Мускулатура

Поверхностная мускулатура передней части тела австралийской ехидны (Tachyglossus aculeatus).
Все туловище словно мешок одевает огромная подкожная мышца (panniculus carnosus). Она толстая на спине и боках, но тонкая в центре брюшной поверхности, где у беременной самки формируется сумка. Сокращая различные части подкожной мышцы, ехидна меняет форму своего тела. Она может свернуться в почти полный шар, спрятав уязвимое брюхо и выставив наружу выпрямленные иглы.
Мышцы конечностей, их поясов и туловища тоже развиты очень хорошо. Так, мощная двуглавая мышца начинается на метакоракоиде и прокоракоиде, проходит через верхнюю головку плеча и заканчивается на теле лучевой кости гораздо ниже, чем у человека, образуя рычаг третьего порядка. В сочетании с короткой и массивной плечевой костью это позволяет передней конечности совершать сильные движения.
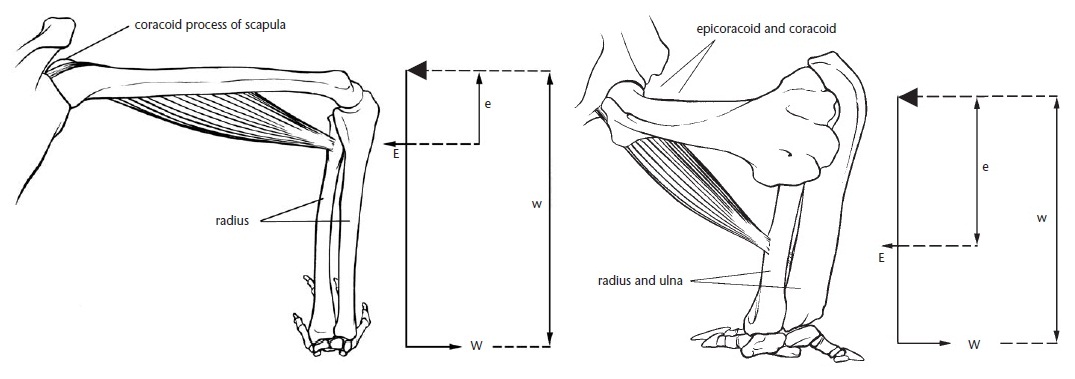
Сравнение механического преимущества в сгибании локтя с помощью бицепса у человека (Homo sapiens) (слева) и австралийской ехидны (Tachyglossus aculeatus) (справа). Эта конструкция представляет собой рычаг третьего порядка. Точкой опоры выступает локоть, рычагом — объединенные лучевая и локтевая кости. Механическое преимущество рычага (W/E) равно длине плеча силы (e, расстояние от локтя до места прикрепления бицепса), разделенному на длину плеча нагрузки (w, расстояние от локтя до передней лапы). Чем длиннее плечо силы по отношению к плечу нагрузки, тем больше механическое преимущество. У человека бицепс прикрепляется к лучевой кости в ее верхней 1/4, поэтому плечо силы довольно короткое. У ехидны место прикрепления бицепса находится примерно посредине лучевой кости, что обеспечивает ей большее механическое преимущество по сравнению с человеком. Большая стрелка указывает точку опоры — локоть.
Мышцы морды, нижней челюсти и языка сильно специализированы для питания муравьями и термитами. Вытянутое клинообразное рыльце дает значительное механическое преимущество для создания плеча рычага при проникновении в труднодоступные места. Червеобразный язык может высовываться из ротовой щели на 15-18 см, а его полная длина достигает 25 см. Он состоит из 2 отделов — подвижного и способного высовываться переднего, овального в поперечном сечении, и жестко зафиксированного заднего. Снаружи язык обильно покрыт слизистой слюной, которую вырабатывают крупные подъязычные железы. От переднего конца каждой железы большой выводной проток подходит к нижней стороне полости рта и делится здесь на ряд более мелких. Благодаря тому, что они открываются вдоль большей части дна ротовой полости, слюна равномерно поступает на длинный язык. Состоит она в основном из гликопротеинов, что делает язык скользким и липким.

Австралийская ехидна (Tachyglossus aculeatus) собирает высунутым языком мучных червей.
Язык втягивается за счет расслабления кольцевых мышц, образующих его правую и левую половины, а также сокращения грудинно-язычных мышц (mm. sternoglossi) — 2 продольных мускулов, залегающих внутри кольцевых мышц и простирающихся от самого кончика языка через его корень до грудины, к мечевидному отростку которой они прикреплены. Высовывание языка осуществляется благодаря частичному сжатию кольцевых мышц и неполному сокращению 2 подбородочно-язычных мышц (mm. genioglossi), которые прикрепляются к корню языка и к нижней челюсти. Поскольку язычное основание неподвижно зафиксировано, сжатие кольцевых мышц меняет форму языка таким образом, что он выдвигается вперед. Этому способствует наличие сосудистого механизма жесткости.
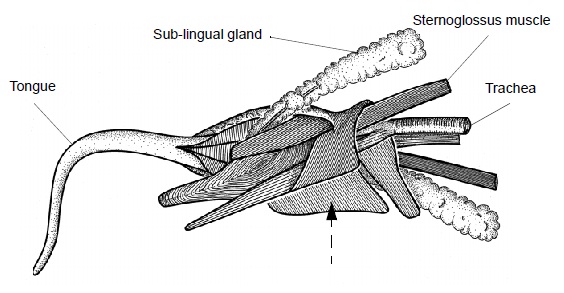
Строение язычной мускулатуры австралийской ехидны (Tachyglossus aculeatus).
Механизм жесткости находится в передней части языка и состоит из центрально расположенной артерии, нерва под ней и сосудистых пространств вверху и внизу. Эти пространства снабжаются кровью из центральной артерии через ряд больших и маленьких каналов. В каждой половине языка синусоиды эректильной ткани объединяются по средней линии перед поперечным синусом, образуя обширное пространство, выстланное эндотелием и содержащее артериальную кровь. Поперечные волокна кольцевых мышц вложены в стенку синуса. Сокращение кольцевых мышц сжимает и искривляет большой синус, повышая давление содержащейся в нем крови. Она вытесняется в верхние и нижние синусоиды, что ведет к быстрому выпячиванию языка. Таким образом, высунутый язык становится жестче за счет быстрого наполнения кровью многочисленных кровеносных сосудов. В этом состоянии он достаточно твердый для того, чтобы проникать в изъеденную термитами древесину.
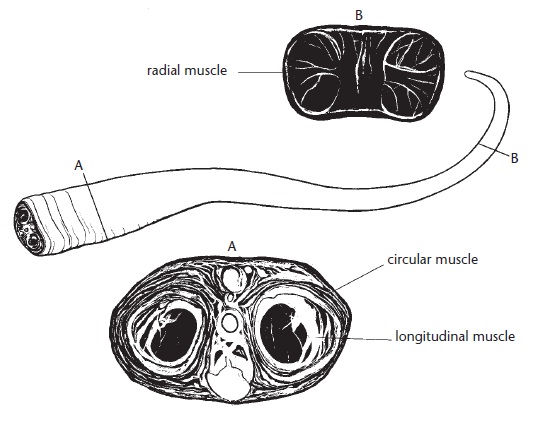
Строение мускулатуры языка австралийской ехидны (Tachyglossus aculeatus). Точки, в которых были сделаны 2 поперечных разреза, обозначены прямыми линиями поперек языка.
Зубы у ехидны отсутствуют на всех стадиях индивидуального развития, но сверху на задней части языка находится кератиновый бугор с большим количеством роговых шипов — преобразованных нитевидных сосочков. Эти шипы направлены назад и имеют высоту около 2,5 мм, постепенно уменьшаясь в размерах по направлению к кончику языка. Над бугром, по небу, расположены 7 поперечных гребней, состоящие из кератинизированного эпителия и тоже несущие мелкие, остроконечные, направленные назад шипы. В эту слегка вогнутую часть неба, образованную задними концами небных костей и крыловидными костями, хорошо вписывается по своей форме язычный бугор. Когда корм достигает заднего конца ротовой полости, бугор прижимается к небу и движется вперед и назад под действием языковой мускулатуры. При этом шипы на верхней поверхности бугра трутся о шипы на гребенчатом небе и таким образом грубо измельчают пищу в однородную массу. Данный процесс сопровождается характерным царапающим шумом, достаточно громким для того, чтобы быть услышанным находящимся поблизости человеком. Кончик языка слегка расширен и имеет на нижней стороне ложкообразное углубление с 10-12 длинными продольными складками. На складках расположены крупные роговые шипы, помогающие подцеплять корм.
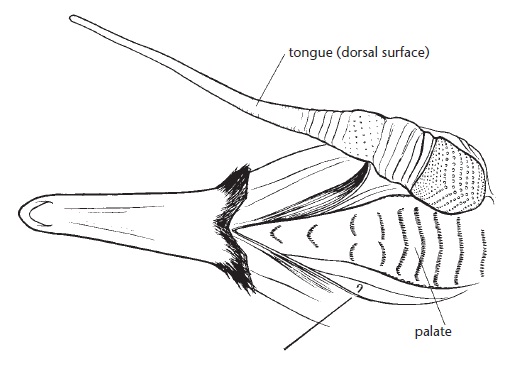
Измельчающий язычно-небный аппарат австралийской ехидны (Tachyglossus aculeatus) (по Augee et al., 2006). Небо открыто, язык отодвинут в сторону и развернут на 180°.
M. detrahens mandibulae, опускающий нижнюю челюсть утконоса, у ехидны практически не функционирует. Обычным шарнирным движениям в нижнечелюстном суставе препятствует продольная краевая связка, простирающаяся вперед примерно до 1,3 см от кончика морды и прикрепляющая ветвь зубной кости к верхнечелюстной и предчелюстной костям. Кроме того, мыщелок нижнечелюстной кости лежит в той же плоскости, что и ее ветвь, а не под прямым углом к ней. Легкое надавливание на кожу, покрывающую угловой отросток зубной челюсти, вызывает ее вращение относительно продольной оси почти на 180°. Поскольку нижняя челюсть несколько изогнута, это вращение приводит к небольшому опусканию ее переднего конца, тем самым приоткрывая ротовую щель. Считается, что это действие вызывается сокращением мышечного пучка, который берет начало в задней части височной мышцы и прикрепляется на внутренней поверхности углового отростка. Сокращение мышцы, прикрепленной к внешней поверхности углового отростка, закрывает рот.
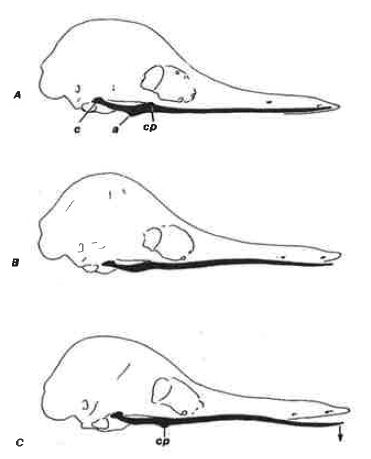
Схема движения нижней челюсти австралийской ехидны (Tachyglossus aculeatus): А — закрытое положение; В — начало вращения; С — полностью повернутая зубная кость и максимально открытое ротовое отверстие; с — мыщелок; а — угловой отросток; ср — венечный отросток.
Пищеварение
Перетертый между языком и небом корм по пищеводу поступает в желудок, имеющий очень своеобразное строение. Он выстлан многослойным кератинизированным эпителием и не содержит пищеварительных желез. В отличие от большинства млекопитающих, желудки которых характеризуются высоким уровнем кислотности, в желудке ехидны кислотность низкая, почти нейтральная (pH = 6,2-7,4). Несмотря на роговую выстилку, стенки желудка достаточно эластичны и могут значительно расширяться, вмещая большое количество проглоченной пищи. Ее измельчение довершают мощные перистальтические движения желудочных стенок, чему способствует случайно захваченный детрит.
За исключением вероятного частичного самопереваривания перетертой массы из насекомых в желудке, все пищеварительные процессы осуществляются в кишечнике. На его внутренних стенках имеются обычные ворсинки и либеркюновы крипты, характерные и для других позвоночных. Снаружи ворсинки одеты в простой столбчатый эпителий, а внутри содержат бокаловидные клетки, секретирующие муцины. Имеются клетки Панета. В начальный отдел тонкой кишки открывается общий выводной проток поджелудочной железы и желчного пузыря. В желчи содержится таурохолевокислый натрий. Также в начале тонкой кишки находятся бруннеровы железы (встречающиеся только у млекопитающих) и вырабатывается щелочный секрет succus entericus.
Слизистая оболочка кишечника взрослой ехидны обладает мальтазной, изомальтазной и трегалазной активностью, но не содержит сахараз и практически не проявляет лактазной активности. Присутствие трегалазы примечательно, поскольку трегалоза (дисахарид из 2 молекул глюкозы) — основной сахар, обнаруженный у насекомых. Также выявлены ферменты фукозидаза и N-ацетил-D-глюкозаминидаза. Это контрастирует с фермантами, присутствующими в слизистой кишечника подсосного детеныша. Они включают некоторые количества лактазы, мальтазы, изомальтазы, нейраминидазы и практически лишенные сахаразной активности, но относительно громадное количества трегалазы, фукозидазы и N-ацетил-D-глюкозаминидазы. Несомненно, это отражает необходимость переваривания 2 основных олигосахаридов, обнаруженных в молоке: сиалиллактозы (4-0-ацетил-N-ацетилнейраминамиллактоза) и фукозиллактозы.
В качестве очевидной компенсации отсутствия отработанного механизма расщепления белков у взрослой особи имеется очень большая тонкая кишка, достигающая приблизительно 3,4 м в длину. Благодаря этому продолжительность пищеварения увеличивается: от поедания порции термитов до выделения экскрементов может пройти более 2 суток. Таким образом, появляется достаточное количество времени для действия пищеварительных ферментов поджелудочной железы и кишечника на мягкие ткани добычи. Большая часть воды реабсорбируется в заднем отделе кишечника. Фрагментированные состоящие из хитина экзоскелеты, а также детрит, не перевариваются и выводятся вместе с калом.
Выделение
В связи с наличием карликовых нефронов без длинных петель Генле, почки ехидны не могут продуцировать высококонцентрированную мочу, как это характерно для ряда пустынных грызунов. По степени концентрации ее моча сопоставима с таковой у кролика и собаки. У животного, лишенного пищи и воды, объем выделяемой мочи невелик, а ее концентрация достигает 2300 мосм/кг воды. Для сравнения, концентрация плазмы крови составляет 281 мосм/кг воды, что приблизительно в 8 раз меньше. Оптимальные показатели веса тела и азотистого баланса фиксировались при температуре воздуха 21-28 °C и его влажности 9,0-15,6 г воды/куб. м. При уменьшении влажности воздуха до 3,3 г воды/куб. м те же самые особи при аналогичном количестве съеденных термитов стабильно теряли вес и выделяли с мочой больше азота. При наличии питьевой воды животные употребляли ее в среднем 40-80 мл в день. Особи, использованные в данных экспериментах, принадлежали к южноавстралийскому подвиду Tachyglossus aculeatus aculeatus. Не исключено, что Tachyglossus aculeatus acanthion из более засушливой центральной области материка переносит недостаток влаги лучше.
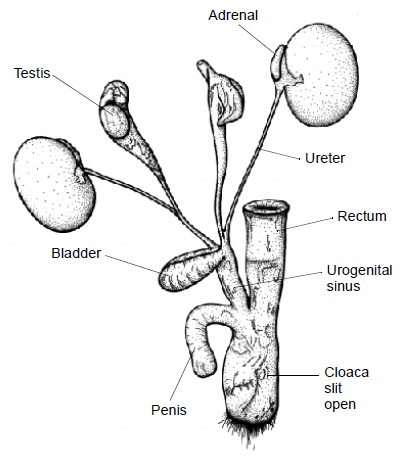
Выделительные и половые органы самца ехидны (Tachyglossus aculeatus). Препуциальный мешок удален, чтобы продемонстрировать пенис.
Испарение воды организмом ехидны сведено к минимуму — на теле отсутствуют потовые железы, а в носовой полости имеется противоточная система теплообмена. Когда воздух вдыхается через рыльце, он проходит через сложный костный лабиринт носовой полости, обеспечивающий его контакт с обширной площадью влажной эпителиальной ткани. Сухой вдыхаемый воздух испаряет влагу из эпителия, тем самым охлаждая стенки полости, и проходит в дыхательные пути глубже уже более теплым и влажным. При выдохе этот влажный воздух возвращается назад вдоль носовых раковин, стенки которых постепенно охлаждаются снаружи. Когда выдыхаемый воздух тоже охлаждается, он насыщается водными парами, которые конденсируются на стенках носовой полости. Во время следующего вдоха эта влага насыщает поступающий воздух и вновь охлаждает полость, продолжая водосберегающий цикл. Помет ехидны относительно сухой. Сохранению влаги также способствует избегание воздействия высоких температур окружающей среды.
Особь весом 3 кг, содержавшаяся в лаборатории при температуре воздуха 25 °C и низком уровне влажности, теряла в сутки 120 г воды: 60 г вместе с мочой, 9 г с фекалиями, 51 г из дыхательных путей и через кожный покров. Согласно расчетам, в подобных условиях животное способно поддерживать положительный водный баланс только за счет поедаемых термитов. Они содержат около 77 % воды, а ехидна потребляет около 147 г термитов в сутки. Это обеспечивает поступление 113 г свободной воды. Распад молекул жира и белка из съеденных термитов дополнительно дает еще 9 г воды, доводя ее общее количество до 122 г взамен утраченных 120 г. Однако по мере повышения температуры окружающей среды и возрастания активности расходы влаги будут увеличиваться. К тому же, замена в рационе термитов муравьями приведет к существенному снижению количества воды, поступающей в организм. Поэтому при свободном доступе к воде ехидна охотно ее пьет. Однако это животное зачастую водится там, где открытые водоемы отсутствуют — по меньшей мере, часть года. В таких местах ехидна слизывает с субстрата утреннюю росу.
Дыхание
Подобно другим однопроходным, легкие несимметричны: правое легкое составляет около 65 % их общего объема.
Частота дыхания в состоянии покоя составляет 5-6 вдохов в минуту. Согласно результатам спирометрии, концентрация углекислого газа, измеренная в самом конце выдоха (альвеолярная), составляет у ехидны 5,3 %, концентрация кислорода — 14,5 %; у тюленя аналогичные показатели составляют соответственно 6,3 % и 11,6 % , у собаки — 5,68 % и 13,6 %. Средний объем вдыхаемого за один раз воздуха составляет 26,9 мл, средний минутный объем — 137 мл/кг массы тела. При уменьшении частоты дыхания минутный дыхательный объем может сокращаться более чем вдвое. И наоборот, при необходимости вдыхания воздуха с высокой концентрацией углекислого газа, минутный дыхательный объем увеличивается, что достигается за счет возрастания объема вдыхаемого за раз воздуха, а не увеличения частоты дыхания.
Объемы вдыхаемого ехидной за раз и за минуту воздуха сопоставимы с этими показателями у плацентарных. Расход вдыхаемого с воздухом кислорода в термонейтральных условиях составляет только 0,13-0,22 мл/г/час, что примерно в 2 раза меньше, чем у плацентарных аналогичных размеров. Впрочем, данное сравнение зависит от конкретного примера; например, ленивец сходного с ехидной веса также потребляет очень мало кислорода. Кроме того, ехидна терпима к значительно повышенному уровню углекислого газа во вдыхаемом воздухе и добровольно не меняет положения, когда его концентрация достигает 6,9 %. По меньшей мере еще одно млекопитающее, кролик (Oryctolagus cuniculus), выдерживает столь же высокий уровень углекислого газа: в воздухе гнездовой камеры, содержащей детенышей, его концентрация составляет 7-14 %.
Изучалась респираторная и сердечная деятельность ехидны, зарытой в землю и дышащей асфиксическим воздухом, создаваемым ее собственным дыханием. После захоронения на глубину около 20 см фиксировалось замедление сердцебиения (брадикардия) до 12 ударов в минуту, значительное снижение периферического кровообращения, некоторое изменение частоты дыхания и устойчивый рост концентрации углекислого газа (гиперкапния) в воздухе вокруг рыльца. Когда уровень углекислоты достигал 10-12 %, почва над животным начинала вздыматься и в ней появлялись трещины. Спустя несколько секунд данная активность прекращалась, но снова возобновлялась через регулярные промежутки времени, что привело к поддержанию во вдыхаемом воздухе примерно 10 % углекислого газа на протяжении всего эксперимента, длившегося 4 часа. Полагают, что благодаря снижению периферического кровообращения экономится кислород для сердца и головного мозга — органов, наиболее чувствительных к его дефициту. В этом отношении сердечно-сосудистая система ехидны напоминает таковую у некоторых ластоногих.
Ехидна хорошо приспособлена к длительному пребыванию под землей. Помимо нечувствительности к углекислому газу, кровь связывает большое количество кислорода и наблюдается хорошо выраженный эффект Бора (0,49). Это гарантирует, что циркулирующая кровь захороненной ехидны обладает достаточным уровнем насыщенности кислородом, чтобы справиться с его нехваткой во вдыхаемом под землей воздухе. Согласно измерения, в плазме захороненного животного значительно возрастает уровень углекислоты при одновременном снижении pH плазмы. Во время эпизодов гипервентиляции, когда ехидна начинала приподнимать почву, брадикардия на короткое время заменялась тахикардией. Помимо гипервентиляции и связанной с ней тахикардии, никаких четко выраженных физиологических адаптаций (в уровне содержания и структуре ферментов, уровне миоглобина, буферной емкости крови и др.) обнаружено не было. Таким образом, указанные приспособления являются вполне достаточными для поддержания метаболического гомеостаза ехидны при ее закапывании в землю на несколько часов подряд. Эта способность помогает в различных критических ситуациях — необходимости укрыться от хищника, пожаре, обрушении норы и т. п.
Кровообращение
Согласно произведенным измерениям, у находившейся в термонейтральных условиях особи сердечный выброс составил 59 мл/кг/мин, артериальное кровяное давление (систолическое/диастолическое) — 123/96 мм рт. ст., ударный объем сердца — 1,4 мл. Частота сердцебиения составляет 50-68 ударов в минуту в состоянии покоя и поднимается до 135-145 ударов в минуту при высокой степени активности.
Подобно другим млекопитающих (за исключением верблюдовых (Camelidae)), эритроциты ехидны имеют вид двояковогнутых безъядерных дисков. Количество гемоглобина в крови довольно высокий — его средний уровень составляет 174 г/л. Как и у утконоса, для эритроцитов характерен крайне низкий уровень содержания аденозинтрифосфата (АТФ) — всего 0,03 мкмоль/мл. Однако столь небольшого количества оказывается достаточно для переноса катионов через эритроцитарные мембраны.
Состав лейкоцитов следующий: нейтрофилы — 15 %, моноциты — 1 %, лимфрциты — 87 %, базофилы и эозинофилы практически отсутствуют (2 последних вида белых клеток, однако, имеются в крови утконоса). Подобно териям, для гранулоцитов характерны сегментированные ядра.
Согласно результатам электрофореза на ацетате целлюлозы, сыворотка крови ехидны содержит 7 четко определенных фракций: aльфа-1-, aльфа-2-, aльфа-3-, бета-1-, бета-2- и гамма-глобулины и, а также альбумин. По своей подвижности они сходны с таковыми в человеческой сыворотке. Бета-1-глобулин (трансферрин) связывает железо для переноса в различные клетки, например ретикулоциты (предшественники красных кровяных телец) и клетки костного мозга. После инъекции антигена Salmonella flagellae в сыворотке были обнаружены иммуноглобины IgM и IgG, имеющиеся также у териев. По своей электрофоретической подвижности, размерам и полипептидной структуре они напоминают, в частности, иммуноглобулины человека. При этом IgG количественно преобладает, а его тяжелая цепь гамма имеет большое сходство с человеческой. Преобладание в сыворотке иммуноглобина IgG в большей мере напоминает эту картину у териев, чем у рептилий.
Нервная система
Головной мозг развит достаточно хорошо и имеет неожиданно крупные размеры. Его средний объем составляет 25 куб. см, что соответствует этому показателю у кошки (Felis silvestris) сходной величины. Обонятельные луковицы очень крупные, их объем составляет около 3,1 % от общего объема головного мозга (у утконоса — только 0,8 %). Древняя кора, или обонятельная доля, у ехидны также относительно больше. Новая кора больших полушарий покрыта довольно многочисленными и глубокими извилинами (гирэнцэфалия), тогда как у утконоса (а также многих сумчатых) извилины полностью отсутствуют. Площадь коры, скрытой в бороздах, составляет 36 % (для сравнения: у кошки (Felis silvestris) — 40 %, у саймири (Saimiri sciureus) — 39 %). Хотя новая кора тоньше, чем у утконоса, в ней содержится больше нейронов, которые «упакованы» вдвое плотнее. Судя по всему, это свидетельствует об относительно более высоком уровне мозговой организации ехидны, а значит о ее более сложной психической деятельности. Кроме того, хорошо выраженные морфологические отличия подтверждают длительную независимую эволюцию ехидны и утконоса.
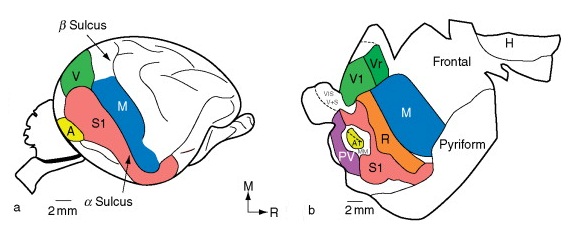
Вверху — схема распределения зон в больших полушариях головного мозга австралийской ехидны (Tachyglossus aculeatus), сбоку (а) и спереди в разрезе (b).
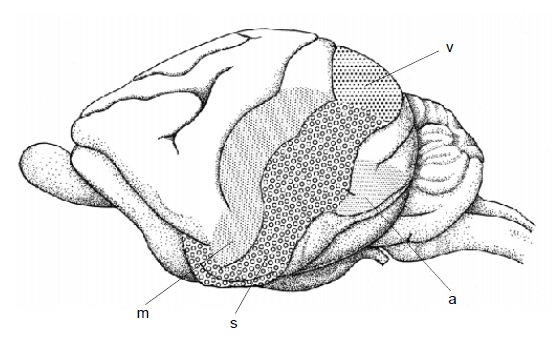
Внизу — области коры головного мозга австралийской ехидны (Tachyglossus aculeatus): s — соматосенсорная и двигательная; m — двигательная; v — зрительная; a — слуховая.
Новая кора занимает 43 % поверхности головного мозга — очень много для насекомоядного млекопитающего с макросматическим мозгом (у человека площадь новой коры составляет 80 %). Еще более необычно то, что предлобный отдел новой коры занимает площадь относительно большую, чем у любого другого млекопитающего — 50 % (для сравнения: у человека — 29 %). Поскольку у высших млекопитающих эта область коры служит для планирования и анализа совершаемых действий, высказано предположение, что ехидна тоже способна рассуждать и выстраивать новую поведенческую стратегию. Она, например, может состоять в том, чтобы не истреблять полностью встречаемые колонии насекомых, что дает им возможность восстановиться и позволяет использовать их для питания в дальнейшем. Наблюдения в Т-образном лабиринте с люками для доступа к пище, визуальные и тактильные тесты, а также способность ехидны удерживать в памяти навыки, приобретенные более месяца назад, свидетельствуют о ее возможностях в обучении и использовании простейших механизмов, вполне сопоставимых с таковыми у крысы (Rattus norvegicus) и кошки (Felis silvestris).
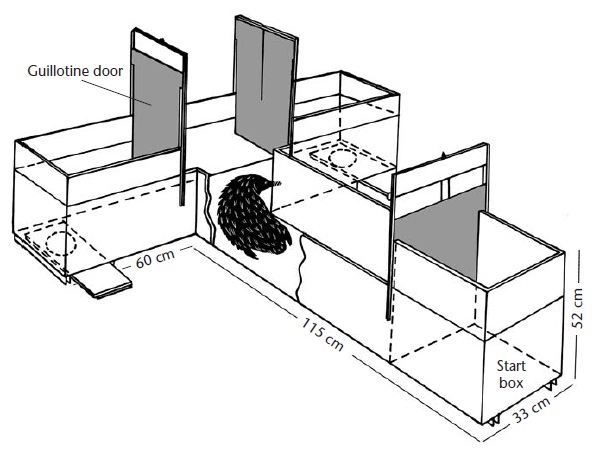
Т-образный лабиринт, используемый для исследования уровня интеллекта австралийской ехидны (Tachyglossus aculeatus).
Корковые зоны для приема и обработки соматосенсорных, зрительных и слуховых импульсов сгруппированы в задней части полушарий (у утконоса они занимают большую часть полушарий, включая лобную область) и отличаются по расположению от таковых у других млекопитающих. Зона, отправляющая двигательные импульсы, частично совпадает с соматосенсорной, а частично простирается дальше вперед. Участок сенсомоторной зоны, обслуживающий рыльце и язык, занимает почти половину всей чувствительной области, но все же существенно уступает аналогичному участку коры утконоса, отвечающему за его клюв.
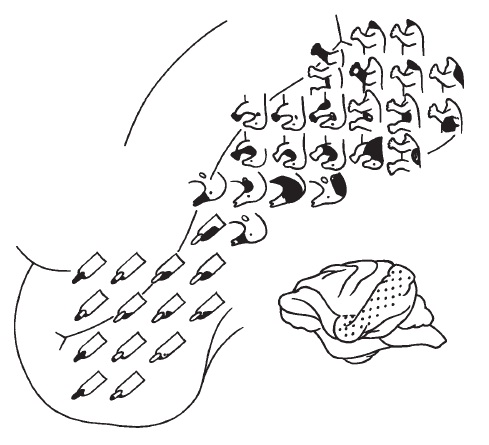
Представительства различных частей тела в сенсорной области коры головного мозга австралийской ехидны (Tachyglossus aculeatus). Обращает на себя внимание обширный участок, отвечающий за рыльце и язык.
Как и у прочих млекопитающих, перераспределение информации, идущей от органов чувств к коре головного мозга, осуществляется таламусом. Идентифицированы 2 архитектонических поля в сенсомоторной области и связанные с ними таламические афференты. Возможно, данные поля гомологичны зонам коры плацентарных, известным как 3а и 3b. Необычной особенностью является относительно большие размеры верхне-передне-бокововых отделов таламуса, которые проецируются на лобную кору. Считается, что этот крупный ядерный регион частично является эквивалентом верхне-бокового ядра териев и обрабатывает обонятельные данные, предназначенные для коры. Данная область мозга тесно связана с той, которая воспринимает чувствительные импульсы, приходящие от механорецепторов рыльца по сильно развитому тройничному нерву. Возможно, увеличение лобных долей ехидны в определенной мере объясняется их обонятельной специализацией.
Под корой больших полушарий залегает тонкая пластинка из серого мозгового вещества — клауструм. Он также присутствует у плацентарных, что может указывать на наличие у них общего с однопроходными предка. Однако существует точка зрения, что клауструм в равной мере характерен и для сумчатых млекопитающих. Мозжечок относительно меньше, чем у утконоса.
Спинной мозг ехидны относительно короче, чем у большинства других млекопитающих. Например, если у человека он заканчивается на уровне I-го или II-го поясничного позвонка, то у ехидны тянется только до VII-го грудного позвонка. Полагают, что это предотвращает чрезмерное растяжение спинного мозга при свертывании животного в защитный шар. Вероятно, растяжению в основном подвержен более эластичный конский хвост (cauda equina) — пучок нервных волокон, исходящий от заднего конца спинного мозга.
Расположение нервных клеток в спинном мозге ехидны аналогично таковому у териев. Примечательно, однако, что волокна двигательного корково-спинномозгового пирамидного пути перекрещиваются и переходят на противоположную сторону ствола головного мозга в варолиевом мосте. Это отличает ехидн от всех других млекопитающих; насколько известно, у утконоса данный морфологический аспект пока не изучался. У большинства териев волокна пирамидного пути перекрещиваются в продолговатом мозге. Интересно, впрочем, что относительно высокий перекрест также характерен для небольшого числа узкоспециализированных млекопитающих, включая броненосцев и панголинов, но с чем это связано — пока неясно. Пирамидный путь ехидны проходит по большей части спинного мозга, что является отличительной чертой продвинутой нервной системы, наблюдаемой также у приматов и хищных, и позволяет коре напрямую управлять двигательными нейронами вплоть до задних конечностей и хвоста. У ежей и тупай волокна пирамидного пути перекрещиваются гораздо выше, чем у ехидны.
Долгое время считалось, что у ехидны отсутствует фаза быстрого сна. Затем было установлено, что мозг спящего животного все-таки проходит эту фазу, но ее продолжительность зависит от температуры окружающей среды. Так, при 25 °C фаза быстрого сна занимает около 15 % его общего времени, а при температуре 15-20 °C и 28 °C она значительно сокращается или полностью исчезает. Приуроченность фазы быстрого сна к невысокой температуре тела может указывать на древность ее происхождения.