Органы чувств
Зрение у ехидны довольно слабое. Глазное яблоко достигает 8-9,5 мм в диаметре и имеет грубо сферическую форму. Его движениями в глазнице управляют 6 глазных мышц. Имеется тонкая мышца, которая тянется от древнего места прикрепления на передне-носовой поверхности глазницы до места присоединения к глазному яблоку и срастается здесь с второй мышцей, которая подходит к месту прикрепления всего в нескольких миллиметрах впереди от глубоко расположенной точки присоединения 4 прямых мышц. У утконоса присутствует только этот длинный мускульный пучок, который, как у териев, возникает из прямой мышцы и становится сухожильным до достижения своего места прикрепления, причем последнее построено из хряща, а не мягкое, как у ехидны. По-видимому, развитие длинной верхней косой мышцы было связано с появлением бинокулярного зрения. Косые мышцы нужны для правильного обзора объекта, когда голова повернута в сторону, чтобы изображение попадало в одну и ту же часть сетчатки обоих глаз. Мигательная перепонка отсутствует. Подобно утконосу, в задней части склеры залегает чашевидное образование из хряща, имеющее поддерживающую функцию.
Маленькая уплощенная роговица занимает примерно 1/6 окружности глазного яблока. Поверхностные слои роговичного эпителия кератинизировались, как это наблюдается у муравьедов и некоторых питающихся муравьями броненосцев. Затвердевшая поверхность роговицы призвана защитить глаз от укусов и едких веществ, выделяемых муравьями и термитами, а также от попадания мусора во время рытья. Кроме того, глазное яблоко оказывается в определенной мере защищенным от случайных уколов иголками при свертывании ехидны в шар или во время ее общения с сородичами. Конечно, все это не препятствует в нужные моменты дополнительно прикрывать глаза толстыми веками. Мышца-расширитель зрачка (m. dilatator pupillae) отсутствует, имеется лишь крупный суживатель зрачка (m. sphincter pupillae), расположенный на краю радужки и окруженный несколькими ее пигментированными слоями. Эта пигментация придает глазу ехидны характерный черный цвет.
У ехидны самый плоский хрусталик среди всех млекопитающих. Индекс его уплощенности (отношение диаметра хрусталика к его толщине) составляет 2,75. К этому показателю приближаются только некоторые высшие приматы включая человека, с индексом уплощенности приблизительно 2,7. Подобная форма хрусталика необходима для четкого видения удаленных объектов. Однако внутренняя ресничная мышца (m. ciliaris), имеющаяся у териев и влияющая на кривизну хрусталика, отсутствует. Следовательно, с помощью хрусталика на близких объектах глаз фокусироваться не способен. В связи с этим считается, что внешние мышцы глазного яблока сжимают и удлиняют его таким образом, что изображение близкого объекта попадает на сетчатку. Это подтверждается тем, что во время еды или при приближении к крупному объекту глаза ехидны несколько выступают из глазниц.
Сетчатка представляет собой однородный слой, лишенный сосудов и углублений. Кислород и питательные вещества поступают в нее из сосудов подстилающей хориоидеи, или собственно сосудистой оболочки. Такая полная трофическая зависимость сетчатки от хориоидеи характерна для избегающих яркого света позвоночных. Однако известно, что ехидна проявляет активность как днем, так и ночью. В ее сетчатке содержится 85-90 % палочек и 10-15 % двойных колбочек с капельками жира (для сравнения: в человеческом глазу на колбочки приходится лишь около 5 % фоторецепторов). В расположении светочувствительных клеток наблюдается определенная специализация. Колбочки концентрируются в центральной части сетчатки в гораздо меньшей степени, чем у териев. Соотношение центральных колбочек к периферическим составляет только 2 : 1, тогда как у кошки оно достигает 7,5 : 1, а у обезьяны — 100 : 1. В ходе лабораторных исследований показано, что ехидна различает близкие ярко окрашенные предметы. Неизвестно, способна ли она к цветовому восприятию, но определенно может различать черный и белый цвета, а также горизонтальные и вертикальные полосы.
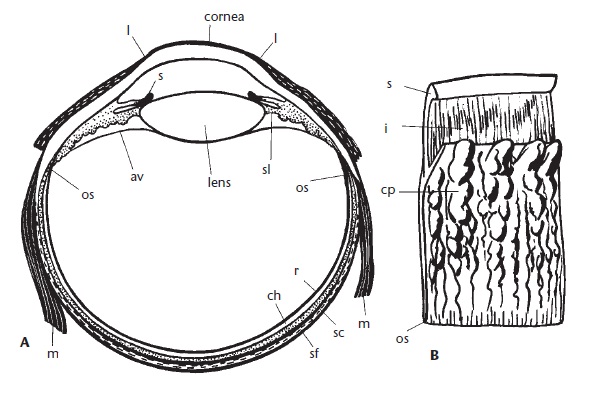
Строение глаза австралийской ехидны (Tachyglossus aculeatus): А — поперечный разрез глаза; B — сегмент задней поверхности радужки и цилиарного тела; av — передняя поверхность стекловидного тела; ch — сосудистая оболочка; cornea — роговица; cp — ресничный отросток; i — радужка; lens — хрусталик; l — боковая граница роговицы; m — прямая мышца; os — передняя граница сетчатки; r — сетчатая оболочка; s — сфинктер; sc — склеральный хрящ (черный); sf — волокнистый слой склеры; sl — связка, подвешивающая глазное яблоко.
Идущие из сетчатки нервные волокна образуют зрительный нерв. В нем насчитывается около 15 000 волокон, что немного по сравнению с приматами, но вдвое больше, чем у большого бурого кожана (Eptesicus fuscus). В оптической хиазме перекрещиваются почти все зрительные волокна и лишь около 1 % их продолжает идти по той же стороне мозга. Казалось бы, столь малая доля неперекрещивающихся волокон не способствует построению в мозгу трехмерной зрительной картинки. В то же время, оптические оси глаз ехидны отклоняются от срединной линии черепа всего на 25°, что указывает на существенное перекрывание ее зрительных полей. Также можно предположить, что несмотря на небольшое количество неперекрещенных зрительных волокон и отсутствие мозолистого тела, между полушариями мозга существуют другие пути обмена визуальной информацией. Таким образом, степень развития бинокулярности зрения окончательно не установлена. В целом зрительные способности ехидны сопоставляют с таковым у серой крысы (Rattus norvegicus). Очевидно, зрение не играет существенной роли при определении пищи или опасности — ехидна зачастую активна в темноте, ее добыча обычно скрыта от глаз и в природе наблюдались успешно выживающие слепые особи.
Напротив, слух для ехидны имеет большое значение. Зачаточная ушная раковина состоит из недифференцированной пластины хряща, прилегающей к коже и мускулатуре. Форма раковины овальная, с длинной верхне-нижней осью. На нижнем конце раковины боковые края ушного хряща образуют куполообразную структуру. Таким образом, по своим очертаниям ухо напоминает треугольник с основание вверху и вершиной внизу. Наружный слуховой проход состоит из хрящевых колец, сочлененных наподобие колец трахеи. Проход сильно изогнут. Его наружный отрезок под прямым углом отходит от ушной раковины и, вытягиваясь проходит вперед. Более короткие средний и внутренний отрезки поворачивают внутрь и вверх соответственно, в конце достигая отверстия в черепе. Проход имеет наибольшую ширину на ушном конце и наименьшую — на черепном. Полагают, что связанные с наружным ухом мышцы закрывают его, прижимая ушную раковину к внутренним стенкам наружного слухового прохода. Во время передвижения наружное ухо обычно закрыто, но при приближении к незнакомому объекту прилегающие к ушной щели иглы поднимаются, тем самым открывая наружный слуховой проход.
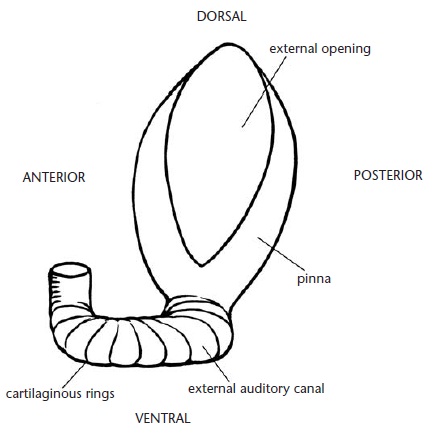
Хрящевое основание левой ушной раковины и наружного слухового прохода австралийской ехидны (Tachyglossus aculeatus).
То обстоятельство, что ушная раковина не выступает наружу, уменьшает возможность ее травмирования при копании грунта и захвата хищником. Волоски, густо растущие в ушной раковине и концевом отрезке слухового прохода, существенно снижают риск попадания сюда мусора и поедаемых насекомых. С другой стороны, это обеспечивает относительно защищенную среду обитания для клещей. Внутримышечное расположение и сравнительная простота строения ушной раковины уменьшают ее способность направлять звуковые колебания в слуховой проход. Последний поворачивает под 3 прямыми углами, располагаясь в 3 различных пространственных плоскостях. Эта изогнутость наряду с узостью прохода могут предохранять его от звуков высокой интенсивности. Проход представляет собой полностью внечерепную структуру, поэтому передача звука через кость в нем незначительна. Таким образом, звуковые помехи воздушной среды глушатся, а чистота звука, передаваемого через костную толщу во внутреннее ухо, возрастает.
Наружный слуховой проход заканчивается на барабанной перепонке, которая отделяет наружное ухо от полости среднего уха, окруженной костями черепа. Подобно другим млекопитающим, через среднее ухо звук проводится цепочкой из 3 маленьких косточек, расположенных в следующем порядке: молоточек, наковальня и стремя. Основание стремени расширяется в округлую пластинку, которая упирается в мембрану овального окна улитки. Данное окно, однако, имеет не овальную, а круглую форму. Молоточек достигает крупных размеров и жестко сращен со следующей костью в цепочке, наковальней. С помощью дополнительного отростка (processus folii) молоточек и наковальня плотно прикреплены к каменистой кости. Барабанная кость полукольцевидной формы плотно присоединяется, а часто даже срастается с отростком (processus gracilis) молоточка. Воздействие на молоточек тонким стержнем показывает, что достаточно сильное давление вызывает лишь незначительное смещение костей среднего уха. Такая устойчивость цепи слуховых косточек обусловлена очень плотным соединением наковальни и молоточка между собой и с черепом. Стременное отверстие у ехидны не отмечено ни на одной из стадий развития, стременной артерии также нет.
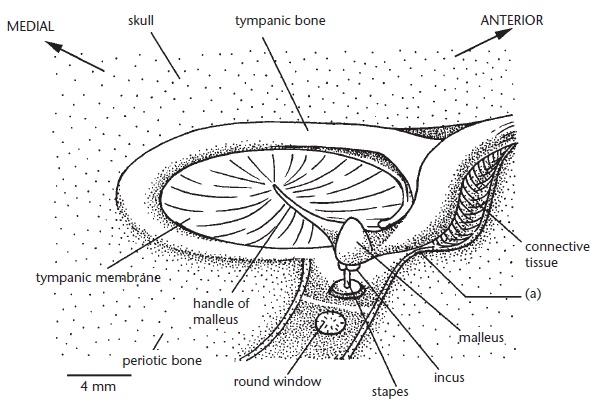
Строение среднего уха австралийской ехидны (Tachyglossus aculeatus) в верхне-боковой проекции.
Были измерены скорость и амплитуда движения стремени при воздействии на наружный слуховой проход звуками в частотном диапазоне от 100 Гц до 24 КГц и с постоянной интенсивностью 100 дБ. Для ехидны оба показателя на низких частотах оказались существенно меньшими, чем аналогичные данные, полученные для морской свинки (Cavia porcellus) и ящерицы дракона (Draco volans). Только после того, как применяемая частота звуковых колебаний была увеличена до 6 кГц, стремя продемонстрировало реакцию, соответствующую таковой у других животных. Судя по всему, малая степень функциональности в низкочастотном диапазоне вызвана значительной громоздкостью звукопроводящего аппарата. Жесткое срастание молоточка с наковальней и прилежащей стенкой черепа способствует тому, что он функционирует скорее как продолжение височной кости, служащей опорой для барабанной перепонки, а не в качестве настоящей слуховой косточки. Это сильно отличает его от легко подвешенного молоточка большинства плацентарных млекопитающих.
Таким образом, среднее ухо ехидны малоэффективно для проведения воздушных звуковых колебаний. С другой стороны, строение слуховых косточек подразумевает повышенную чувствительность к вибрациям, проводимым телом и передаваемых в ухо через толщу черепных костей. При этом легкие постукивания по различным частям тела вызывают лишь незначительное колебание мембраны круглого окна, в противоположность отчетливо выраженному отклику при постукивании по рыльцу. Это позволяет предположить, что часть слуховой информации ехидна получает через рыльце и нижнюю челюсть, когда роется ими в почве или гнилой древесине либо просто касается последних. Неинвазивные методы, используемые для измерения чувствительности слуха у легко анестезированных пойманных ехидн показали, что они улавливают звуковые колебания довольно узкого диапазона, но чувствительность в этом диапазоне сопоставима с чувствительностью у таких типичных териев, как кролик (Oryctolagus cuniculus) и песчанка (Psammomys obesus).
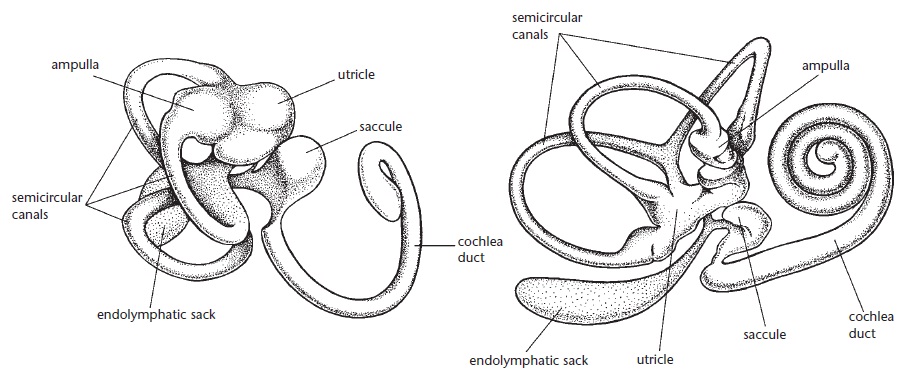
Строение правого внутреннего уха австралийской ехидны (Tachyglossus aculeatus) (вверху) и типичного плацентарного млекопитающего (внизу).
В отличие от спирально свернутой улитки внутреннего уха большинства териев, улитка ехидны имеет вид бананообразной кривой со спиральным поворотам на 0,75 на конце (у человека улитка делает 2,75 оборота). Но несмотря на рептильный внешний вид, внутренняя структура улитки вполне сходна с таковой у териев. Находящийся здесь кортиев орган достигает приблизительно 7,6 мм в длину — намного меньше, чем у эвтериев. В нем содержится примерно столько же внутренних волосковых клеток (2700), что и у человека (3000), но при этом у ехидны выше число клеточных рядов и меньше плотность размещения в них клеток. Судя по всему, внутренние волосковые клетки обеспечивают большую часть чувствительности кортиева органа. Число внешних волосковых клеток ехидны (5050) значительно уступает таковой у человека (12 000), но сопоставимо с их числом у живущего под землей голого землекопа (Heterocephalus glaber). Судя по всему, лучше развитые у большинства териев внешние волосковые клетки улавливают ту часть звукового диапазона, которая не особенно важна для ехидны.
Установлено, что кортиев орган ехидны наиболее чувствителен к звуковым колебаниям частотой около 5 кГц. Они являются низкими по сравнению с диапазоном частот, максимально стимулирующим внутреннее ухо териевых млекопитающих, но вполне сопоставимы с таковыми у утконоса. Слуховая чувствительность подобного рода может идеально подходить для улавливания шумов и вибраций, которые под поверхностью грунта производятся крупными скоплениями муравьев и термитов и которые через кости рыльца и черепной коробки передаются во внутреннее ухо. Таким образом, особенности слуха ехидны способствуют лучшему обнаружению пищи.
Отолитовый аппарат внутреннего уха, выполняющего функцию датчика силы тяжести, развит очень хорошо. Как и у териев, он представлен ампулами 3 полукружных каналов и макулами овального и круглого мешочков. Площадь макулы овального мешочка составляет 4,5 кв. мм (для сравнения: у человека — 4,29 кв. мм, у морской свинки — 0,54 кв. мм). Число волосковых клеток в макуле овального мешочка является наибольшим среди млекопитающих — 56 3000 (человек — 33 100, саймири — 11 360, морская свинка — 9260). Кроме того, подобно рептилиям и в отличие от териев у ехидны (и остальных однопроходных) имеется макула улитки — узкая полоса чувствительных клеток в концевом отделе улитки. Такое выраженное развитие у ехидны вестибулярного аппарата может быть связано с необходимостью ориентироваться в подземных ходах, а также отображать ее происхождение от маневренных водных предков.
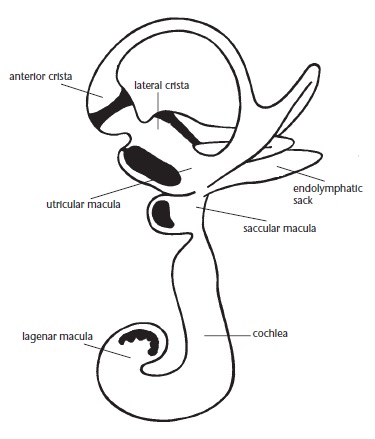
Расположение вестибулярных органов правого внутреннего уха австралийское ехидны (Tachyglossus aculeatus) во внутренней проекции.
Кончик рыльца богато иннервирован ответвлениями большого тройничного нерва, по которым в головной мозг непрерывно передается информация о состоянии окружающей среды. Нервные волокна, связанные на концах со слизистыми железами и выступающие через микроскопические поры кожи, действуют в качестве электрорецепторов. В общей сложности их насчитывается порядка 400 — в среднем около 7 на 1 кв. мм кожной поверхности. Эти рецепторы способны улавливать электрическое напряжение всего в 1,8 мВ/см — величину в 1000 раз меньшую, чем способен воспринять человек. С их помощью ехидна улавливает колебания электрического поля, возникающие при сокращении мускулатуры мелких животных. Постоянно влажный кончик рыльца обеспечивает прием из субстрата электромагнитных колебаний, производимых находящейся в нем добычей. Однако в сухих местностях, которые в основном и населяет ехидна, это становится возможным только в сырую погоду, например когда идет дождь. Не исключено, что при благоприятном стечении обстоятельств электрорецепторы могут не только определять местонахождение добычи, но и показывать направление ее движения. С другой стороны, остаточные электрорецепторы ехидны могут являться наследием более водных предков и не играть в ее жизни заметной роли.
Полагают, что электрорецепторы представляют собой преобразованные эккриновые железы с иннервированными выводными протоками. Железа состоит из извитого секреторного отдела, лежащего в толще кожи (дерме), и поднимающегося к кожной поверхности выводного протока. Отдел железы в наружном слое кожи (эпидермисе) имеет булавовидное расширение, которое образуется в результате впячивания эпидермиса. Чувствительные нервные окончания, поднимающиеся из смежной сети кожных нервов, проникают в эпидермальный слой и заканчиваются рядом с клетками, сгруппированными в булавовидном расширении. Идентифицированы 2 типа нервных окончаний, хотя их конкретные функции неизвестны. Также остается неясным, каким образом слабые электрические токи возбуждают нервные окончания. Помимо этого имеются иннервированные эпидермальные ямки без видимых связанных с ними желез, которые также могут выполнять электрорецепторную функцию. Эти структуры распределены позади и по бокам от ноздрей, в области с электрочувствительными участками. Электрорецепторы окружены обычными эккриновыми железами, которые не иннервированны и производят серозно-слизистый секрет. Возможно, он обеспечивает лучший електоконтакт с почвой.
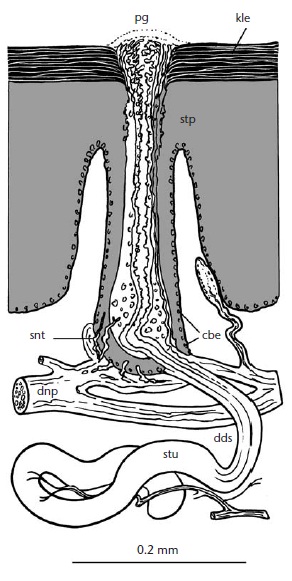
Схема строения электрорецептора австралийской ехидны (Tachyglossus aculeatus): pg — пора железы; kle — ороговевающий слой эпителия; stp — эпидермальная прямая часть протока; snt — чувствительные нервные окончания, примыкающие к протоку; cbe — булавовидное расширение эпителиального сосочка; dnp — дермальное нервное сплетение; dds — извитая дермальная часть протока; stu — секреторная трубочка.
На рыльце также расположены механорецепторы, несколько менее дифференцированые, чем у утконоса. Каждый рецептор состоит из столбца уплощенных, остроконечных клеток, прочно соединенных десмосомами с мембранами соседних клеток. Механорецептор дифференцирован на центральный и периферический отделы. В центральном отделе концентрация десмосом выше. Размеры механорецепторов варьируют на разных участках кожи, но обычно они достигают около 300 мкм в длину и 50 мкм в диаметре. При хорошем освещение можно увидеть куполообразные конусы — кончики рецепторов, возвышающиеся над кожной поверхностью примерно на 25 мкм. Вдоль глубже расположенных 2/3 длины рецептора он отделен от соседнего эпителия посредством выпячивания гораздо более свободно расположенных клеток соединительной ткани, образующих дермальный сосочек. Полагают, что благодаря такому строению рецептор обретает относительную подвижность в вертикальном направлении.
С механорецептором связаны нервные окончания 3 типов, реагирующие на поверхностное давление: цепочки чувствительных пузырьков, клетки Меркеля и тельца Паччини. Этот замечательный комплекс рецептора иннервируется приблизительно 10 миелинизированными аксонами, поднимающимися из соседнего дермального сплетения. Цепочки чувствительных пузырьков расположены как в центральном, так и в периферическом отделах рецептора. Их количество колеблется между 10 и 24 в зависимости от размера рецептора. Цепочка чувствительных пузырьков — это немиелинизированный аксон, поднимающийся к эпидермальному конусу рецептора. Аксон обладает рядом уплощенных округлых расширений, по виду напоминающих нанизанные на нить жемчужины. Срединная цепочка чувствительных пузырьков окружена 2 слоями кератиноцитов, а эти клетки в свою очередь окружены параллельными цепями периферических пузырьков. Ряды вакуолей в цитоплазме кератиноцитов проходят параллельно между пузырьками центральной и периферических цепей.
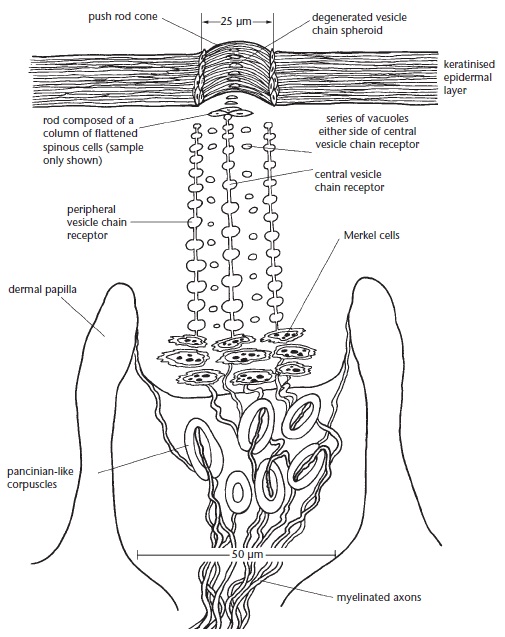
Схема строения механорецептора австралийской ехидны (Tachyglossus aculeatus).
Вероятно, чувствительные пузырьки являются специализированными клетками Меркеля. Возможно, их расположение в центре и на периферии способствует определению направления действия стимула на конус механорецептора. Эта функция может быть облегчена за счет увеличения гибкости тканей из-за наличия рядов вакуолей и снижения плотности десмосом в кератиноцитах, расположенных между центральной и периферическими цепочками пузырьков, тем самым способствуя их дифференцированному смещению. В основании механорецептора 12-26 клеток Меркеля образуют до 3 слоев и иннервируются примерно 4 аксонами. В дерме под механорецептором находится 5-8 телец Паччини, иннервируемых 2-3 аксонами. По строению и присущей функции механорецепторы рыльца ехидн в значительной мере напоминают чувствительный орган Эймера, расположенный на кончике мордочки плацентарных кротовых (Talpidae).
Плотность размещения механорецепторов варьирует от 30 до 40 на 1 кв. мм, их количество постепенно увеличивается по направлению к кончику рыльца. Толстая ороговевшая кожа последнего достаточно жесткая для того, чтобы противостоять травмам во время копания рыльцем твердой почвы и при использовании его в качестве рычага для разламывания трухлявой древесины. В то же время, прогибающиеся под поверхностным давлением конусы рецепторов сигнализируют о малейших и тонко различаемых прикосновениях нижележащим нервным окончаниям, по которым информация передается в головной мозг. Механорецепторы способны сообщать даже о текстуре поверхности жертвы для идентификации конкретной части ее тела. Данная способность может оказаться ценной, если добыча владеет какой-нибудь вредной для ехидны защитной стратегией. Помимо этого, на коже рыльца залегают терморецепторы, одни из которых реагируют на понижение, а другие — на повышение ее температуры. Сигналы, происходящие от рыльца и языка, принимает почти половина сенсорной области новой коры.
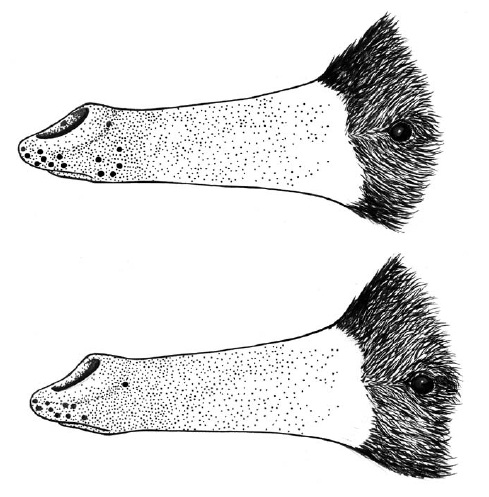
Распределение механорецепторов и электрорецепторов на рыльце австралийской ехидны (Tachyglossus aculeatus). На верхнем рисунке показано расположение слизистых секреторных желез (крупные черные точки), определенное путем микроскопического исследования участков кожи. Нижний рисунок демонстрирует расположение 10 электрочувствительных пятен, определенное посредством слабой электрической стимуляции кожной поверхности. Мелкие точки в обоих случаях обозначают расположение на рыльце осязательных рецепторов.
Во время экскурсий, совершаемых преимущественно в темное время суток, ехидна полагается в первую очередь на превосходно развитое обоняние. Обонятельный эпителий покрывает очень большую площадь — 7 вертикально расположенных решетчатых pаковин (ethmoturbinalia), а также набор носовых (nasoturbinalia) и верхнечелюстных (maxilloturbinalia) раковин. Продырявленную пластинку (lamina cribrosa) пронизывает значительное количество мелких отверстий для прохождения нервных волокон к крупным обонятельным луковицам. По количеству данных волокон ехидна в 13 раз превосходит утконоса, что предполагает относительно большую важность обоняния в ее жизни. Судя по всему, оно играет главную роль в поисках добычи. Впрочем, наличие какого-либо специфического вида химической чувствительности (например, по отношению к муравьиной кислоте) у ехидны не обнаружено. Нюх также необходим для обнаружения полового партнера в период спаривания. Кроме того, чувство обоняния может иметь решающее значение для недавно вылупившегося детеныша, разыскивающего молочные поля на брюхе матери. Об этом свидетельствуют его сильно увеличенные ноздри и полностью сформированный обонятельный тракт, тогда как другие органы чувств еще не развиты (глазные щели не прорезались, а молоточек является частью нижней челюсти).
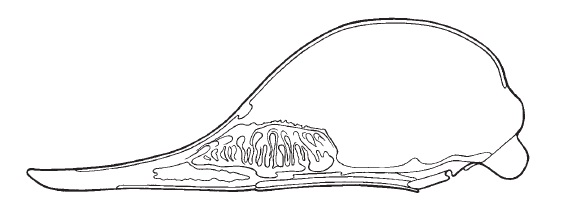
Продольный разрез черепа австралийской ехидны (Tachyglossus aculeatus), демонстрирующий носовую полость и изгибы решетчатых раковин.
На верхней поверхности языка позади кератинового бугра с шипами V-образно расположены 2 щели. Они ведут в глубокие бороздки, на дне которых находятся желобовидные сосочки, содержащие на своей поверхности вкусовые луковицы. Кроме того, многочисленные листовидные сосочки, также снабженные вкусовыми луковицами, располагаются по бокам кератинового бугра и позади его. Таким образом, воспринимающие вкус рецепторы сосредоточены в ротовой полости именно в том месте, где происходит измельчение пищи.