в оглавление
назад
Тип неустойчивого движения численности
Многочисленные факты ясно указывают, как особенно чувствительна воспроизводительная система даже к самым слабым изменениям в окружающих условиях. Из этих условий важнейшими являются кормовые. Уже среди зверьков-«эфемеров» имеется целый ряд узкоспециализированных семеноедов, в движении численности которых урожай плодов (и семян) играет роль важнейшего внешнего толчка к массовому размножению. Типичные представители таких грызунов — европейская лесная (Apodemus sylvaticus), желтогорлая (Apodemus flavicollis) и домовая (Mus musculus) мыши, в некоторых лесных районах также рыжая (Myodes glareolus) и красная (Myodes rutilus) полевки. Длительные наблюдения в лесах левобережья Волги у города Казани показали, что наибольшее значение из факторов, определяющих численность рыжих полевок, имеет обеспеченность продуктивными кормами, на втором месте стоят условия зимовки, и, в частности, высота снежного покрова, и на третьем — условия погоды в период размножения. Сходные выводы на 11-летнем материале были получены и при изучении рыжих полевок хвойных лесов северной части Костромской области. Даже среди видов, питающихся грубыми растительными кормами, способных усваивать не только свежую зелень, но и засохшую, выгоревшую траву, сухие побеги и т. п., установлены случаи значительных изменении численности в связи с недостатком или обилием пищи.
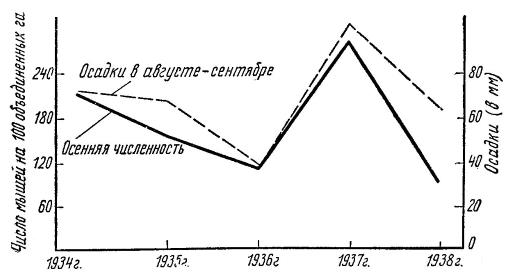
Изменение осенней численности курганчиковых мышей (Mus spicilegus) на юге Украины за 1934-1938 гг. в сопоставлении с величиной осадков за август - сентябрь. Осенние осадки обусловливают увеличение урожая семян сорняков, за счет которых обеспечивается кормом поголовье половозрелых мышей и их успешное размножение.
Большая песчанка (Rhombomys opimus) — грызун, свойственный полупустыням и пустыням Азии, отличается крайней неприхотливостью в отношении выбора кормов. Она поедает большинство видов растений, имеющихся по соседству с ее норой, в том числе формы, совершенно непривлекательные для других фитофагов — жгучие, эфироносные, колючие, ядовитые. Так, например, она без видимого вреда для себя питается гармалой — ядовитым для скота растением, содержащим несколько алкалоидов. Гармала часто и в большом количестве селится на выбитых скотом пастбищах, так как домашние животные в период вегетации ее не трогают, а конкурентов поедают и угнетают. Казалось бы, что столь нетребовательный грызун, как большая песчанка, всегда может найти достаточное количество корма. Выяснилось, однако, что в условиях засушливого климата полупустынь и пустынь нередки годы, крайне бедные осадками, когда растительность сильно угнетена и дает ничтожно малую продукцию. Большая песчанка — типичный норник, она кормится не далее 40-60 м от норы, куда готова скрыться при первом признаке опасности. В засушливые годы участок диаметром 100-120 м не может полностью удовлетворить потребности семьи песчанок в корме. При более далеких рейсах зверьки чаще становятся добычей хищников. Состояние растительности в пустынях, особенно наиболее ценных в кормовом отношении эфемеров, эфемероидов, и части кустарничковых многолетников (полыней, солянок) зависит от запасов почвенной влаги, непосредственно обусловливаемых величиной осадков, выпадающих в холодную половину года (летом осадков в части пустынь или совсем не бывает, или, там, где выпадают, они не успевают впитываться в почву и полностью испаряются). Длительные наблюдения зоологов противочумных станций показали, что численность больших песчанок зависит в основном от состояния растительности в период их размножения и, в конечном счете, от количества осадков за предшествующую зиму.
Установлено также, что сходное движение численности достаточно постоянно повторяется у популяций больших песчанок, занимающих целые природно-географические области. Это легко понять, зная, что условия погоды обычно бывают одинаковыми на больших пространствах равнин Южного Казахстана к Средней Азии. Рисунок ниже показывает, как следом за сезонами с хорошим зимне-весенним увлажнением поднималась численность больших песчанок в Панфиловском районе Казахстана (идеального совпадения кривых нет, так как на амплитуде изменений численности этих грызунов сказывается и величина поголовья производителей, сохранившихся до весны — момента начала размножения). Хорошая осенняя вегетация солянок при теплой погоде обусловливает появление новой, третьей волны размножения и смещение пика численности на позднюю осень. Неблагоприятный ход весенних температур, а именно дефицит тепла, поздние заморозки могут задержать размножение и оказать отрицательное воздействие и на второй генеративный цикл.
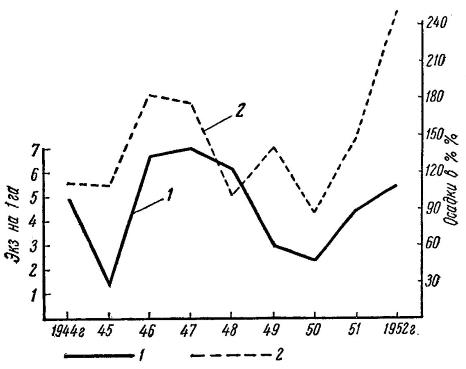
Изменение численности больших песчанок (Rhombomys opimus) в зависимости от количества осадков: 1 — количество осадков за октябрь - май 1944-1952 гг. в Панфиловском районе Южного Казахстана; 2 — среднее число песчанок на гектар.
В целом наиболее важны обеспеченность кормом, с которой связан и ход смертности производителей, и интенсивность их размножения. Поэтому при прогнозах численности больших песчанок, занимающих отдельные природно-географические области, основное внимание уделяют характеристике погоды холодного периода и состояния растительного покрова в весенние месяцы и осенью. Большая песчанка играет первостепенную роль в эпизоотии чумы в пустынях Средней Азии и Казахстана. Прогнозирование движения численности этого грызуна облегчает организацию борьбы с эпизоотиями чумы и помогает предупреждать возможность заболевания людей.
У белки (Sciurus vulgaris), находящейся в такой же зависимости от урожаев семян лесных пород, как, например, желтогорлая мышь (Apodemus flavicollis), все же вследствие ее меньшей плодовитости не бывает столь быстрых и резких подъемов и падений численности. Малая плодовитость (по сравнению с мышевидными грызунами) обусловлена ее большей стойкостью и относительно высокой продолжительностью жизни: в природных условиях небольшая часть белок доживает даже до 6-7 лет. Для мелких мышей в урожайный на семена год характерно длительное осенне-зимнее размножение, начинающееся обычно после опадения на землю желудей дуба, орешков бука, липы и др. и продолжающееся до начала лета следующего года включительно. Белки же при высоком урожае, например семян ели, успевают за счет высокоценного свежего корма принести лишь один выводок в конце лета того же года — конце августа - сентябре (они начинают использовать семена нового урожая при их молочной спелости, в лесах северной половины России — с конца июня или начала июля). В основном же используют его в теплую половину следующего года, поэтому и подъемы их численности всегда отстают на один год по сравнению с урожайными годами лесных пород.
Спектр кормов, потребляемых белкой, достаточно широк: в него входят семена и орехи лесных пород, ягоды, почки ели, березы и других деревьев, грибы шляпочные и почвенные (трюфели), нежная кора и сок побегов сосны, ели, пихты, березы, лишайники, трава и листья, личинки мух, куколки и взрослые муравьи, изредка яйца и птенцы. Несмотря на такое разнообразие кормов, давно установлено, что белка и по морфологическим (строение зубов, желудка, кишечника), и по биологическим особенностям — узкоспециализированный потребитель концентрированных кормов, а именно семян лесных пород.
На преобладающей части ареала важнейшую роль в питании белки играют семена ели, лиственницы и кедров (сибирского, корейского, кедрового стланика), в меньшей степени — сосны, дубов и лещин. Семена хвойных пород бывают в изобилии не всегда, а только в годы высоких урожаев, повторяющихся раз в несколько лет. Перерывы между урожаями зависят от биологии данной древесной породы, а также от широты местности и степени континентальности климата в разных частях ее ареала. В условиях равнинной тайги нередко пространства в сотни тысяч квадратных километров еловых или кедровых насаждений одновременно приносят высокий урожай или поражаются полным неурожаем.
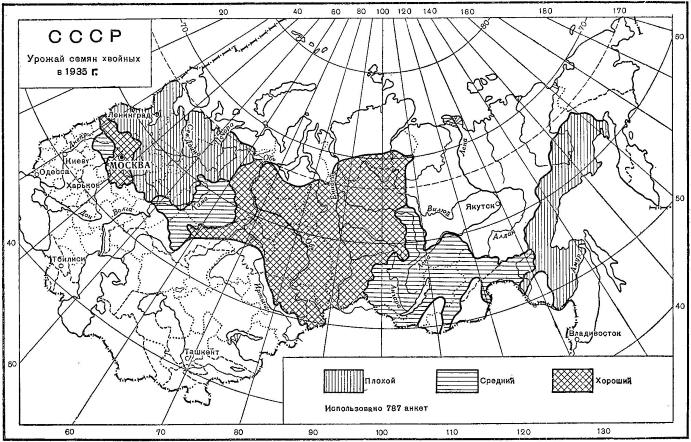
Урожай семян хвойных пород осенью 1935 г. по глазомерной оценке в баллах.
Условия добывания корма белкой зависят не только от величины урожая, но отчасти и от деятельности многочисленных конкурентов: насекомых, повреждающих шишки, клестов, дятлов, кедровок, в массе уничтожающих семена. Если корм с осени осыпается на землю (шишки кедра, желуди дуба), большую роль играет также структура снежного покрова в данную зиму: появление наста, корок гололеда очень затрудняет отыскивание и выкапывание белкой пищи из-под снега. Вызываемое недостатком семян изменение диеты белки (использование вынужденных и балластных кормов) обусловливает истощение организма животного, заболевания, иногда, кроме того, откочевки и в конечном итоге — массовую гибель. Особенно много истощенных белок погибает в морозные месяцы зимы и при позднеосенних миграциях, что сильно сокращает количество производителей перед началом гона. Длительное использование биологически неполноценного корма сильно сказывается на половой функции белки, вызывая резкое снижение плодовитости и даже полное прекращение размножения.
В нормальных условиях большинство самок приносит в течение года два помета (первый весной, второй — летом), производя в среднем 9 бельчат. Осенью молодые белки составляют в популяции около 82 % (большинство из весеннего помета). Соотношение полов в популяции 1: 1. В годы, несколько худшие по кормовым условиям, число пометов остается обычным, но среднее число бельчат на самку к концу сезона размножения не превышает 4,9-5,8 (большинство из летнего помета), а молодые составляют в популяции около 70 %. После бескормной зимы размножения совсем не бывает, реже часть самок приносит один весенний или один летний помет (процент яловости очень велик). Количество молодых к осени составляет значительно меньше 50 %, число самок становится ниже нормы. В итоге после неурожайного года, а особенно после двух последовательных неурожайных лет численность белки резко сокращается; из обычного массового грызуна она временно превращается в редкое и малозаметное животное леса. Условия погоды непосредственно в гораздо меньшей степени влияют на движение численности этого грызуна, чем на нестойких гетеротермных полевок. В зависимости от чередования семенных и неурожайных лет основной кормовой породы «урожаи» белки повторяются с промежутками в 3, 4, 5, 6 лет. Даже в пределах одной области или республики (например, Якутской) разные ее районы могут существенно отличаться в частоте «урожаев» белки, в их величине и т. д. Это обусловливается особенностями лесных площадей, средними размерами урожая семян, продолжительностью периода размножения белки и т. п.
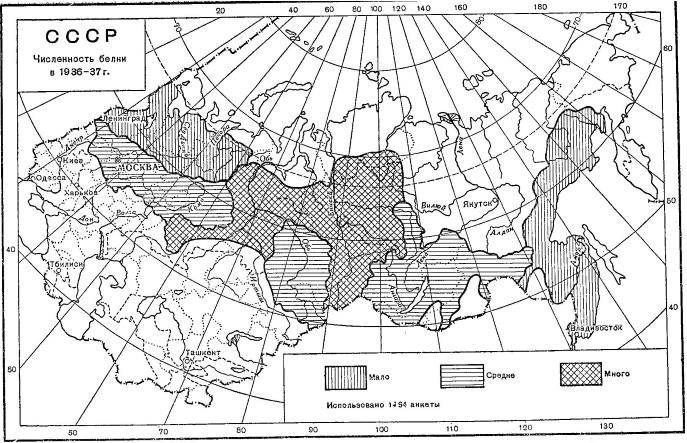
Количество обыкновенной белки (Sciurus vulgaris) в промысловый сезон 1936/37 гг. При сопоставлении с картограммой урожая семян хвойных 1935 г. видно, что в обширных районах, не имевших кормов, белки было мало. Наоборот, после хорошего урожая семян в Западной Сибири почти со всей этой территории поступили сообщения о хорошем «урожае» белки.
Подобного рода зависимость численности от изменчивости обилия кормов установлена для песца (Vulpes lagopus), лисицы (Vulpes vulpes), горностая (Mustela erminea) и ряда других зверей. Еще 200 лет тому назад со слов якутских промышленников было записано, что «урожаи песцов» на северо-востоке Сибири повторяются с известной правильностью через каждые 3-4 года. Специальные исследования, проведенные в той же Якутии, показали, что этот 3-4-летний ритм колебаний численности остался неизменным. Для севера Канады имеются цифры за большой ряд лет, показывающие, как такие же изменения количества песцов отражаются на заготовке его шкурок. Так, например, в одном из участков северной Канады численность песца была наибольшей в годы: 1856, 1861, 1864, 1869, 1873, 1878, 1884, 1888, 1892, 1896, 1899, 1903, 1907 и 1911. Следовательно, «урожайные» годы повторялись здесь через промежутки в 4, 3, 3, 5, 3, 3, 4, 4, 4, 4, 4, 4, 4, 5, 4 лет, в среднем — 3,9 года. Сходна цикличность подъемов и падений численности песцов установлена наблюдениями зоологов и промышленников на Европейском Севере России, включая Новую Землю, и в тундрах Сибири, причем годы урожая песца в Якутии не совпадают с таковыми на Ямале и т. п.
Следовательно, одновременным урожаем или неурожаем охватывается только треть или четвертая часть огромного протяжения российских тундр. Повсеместно высокая численность песцов обусловливается массовым размножением леммингов — на западе — норвежского лемминга (Lemmus lemmus), а восточнее Белого моря — главным образом обского (Lemmus sibiricus) и отчасти копытного (Dicrostonyx torquatus). Наблюдениями на Ямале установлено, что в зимы, когда идет подснежное размножение леммингов, в желудке песцов часто находят целые выводки, выкопанные из гнезд. После такой зимы обычно и летняя численность леммингов оказывается высокой, что обеспечивает выкармливание больших пометов песца. Был предложен остроумный способ прогноза численности песцов на следующий промысловый сезон, основанный на учете встреч молодых леммингов в желудках песцов, добытых в течение предшествующего сезона.
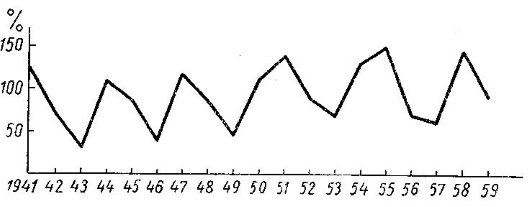
Колебания численности песца (Vulpes lagopus) в Восточной Сибири.
Плодовитость песца в годы массового размножения служащих им пищей грызунов высока: в выводке нередко бывает 8-12 подросших молодых. Наибольшее известное число эмбрионов — 16, а количество послеродовых пигментных пятен в матке до 26. Хорошо обеспеченные продуктивным кормом осенью и в течение всей зимы песцы не рассеиваются по тундре, лесотундре и льдам моря, а держатся близ нор и рано приступают к гону; овуляция и беременность самок протекает нормально, они легко вынашивают и выкармливают многочисленных щенков. В такие годы заселенность нор бывает высокой. Так, например, в Карском и Пайхойском очагах размножения песца в 1955 г. было заселено 100 % осмотренных нор, но в 1956 г. только 20 %, в 1957 г. — 20 % и 10 % (в Пайхойском), а в 1958 г. снова 80 % и 100 %. Среднее число выкормленных щенков в выводке в августе было 8 (1955 год, обилие леммингов), 4 (1953 и 1956 гг., леммингов крайне мало или нет) и даже 3 (1957 г., лемминги отсутствовали).
В годы низкой численности леммингов и узкочерепных полевок (Microtus gregalis), которые тоже служат ценным кормом, но играют в жизни песца второстепенную роль, многие самки этого хищника остаются бесплодными или приносят слабых щенят, погибающих уже осенью. При остром недостатке пищи отдельные пары бросают свои выводки. В течение одного-двух бескормных лет население песцов резко уменьшается, так как при предельно сократившейся плодовитости одновременно идет интенсивное вымирание производителей.
Осенью в год высокой численности песцов, часто совпадающий с вымиранием леммингов, начинаются массовые перекочевки хищников, среди них распространяются широко разлитые эпизоотии, наносящие огромный урон поголовью. Преобладают случаи поражения нейротропным вирусом — болезнь, известная на севере как «дикованье». «Дикующий» песец крайне возбужден, беспокоен, подвижен, утрачивает всякую осторожность — смело бросается на упряжки ездовых собак, нередко заскакивает в освещенные окна промысловых избушек и т. д. Ездовые собаки, лисицы и волки тоже вовлекаются в эти эпизоотии. Среди охотников существует мнение, что падеж ездовых собак и песцов вызывается бешенством. «Дикованье», действительно, напоминает его некоторыми симптомами.
В тундре большинство песцов умирает в первую же зиму после начала эпизоотии. Количество трупов, вытаивающих весной с сохранившимся, хорошим мехом, нередко бывает огромным. Практикуется даже специальный промысел и заготовка шкурок павших песцов. На оленьих упряжках промышленники обследуют проталины тундры и отыскивают белых, издалека заметных зверьков. Так, например, в районе одной только речки западносибирской тундры однажды нашли 130 павших песцов, большинство которых было в зимнем меху и отличались высокой упитанностью. В другой раз на протяжении примерно 20 км пути были насчитаны остатки 16 трупов песцов. В Большеземельской и Малоземельской тундрах после сильных вспышек заболевания весной находили иногда до двух трупов на 1 кв. км.
Годы появления большого числа леммингов в Большеземельской и Малоземельской тундрах были зафиксированы, в частности, в следующие годы: 1926, 1929, 1932, 1935, 1940, 1943, 1946, 1949, 1952, 1958. Таким образом, урожай леммингов в российских тундрах повторяется через два года на третий. Это дает возможность составлять многолетние прогнозы урожаев леммингов и песцов, основываясь на длительном изучении экологии песца северо-востока Европейской части России. Таким образом, движение населения песца с его очень лабильной, резко колеблющейся по годам плодовитостью, в благоприятные годы почти в два раза превышающей плодовитость лисицы, отличается частыми подъемами численности и быстрыми ее сокращениями. Пик численности длится всего один сезон; цикличность изменений копирует свойственную массовым грызунам тундры — леммингам — важнейшему продуктивному корму песцов.
Движение численности лисицы (Vulpes vulpes) в основных чертах сходно с описанным для песца, но имеет и существенные отличия. Лисица распространена необычайно широко и населяет все природно-географические зоны России и ряда прилежащих стран, отличающиеся и по климатическим условиям и по характеру фауны наземных животных, многие из которых служат кормом этого хищника. Популяции лисиц, занимающие разные природно-географические зоны, характеризуются каждая своими особыми вариантами движения численности, черты которых обусловлены как естественными биотическими и абиотическими факторами, так и влиянием хозяйственной деятельности человека.
В средней и степной полосе России, где в населении мелких грызунов господствуют обыкновенные (Microtus arvalis) и общественные (Microtus socialis) полевки и степные пеструшки (Lagurus lagurus) подъемы и падения их численности, повторяющиеся обычно с промежутками в 2-3 года, вызывают соответствующие, но более сглаженные, изменения количества лисиц. Картина осложняется проявлением влияния «больших волн» массового размножения грызунов, повторяющихся с интервалами в 9-10 лет. Исключительное, но кратковременное обилие мелких зверьков, на огромных пространствах степной, лесостепной и отчасти лесной полос, не может не вызывать соответствующих сдвигов в плодовитости и смертности лисиц. Наличие обусловленных этими «большими волнами» и почти синхронных с ними размножений лисиц служит убедительным доказательством реальности существования вспышек размножения грызунов с 10-летней цикличностью.
При всем разнообразии используемых кормов полевки и пеструшки играют для лисицы роль самого важного продуктивного корма, особенно осенью и зимой. В годы обилия полевок хищники хорошо упитаны всю зиму и весной успешно размножаются. В 1921-24 гг. и через 10 лет — в 1932-34 гг. массовым размножением грызунов были охвачены средняя и южная части Европейской территории России, Белоруссия и Украина. На один-два года позднее (время, необходимое для использования хищниками благоприятной для размножения ситуации) следовали большие подъемы численности лисиц, хорошо отраженные в изменениях заготовки шкурок этого вида на Кавказе, Украине, в Нижнем и Среднем Поволжье, черноземной полосе и областях Промышленного Центра за 1924-29 и 1933-35 гг. (следующая грандиозная вспышка размножения пришлась на 1943-45 гг. и была далеко не полностью использована охотниками).
Судя по количеству добываемых лисиц, численность их в годы, благоприятные для размножения, увеличивается не менее чем в 5-7 раз. Это подтверждается и непосредственными наблюдениями. В период предзимья, в год большого «урожая» лисиц с одного места можно одновременно увидеть в Ставрополье и Нижнем Поволжье до 10-12 мышкующих на полях лисиц, а в Подмосковье — до 4-5, тогда как в неурожайные — не более 1-2.
Ухудшение условий добывания корма в осенне-зимний сезон, предшествующий периоду гона, вызывает сильное снижение плодовитости лисиц. Прежде всего, возникает несоответствие в сроках готовности к спариванию у самцов и самок. Например, в малокормном 1939 г. в Ставрополье максимум готовых к гону самцов приходился на январь, большинство же самок было в состоянии течки во второй половине февраля, а у некоторых она наступила только в марте. В этих условиях самцы, рано заканчивающие гон, не могут покрыть всех самок, и процент прохолоставших лисиц резко возрастает. Кроме того, в связи с длительным полуголодным существованием истощенный организм уже оплодотворенной самки во многих случаях оказывается не способным обеспечить нормальное развитие эмбрионов. В упомянутом 1939 г. из шести добытых в Ставрополье самок у пяти резорбировались полностью все зародыши и только у одной была частичная резорбция эмбрионов (из 8 погибли 2).
Все это приводит к тому, что после голодной зимы количество пустующих выводковых лисьих нор увеличивается соответственно большому проценту прохолоставших или не ощенившихся лисиц. В 1939 г. в Ставрополье к концу весны таких самок было 67 %. Но среднее число лисят в выводках и в этих условиях оставалось близким к обычному. В 1937 г., благополучном по обеспеченности кормом, средняя величина выводка была 6, в 1938 г. — 5,4. Таким образом, уменьшение плодовитости лисиц в неблагоприятные годы идет, видимо, прежде всего за счет сокращения числа выводков, увеличения процента холостых особей, а не за счет уменьшения числа щенков в помете (однако по некоторым наблюдениям в Тульских засеках в 1938 г. при хороших кормовых условиях среднее число лисят в выводке было 5,5, а в следующем малокормном 1939 г. — всего 2 щенка).
В Крапивенском районе Тульских засек на учетной площади леса в 4465 га, пересеченной множеством оврагов и имевшей много нор, количество выводков лисиц за десятилетие (1936-45 гг.) изменялось следующим образом:

Поскольку одновременное снижение численности полевок нередко охватывает обширные территории (например, все степные и лесостепные области юга бывшего СССР от Украины до Западного Казахстана и от Предкавказья до Нижнего Поволжья), количество лисиц сокращается на таком же пространстве. Помимо снижения плодовитости длительный недостаток продуктивного корма вызывает увеличение смертности взрослых лисиц: часть их погибает от голода, часть от болезней, усиливающихся в связи с истощенностью животных, их большей подвижностью и т. д. Количество лисиц становится особенно большим вскоре после массового размножения грызунов, которое, как уже отмечено выше, обычно заканчивается быстрым вымиранием. При этом сильно размножившиеся лисицы в течение двух-трех лет оказываются в крайне неблагоприятных кормовых условиях. Такое положение возникло в средних областях Европейской части СССР в 1945-46 гг. По причине полного прекращения охоты в годы Великой Отечественной войны маточное поголовье лисиц было особенно большим, а после массового появления полевок в 1945 г. и успешного размножения количество хищников необычайно увеличилось. Начиная с осени 1945 г. и весь 1946 г. поступали донесения от охотников и лесников о наличии среди лисиц какой-то эпизоотии в популяции Крапивенского района Тульской области. Лисицы не уходили от гончих собак, иногда падали мертвыми во время гона; некоторые лежали на земле, не имея сил подняться при приближении людей. Часто попадались лисьи трупы. В эти годы в Калужской, Московской и других областях среди лисиц были отмечены отдельные случаи бешенства и более многочисленные — другой болезни, видимо, энцефалита. В западных областях резко увеличилось количество лисиц, пораженных чесоткой.
В зоне тайги и лесотундры лисица зимой испытывает сильные затруднения в добывании мышевидных грызунов из-за большой мощности снежного покрова. Как показали наблюдения, проведенные в тайге верхнего течения реки Печоры, основу зимнего питания местной лисицы составляют заяц-беляк (Lepus timidus) и тетеревиные птицы; по частоте встреч — 59 % — беляк стоит на первом месте среди других кормов. Лисица систематически пользуется тропами беляка при передвижении в многоснежный период и на них же подкарауливает и скрадывает зайцев. Учеты лисиц и данные заготовок показали наличие значительных колебаний их численности в Печорской тайге: минимальные и максимальные показатели разнятся в 9-12 раз. После зим, обильных кормом (беляком или тетеревиными), количество лисиц в районе остается стабильным, а в течение наиболее голодных зим их погибает около 75 %. Судя по имеющимся данным, основная масса лисиц умирает от крайнего истощения и небольшую часть ослабевших ловят росомахи (Gulo gulo). Зимой 1940/41 гг. у погибших от голода лисиц отношение веса тушки (в граммах) к длине (в сантиметрах) равнялось 30, тогда как этот показатель упитанности у особей, добытых в обильную кормами зиму 1937/38 гг., равнялся 54.
Глубокий и рыхлый снежный покров тайги крайне ограничивает набор кормов, доступных лисице, и ставит хищника в тесную зависимость от зайца-беляка, подверженного очень резким колебаниям численности. Это в основном и обусловливает движение населения таежных лисиц по годам, что давно подмечено при сопоставлении кривых заготовок лисьих и заячьих шкурок в тайге Сибири и Северной Америки. Здесь высокая численность лисиц отмечается с промежутками в 8-10 лет. Очень отчетливо цикличность колебаний количества лисиц выражена в северной Канаде. В песчаных пустынях Средней Азии мелкие лисицы местной географической расы (Vulpes vulpes flavescens) связаны по питанию более всего с песчанками (Gerbillinae), отчасти с сусликами (Spermophilus), тушканчиками (Dipodidae). Зимние условия кормодобывания здесь мало отличаются от летних, так как снежного покрова почти не бывает. Несомненно, что детали динамики численности лисиц здесь иные, чем на севере.
Изучение в течение пяти смежных лет кормового режима и плодовитости соболя (Martes zibellina) Зауралья показало, что хорошее размножение этого хищника обусловливается обилием лесных полевок (Myodes), серых полевок (Microtus) и лесных леммингов (Myopus schisticolor). Прочие корма (птицы, белки (Sciurus vulgaris), беляки (Lepus timidus), землеройки (Soricidae), насекомые, орехи, ягоды и др.), каждый в отдельности, имеют для соболей второстепенное значение, но в совокупности играют роль, в количественном отношении близкую к таковой мышевидных грызунов. В годы обилия рыжих, серых полевок и лесных леммингов — продуктивного корма соболей, 90-100 % соболюшек участвуют в гоне и оказываются оплодотворенными. Число эмбрионов, судя по подсчету желтых тел беременности, колеблется от 2 до 6, а в среднем равняется 4-4,5. Число соболей-сеголетков в пробе из популяции составляет около 50 %, соотношение полов равномерное. В год с ухудшением запасов корма (уменьшение количества полевок, неурожай семян кедра) число оплодотворенных самок равнялось 70 % от общего числа исследованных, среднее количество эмбрионов было 3,2 (при колебаниях 2-4). Число сеголетков в популяции составляло 32 %. Среди добытых зверьков резко повысилось (до 48 % от общего числа) количество взрослых самцов. Для близкого вида — лесной куницы наличие 50 % прибылых животных в добыче охотников считается показателем хорошего состояния популяции, от 50 до 30 % — удовлетворительного и ниже 30 % — плохого, т. е. отличавшегося низкой плодовитостью в предшествующий сезон размножения. Колебания плодовитости несомненно оказывают влияние на изменение численности соболей, но, при свойственной виду значительной стойкости, падения и подъемы кривой, показывающие движение их населения, гораздо более плавны, и не столь резки, как у грызунов или узкоспециализированных хищников мышеедов, отличающихся относительно высокой плодовитостью. Урожайные и неурожайные на соболя годы повторяются с интервалами в 11 лет. Подъемы численности близкой к соболю американской куницы (Martes americana) повторяются в среднем через 9,5-10 лет.
К группе видов с неустойчивой численностью населения принадлежат и зайцы, в частности русак (Lepus europaeus) и беляк (Lepus timidus). Беляк — вид с огромным ареалом, в разных частях которого движение населения имеет некоторые местные отличия. Он приспособлен к жизни в тундре, лесотундре, тайге, смешанных лесах, лесостепи и даже степи, если находит в ней осиновые колки и участки, поросшие ивами, таволгой и т. п. Как уже указывалось, обычно он не устраивает постоянных нор и даже детенышей приносит в простых, почти ничем не укрытых логовах. Несмотря на это зайчата, быстро приобретающие самостоятельность, относительно мало страдают от нападений хищников и непогоды (целиком погибают только ранние выводки в случае возврата холодов). По своей неприхотливости в отношении питания беляк напоминает лесных оленей, питающихся большую часть года веточным кормом, и некоторых серых полевок. Как и для этих животных, состояние и доступность корма в движении численности популяции беляка играют второстепенную роль. Беляки почти оседлы, массовые перекочевки бывают у них исключительно редко. В годы высокой численности за зимний период питания веточным кормом и корой они наносят очень большие и хорошо заметные повреждения кустарникам и молодым деревцам — ивам, осине, рябине, можжевельнику, дубу, лещине и многим другим. Особенно серьезно страдают от них лесные питомники и молодые плодовые сады. Однако год высокой численности вскоре сменяется периодом депрессии, и беляк становится иногда настолько редким зверьком, что за целое лето работы в лесу его совсем не удается встретить. В течение нескольких лет лесная растительность полностью оправляется от повреждений, а излюбленные биотопы беляка очищаются от рассеянных зайцами бесчисленных яичек гельминтов. Сильные «заячьи моры» известны издавна в Евразии и на материке Северной Америки. В северной полосе России весной после мора в мелколесьях и на зарастающих вырубках иногда почти под каждой кучей хвороста находят трупы зайцев в зимнем меху. В литературе есть много описаний опустошительных эпизоотии, вызывавших резкое падение численности американского беляка (Lepus americanus) на пространстве от северной Аляски до южных штатов. Так, в 1887 г. из окна вагона Канадско-Тихоокеанской железной дороги на протяжении 50 миль можно было видеть трупы зайцев, которые лежали сотнями, если не тысячами.

Связь колебаний численности хищников (рысь (Lynx lynx), волк (Canis lupus), лисица (Vulpes vulpes)) и жертвы (заяц-беляк (Lepus timidus)) в центральных районах Европейской части России в 1932-1954 гг.
В России в таежной полосе численность беляка колеблется в 4-6-кратных пределах, а в лесостепной — в 3-4-кратных. У северо-восточной границы ареала, в отдельных районах Якутии по недавним исследованиям количество беляков всего за 5-10 лет изменяется в сотни и даже в 2500 раз. Якутия давно известна крайней неустойчивостью популяций зайца-беляка; здесь продолжительность циклов (от одного пика до другого) составляет около 12 лет. В Западной Сибири она равна 8-10 годам, на Европейском Севере России 9-11, в Ленинградской области — 5-7, в Московской — 4-5 и т. д. Следовательно, в областях с более суровыми природными условиями нужно большее число лет, чтобы сохранившиеся производители могли обеспечить достаточный рост популяции.
Продолжительность жизни зайца равна 6-7 годам, плодовитость его довольна велика. В средней полосе беляк чаще приносит в год только 2 помета; в первом, ранневесением гоне принимает участие до 83-100 % самок, во втором, весеннем — практически все половозрелые самки. В третьем, летнем гоне в средних и южных областях России и прилежащих странах участвуют не более 30-40 % самок беляка, а в северных — 10-15 %. В полосе тундр больше двух выводков вообще не бывает по причине малой продолжительности теплого периода года (уже в 9-10-дневном возрасте зайчата начинают есть траву; таким образом, их благополучие зависит от наличия свежей зелени, а длительность всего периода размножения беляка — от продолжительности вегетационного периода). Числа молодых в помете колеблется от 1 до 9, в среднем 3,5-5,2 зайчат. Наибольший приплод дает второй — летний помет.
Выяснено, что плодовитость беляков меняется по годам, причем это обусловливается изменением величины выводка, тогда как процент самок, участвующих в размножении, остается постоянным (несомненное влияние относительно равномерной обеспеченности кормом в разные годы). Основная причина колебаний плодовитости — интоксикация организма самок, возникающая на почве глистных и протозойных инвазий. Она уменьшает вероятность имплантации оплодотворенных яиц, вызывает резорбцию эмбрионов. Но изменение плодовитости по годам — не единственная причина колебаний численности беляка, оно только усиливает несоответствие между рождаемостью и смертностью. Последняя в фазу падения численности особенно велика и в основном обусловлена гибелью зайцев от болезней и деятельностью хищников, уничтожающих много ослабевших зайцев.
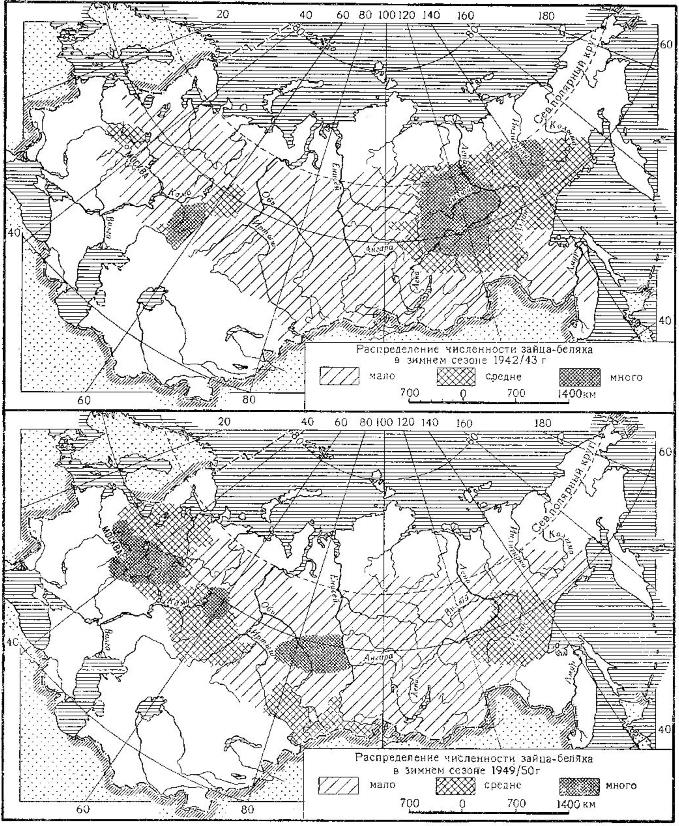
Распределение численности зайца-беляка (Lepus timidus) в 1942/43 и 1949/50 гг.
Беляки бывают сильно инвазированы нематодами Protostrongylus (до 100 % зараженных при среднем количестве от 25 до 180 экземпляров глистов на одного зайца) и Trichostrongylus (до 100 % зараженности при среднем числе 4-570 экземпляров на одного зайца), а также заражены кокцидиозом. Токсичность легочных нематод выше, чем кишечных. Наиболее широкое распространение болезни получают в холодные дождливые годы при достаточно высокой численности беляков. Своеобразная особенность пищеварения зайцев заключается в том, что дефекация у них происходит не на местах отдыха, а в основном во время кормежки; поэтому участки с зарослями привлекательных кормовых растений всегда особенно густо усеяны их «орешками». Это приводит к сильной зараженности таких участков яичками гельминтов, что естественно облегчает обмен эндопаразитами между зайцами и усиливает интенсивность инвазий. На постоянно влажной почве и лесной подстилке дольше сохраняют жизнеспособность яйца и личинки гельминтов, ооцисты кокцидий, и, следовательно, легче происходит заражение. Засушливые годы задерживают распространение инвазий даже при большой численности беляков. С этими особенностями этиологии гельминтозов и кокцидиоза связан и тот факт, что сильные заячьи моры более обычны в равнинных заболоченных местностях с подзолистыми почвами. В щебнистых и песчаных районах с хорошим дренажем эпизоотии случаются реже и уносят меньше жертв.
Таким образом, на движение численности беляка условия погоды воздействуют не прямо, а через посредство паразитических организмов. Они вызывают у беляков снижение плодовитости и тяжелые заболевания, нередко кончающиеся смертью наиболее инвазированных особей. В то время как для мелких грызунов большое значение имеет погода осенью, зимой и весной, в жизни беляка эту роль играют метеорологические условия лета, когда происходит развитие во внешней среде его паразитов или их промежуточных хозяев.
вперед
в оглавление