятельных, вкусовых и тактильных ощущений, которые могут иметь и яркую эмоциональную окраску. Собаки во сне могут яростно лаять или рычать, демонстрируя этим виртуальную злобу или агрессию. Кошки скалятся, рычат, урчат или фыркают на стадии парадоксального сна. Наблюдения за спящими жеребцами-производителями свидетельствуют о том, что они видят эротические сны. Во время глубокого сна на парадоксальной стадии собака иногда принюхивается: учащенно и поверхностно втягивает носом небольшие порции воздуха.
Накопленный фактический материал позволяет утверждать, что сновидения причастны к процессам организации памяти и, следовательно, научения. Эмоциональное переживание событий представляет собой способ сортировки или фильтрации информации, в результате которой эмоционально окрашенные картины остаются в памяти, а наименее значимые — стираются. Следовательно, парадоксальный сон обслуживает механизм запоминания биологически значимой информации. Так или иначе, с точки зрения этологии сновидения участвуют в формировании личного опыта индивидуума.

Поза саванного слона (Loxodonta africana) зависит от фазы сна. Во время медленной фазы слон спит стоя. Во время быстрой фазы он ложится на землю. Перед сном бивни слоны всегда на что-то опирают и в стаде спят с часовыми, меняясь между собой.