Гормоны
Гипофиз ехидны по своему строению очень сходен с таковым у утконоса и рептилий. В его передней части — аденогипофизе — установлено присутствие лютеинизирующего и адренокортикотропного гормонов.
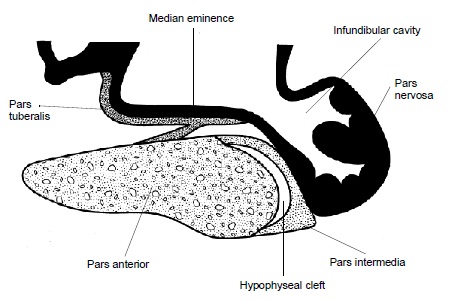
Срединный продольный разрез гипофиза австралийской ехидны (Tachyglossus aculeatus).
Надпочечники маленькие, достигающие около 0,9 см в длину и 0,4 см в ширину. Основными кортикоидами, секретируемыми их стероидогенными клетками, являются кортизол, кортикостерон и альдостерон. Скорость секреции глюкокортикоидов, кортикостерона и кортизола крайне низка, как и плазменный уровень кортикостерона. Эти факты в совокупности с наблюдениями над содержавшимися в неволе ехиднами с удаленными надпочечниками свидетельствуют о том, что секреторная деятельность данных желез не особенно важна для нормального обмена веществ. Тем не менее, подвергшиеся адреналэктомии особи, помещенные в температурную среду 4-10 °C, даже при наличии корма становились вялыми спустя 12-28 часов. Вместе с тем, они начинали вести себя нормально после впрыскивания им кортизола. Введение глюкозы тоже предотвращало снижение температуры тела у этих животных. Все это говорит о том, что действие глюкокортикоида у ехидн, как и у плацентарных, заключается в стимуляции печеночного глюконеогенеза из белка и, следовательно, в восполнении энергетических резервов углеводов, истощенных стрессом.
Впрыскивание в кровяное русло адренокортикотропного гормона (АКТГ) на протяжении 4-дневного периода увеличивает уровни кортизола и кортикостерона, но существенно не влияет на содержание в крови прочих кортикостероидов. У контрольных животных гомогенаты надпочечников конвертируют прогестерон в 11-дезоксикортикостерон и 11-дезоксикортизол, но после введения АКТГ основным продуктом гомогенатов становится кортикостерон. Эти эффекты АКТГ-стимуляции сходны с теми, что наблюдаются у плацентарных.
В искусственных условиях после двусторонней адреналэктомии ехидна может существовать неопределенно долго без развития нарушений электролитного и углеводного обмена. Это свидетельствует о том, что минералокортикоид, альдостерон, играет лишь незначительную роль в регуляции баланса плазменных электролитов. Действительно, альдостерона у ехидны секретируется мало и его уровень в плазме очень низкий. Введение АКТГ и ангиотензина II, однако, приводит к заметному увеличению этих показателей. Возможно также, что отсутствие надпочечников компенсируется высоким содержанием в рационе натрия.
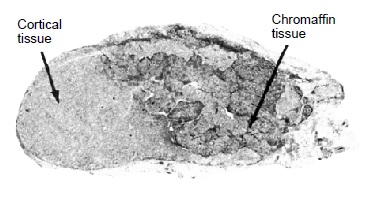
Срединный продольный разрез правого надпочечника австралийской ехидны (Tachyglossus aculeatus).
Как и у утконоса, поджелудочная железа дискретная. Факт хранения инсулина в ней не зафиксирован, однако быстрое возвращение уровня сахара в крови к нормальному в тестах на толерантность к глюкозе свидетельствует о том, что реакцией на гипергликемию является быстрое выделение накопленного инсулина. Инъекция экзогенного инсулина вызывает у ехидны гипогликемию. Хотя инсулин оказывает мощное действие на углеводный, белковый и жировой обмен, антагонистический эффект глюкокортикоидов, который они оказывают на этот процесс, является минимальными.
Тимус, щитовидная и паращитовидные железы расположены жировой ткани позади грудины. Щитовидная железа демонстрирует обычный набор фолликулов, покрытых 1 слоем секреторных клеток, окружающим центральную коллоидную массу. Уровень тироксина в сыворотке (16 нмоль/л) примерно такой же, как у многих сумчатых, но ниже, чем у большинства плацентарных. Щитовидные железы особей, находящихся в оцепенении, сильно гипертрофируются и выглядят как отдельные железы. Как ни странно, уровень тироксина при этом значительно понижен. Разрушение щитовидной железы путем инъекции йода-131 не приводит к существенному снижению метаболизма или температуры тела. По-видимому, низкий уровень гормональной секреции обусловливает замедленный обмен веществ ехидны. По данному критерию она разительно отличается от утконоса.
Такие лимфоидные органы, как селезенка, тимус, аппендикс, миндалины, лимфатические узлы и пейеровы бляшки занимают анатомическое положение, сходное с таковым у териев. Подслизистые разрастания кишечных желез в лимфоидные скопления, лимфоэпителиальные железы, имеются в подвздошной кишке и подобных структурах в толстой кишке и клоаке. Данные образования обладают значительным сходством со стурктурами, обнаруженными в фибрициевой сумке молодых птиц.
Ехидна демонстрирует как как первичные, так и вторичные (анамнестические) реакции на инъекцию антигена Salmonella flagellae путем продуцирования антител IgM и IgG. Хотя анамнестический ответ и может быть выведен, его величина неустойчива и всегда намного меньшая, чем у плацентарных. Анамнестический иммунный ответ ехидны несколько превосходит наблюдаемый у рептилий.
С семенниками самца происходят сезонные изменения. Так, на юго-востоке Австралии в период с октября по март вес семенников составляет 1-3 г/кг веса тела, а средний диаметр семенных канальцев колеблется от 70 до 150 мкм. В начале апреля вес семенников может достигать 8 г/кг. В апреле - мае в них начинается сперматогенез. Наибольшего веса семенники и диаметра семенные канальцы достигают в августе и сентябре. Максимальная зафиксированная масса семенников составила 17 г/кг (обычно она находится в пределах 15 г/кг), а самый большой средний диаметр канальцев — 370 мкм. После этого семенники начинают резко регрессировать и к концу ноября опять сокращаются до размера фасолин.
Наряду с увеличением семенников и расширением семенных канальцев, меняется их гистология. В период покоя летом и осенью канальцы лишены просветов и заполнены сперматогониями, перемежающимися с клетками Сертоли. Сперматогенез начинается в мае: митотическое деление сперматогониев приводит к появлению поколения первичных сперматоцитов. К концу июня канальцы значительно увеличиваются в размерах, в них содержатся первичные и вторичные сперматоциты, сперматиды и появляются первые признаки просветов. В конце июля - середине августа процесс созревания заканчивается, в уже расширенных просветах канальцев содержатся сперматозоиды. В сезон размножения у самцов в плазме крови возрастает уровень тестостерона.
Лактация
Каждая из 2 млечных желез состоит из 100-150 булавовидных долек, связанных соединительной тканью, которая гипертрофируется в период беременности и высиживания яиц. Каждая долька на конце расширяется, а поскольку все они сходятся к относительно небольшому млечному полю, железа имеет вид веера или полумесяца размером около 5 × 2,5 см. Увеличение желез способствует набуханию вышележащей кожной складки, которая формирует выводковую сумку. Внутри каждая долька представляет собой ряд слепо заканчивающихся толстостенных извитых канальцев, лишенных просветов. К моменту вылупления детеныша большинство канальцев получают просветы и расширяются с образованием тонкостенных альвеол. Дольки желез открываются отдельными порами на млечных полях. Каждая пора снабжена специализированной волосяной сумкой, структурно отличной от сумок обычных волос.
Внутри альвеолы выстланы клетками эпителия, секретирующими различные компоненты молока. Их основания примыкают к миоэпителиальным клеткам и базальной мембране. Миоэпителиальные клетки представляют особый интерес, поскольку также присутствуют в млечных железах териев. Они имеют форму нитей и образуют на поверхности альвеол сеть. В цитоплазме этих клеток содержатся мускульные нити, которые сокращаются под действием окситоцина. Это сокращение деформирует альвеолы, что приводит к увеличению в них внутреннего давления и вытеснению молока в канальцы, а оттуда — наружу. Окситоцин рефлекторно секретируется гипофизом самки в ответ на сосательный стимул, который осуществляется ее детенышем. При искусственном введении окситоцина наблюдается аналогичный эффект.
Млечные железы продолжают увеличиваться и ко времени достижения детенышем длины 10 см и веса 90 г составляют около 6-8 × 3 см. Однако после завершения молочного выкармливания они сокращаются в размерах и регрессируют от альвеолярного состояния до плотных канальцев, лишенных просветов и погруженных в жировую подушку. В таком состоянии млечные железы остаются до следующего сезона размножения. Эти связанные с лактацией структуры и процессы в целом соответствуют таковым у других млекопитающих.
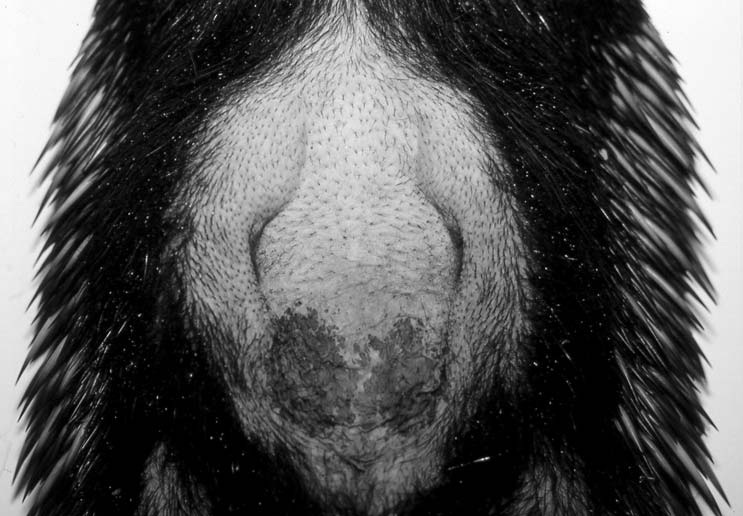
Выбритая брюшная поверхность самки австралийской ехидны (Tachyglossus aculeatus) с сумкой и млечными железами.
Молоко желтоватое, его состав меняется по мере роста детеныша. В момент появления на свет последнего молоко относительно жидкое и содержит приблизительно 12 % сухих веществ: 1,25 % жиров, 7,85 % белков, 2,85 % углеводов и минералов. Из числа последних уровень содержания железа составляет 8,3 мг/мл. Отношение олеиновой кислоты к пальмитиновой кислоте составляет около единицы (32,1 : 28,9). По мере созревания концентрация питательных веществ в молоке возрастает, оно становится густым и приобретает тягучую консистенцию. Спустя несколько месяцев молоко уже содержит 48,9 % сухих веществ: 31 % жиров, 12,4 % белков, 1,6 % гексозы, 0,7 % сиалиллактозы и 0,5 % минеральных веществ. Содержание железа в зрелом молоке достигает 43 мг/л — примерно вдвое больше, чем у утконоса. Также в нем выявлены натрий, калий, кальций, магний, цинк, медь.
Как и у других млекопитающих, молочные жиры включают небольшое количество свободных жирных кислот, сложные эфиры холестерина, фосфолипиды, а также до 95 % триглицеридов. Из триглицеридов зрелого молока ехидны основными жирными кислотами являются олеиновая (61,2 %), пальмитиновая (15,9 %), пальмитолеиновая (6 %), линолевая (5 %) и стеариновая (4 %). В отличие от раннего молока, отношение олеиновой кислоты к пальмитиновой кислоте составляет 3,8 : 1. Триглицериды зрелого молока содержат очень мало полиненасыщенных жирных кислот, в отличие от триглицеридов молока утконоса, вмещающего значительные их количества. Примечательно, что морфологически очень похожие молодая ехидна и утконос растут на столь различающемся молоке.
Количество белка в молоке возрастает вплоть до самого завершения лактации. Это может быть связано с необходимостью синтеза кератина для волос и иголок, обеспечивающих готовящимся к самостоятельной жизни молодым особям защиту от низких температур и хищников. Расшифровка генетических последовательностей белков показала как их сходство с таковыми териевых млекопитающих, так и наличие компонентов, специфических для однопроходных.
Основными углеводными компонентами молока являются фукозиллактоза и сиалиллактоза. Расщепление этих и ряда дополнительных олигосахаридов осуществляется не на границе кишечных ворсинок, как это характерно для плацентарных, а происходит внутриклеточно, с высвобождением пищеварительных ферментов из лизосом. Для транспортировки внутрь клеток молочных олисахаридов в апикальной цитоплазме клеток слизистой оболочки кишечника подсосного детеныша имеется специальная сеть канальцев.
Из сыворотки молока ехидны можно добыть лишь небольшое количество лактозы. В то же время лактоза является главным молочным сахаром плацентарных, у которых ее синтез катализируется 2 белками, включая альфа-лактальбумин. Последний в очень низкой концентрации выявлен и в молоке ехидны. Его аминокислотная последовательность существенно отличается от таковой у териев и указывает на родство с утконосом. Другой молочный белок — лизоцим — структурно подобен альфа-лактальбумину. Он имеется у ехидны, но, насколько известно, его не удалось выделить из молока утконоса. Ранее полагали, что лизоцим в определенной мере способствует образованию у ехидны лактозы. Однако оказалось, что даже при высокой концентрации этот белок на синтез лактозы никак не влияет. Таким образом, производство лактозной составляющей олигосахаридов молока ехидны происходит по той же схеме, что у утконоса и других млекопитающих.
Энергообмен
Ехидна преимущественно гомойотермна, но при этом температура ее тела относительно низкая и может значительно колебаться. При температуре воздуха 0-24 °С, средняя температура тела составляет 30,7 °С, с максимальным суточным отклонением 4,1 °С в ту или другую сторону. В выводковой сумке температура достигает 33-35 °С. Полевые наблюдения в Снежных горах показали, что при температуре среды 0 °С температура тела активной особи остается в пределах обычных 29-32 °С. В лаборатории при снижении температуры воздуха до 5 °С температура тела не опускается даже спустя 12 часов. В ответ на низкую температуру среды в несколько раз увеличивается потребление кислорода (т. е. возрастает уровень обмена веществ) и появляется мышечная дрожь, что предотвращает снижение температуры тела. Термонейтральная зона (температурный диапазон окружающей среды, при котором не происходит никаких изменений в потреблении кислорода) составляет 20-30 °С. Следовательно, при температуре среды ниже 20 °С метаболическое тепло, вырабатываемое для поддержания температуры тела, должно сохраняться путем сокращения его оттока в окружающее пространство.
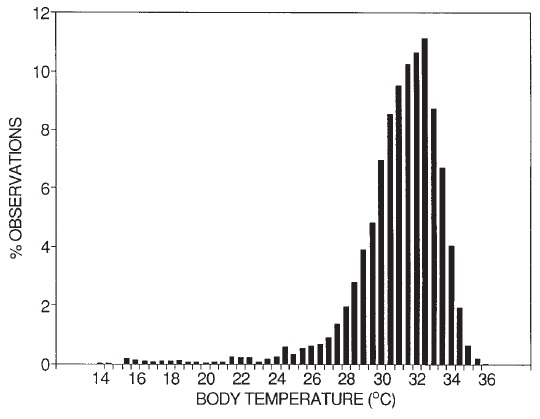
Частотное распределение всех показателей температуры тела австралийских ехидн (Tachyglossus aculeatus), измеренных посредством радиотелеметрии во время периода активности в Снежных горах.
Вырабатываемое организмом метаболическое тепло измеряется в калориях кв. см/час на °C. Согласно расчетам, при температуре окружающей среды 20 °C теплопроводимость тканей ехидны составляет 0,15 кал/кв. см/ч на °C. Это свидетельствует о том, что вазоконстрикция достигает максимума уже при 20 °C. Выше 20 °C наблюдается постепенное увеличение теплопроводимости, пока она не достигнет 0,7 кал/кв. см/час на °С при температуре среды 34 °С. Этот показатель для ехидны является гипертермическим и вызывает повышение ее собственной температуры примерно до 37 °C. При температуре тела свыше 38 °С животное обычно гибнет. Тем не менее, полевые наблюдения продемонстрировали, что хотя в некоторых районах Западной Австралии температура окружающей среды может достигать порядка 47 °C, температура тела обитающих там ехидн не поднимается выше 30 °C.
Отводить из организма избыточное тепло ехидне непросто, поскольку у нее отсутствуют потовые железы и не развит механизм тепловой одышки. Также она лишена возможности вылизывать себя влажным языком, как это делают многие другие млекопитающие. Установлено, что справляться с жарой ехидне помогает выдувание из ноздрей пузырьков слизи. Пузырьки лопаются и увлажняют кончик рыльца, содержащий множество кровеносных сосудов. Благодаря дальнейшему испарению влаги с рыльца кровь, а затем и весь организм животного охлаждается. Сделанные в инфракрасных лучах изображения ехидн, демонстрирующие картину распределения их телесной температуры, показываают: самыми теплыми обычно оказываются глазо-ушные области, а наиболее прохладным — кончик рыльца с ноздрями.

Пузырек слизи на кончике рыльца австралийской ехидны (Tachyglossus aculeatus).
Кроме того, для понижения или повышения внутренней температуры ехидна направляет кровь к коже и прочь от нее. Наиболее качественно терморегуляция реализуется через открытые области на брюхе и конечностях, которые можно просто прижать прохладной земле. Иглы, покрывающие тело животного сверху и с боков, обеспечивают ему дополнительную теплоизоляцию. Но поскольку вазомоторные реакции полностью проявляются уже при температуре среды 34 °С, вполне очевидно, что небольшое испарительное охлаждение в этом случае является явно недостаточным для предотвращения летального исхода. В воздухе с температурой 34 °С при испарении рассевается только 1/3 метаболического тепла. В связи с этим ехидна избегает активности в жаркую погоду. Это достигается путем смещения периода активности на более прохладное время суток. Остальное время животное проводит в естественных или специально вырытых подземных убежищах, температурный режим которых относительно стабилен. При наличии водоема ехидна может в нем плавать, что тоже помогает избавиться от излишков внутреннего тепла.7
В ходе эксперимента особи из жарких засушливых регионов продемонстрировали наличие преимущества в борьбе с высокой температурой окружающей среды по сравнению с их сородичами из районов с более умеренным климатом. Представители подвидов Tachyglossus aculeatus acanthion, Tachyglossus aculeatus aculeatus и Tachyglossus aculeatus setosus имеют соответственно короткий и редкий, средней длины и густоты, а также длинный и густой шерстный покров, обеспечивающий различную теплопроводимость. После 16 дней пребывания в засушливой среде температурой 20 °C теплопроводимость у Tachyglossus aculeatus acanthion составила 0,336 кал/кв. см/час на °C, у Tachyglossus aculeatus aculeatus — 0,254 кал/кв. см/час на °C, у Tachyglossus aculeatus setosus — 0,195 кал/кв. см/час на °C. Температура тела при этом составила 28,2 °C, 29,6 °C и 30,1 °C соответственно. Таким образом, высокая теплопроводимость тканей Tachyglossus aculeatus acanthion явно полезна при нахождении в жаркой и засушливой среде.

Изображение австралийской ехидны (Tachyglossus aculeatus), полученное с помощью инфракрасной термографической камеры. Наиболее прохладные участки тела отмечены голубовато-фиолетовым цветом, а самые теплые по мере возрастания — красным, оранжевым и желтым.
Обратное было продемонстрировано путем снижения температуры среды до 5 °C на протяжении следующих 42 дней. В итоге температура тела Tachyglossus aculeatus acanthion значительно понизилась и составила 23 °C, у Tachyglossus aculeatus aculeatus — 27 °C, у Tachyglossus aculeatus setosus — 28,5 °C. Благодаря невысокой теплопроводимости тканей сильно обволошенные особи способны поддерживать нормальную температуру тела даже при очень низкой температуре внешней среды. После того, как температура была постепенно повышена до 30 °С в течение ближайших 35 дней, представители всех 3 подвидов приобрели теплопроводимость 0,17 кал/кв. см/час на °С. На приведенные результаты оказали влияние такие неучтенные факторы, как толщина подкожной жировой клетчатки и условия содержания, в которых животные находились до начала проведения эксперимента.
Согласно результатам другого исследования, средняя температура тела особей, содержавшихся при 25 °C, стабильно составляла 32,2 °C, колеблясь не более чем на 2 °C в сутки. В то же время у особей, постоянно находившихся в среде с температурой 10 °C, регулярные колебания температуры тела достигали 10 °C на протяжении суток. По этой причине ехидна нередко рассматривается как гетеротермное животное. Относительно низкая температура тела наблюдается ранним утром, когда животное еще неактивно. Увеличение температуры тела на протяжении дня совпадает по времени с возрастанием активности и поисками корма. Таким образом, основным источником телесного тепла ехидны выступает сократительная деятельность ее поперечно-полосатой мускулатуры. Это подтверждается результатами инъекции флакседила (препарата, устраняющего мышечный тонус) на животных, содержавшихся при температуре 5 °C. Ответной реакцией организма явилось основательное падение температуры примерно до 22 °С за 3,5 часа. Вызванную флакседилом гипотермию можно легко преодолеть, поместив ехидну в среду с комнатной температурой.
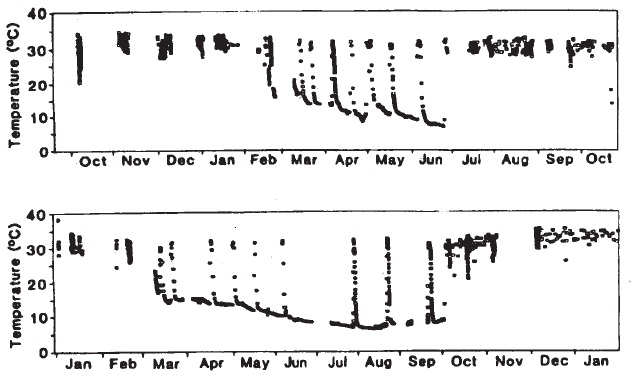
Показатели температуры тела 2 спящих австралийских ехидн (
Tachyglossus aculeatus), регистрируемые радиотелеметрией в Снежных горах (Новый Южный Уэльс). Сезон зимней спячки в обоих случаях отчетливо выражен.
Итак, когда ехидна не активна, она "отключает" терморегуляцию и благодаря этому экономит энергию. На отдыхе в тени температура тела опускается до 28 °C, а во время летнего оцепенения — до 18 °C. При зимнем понижении температуры среды до 5 °С и ниже, температура тела спящего животного опускается до 3-6 °С, оно становится жестким и практически неподвижным. Однако и в это время раз в 2-3 недели она быстро (за 12 часов) повышается до 30-32 °С и остается на этом уровне в течение 1-2 суток. В такие периоды ехидна нередко начинает "суетиться", перебегая с места на место, но потом снова цепенеет. По некоторым данным, минимальная зарегистрированная температура тела спящей ехидны составляет 0,5 °С.
Наряду с температурой тела, во время зимней спячки значительно понижаются и другие жизненные показатели. Частота сердечных сокращений падает до 4-7 ударов в минуту (по сравнению с 50-70 ударами у активного животного), а число дыхательных движений уменьшается до 1 в 3 минуты (средняя частота во время активности — 6 вдохов в минуту). Могут наблюдаться периоды апноэ продолжительностью до 2 ч. Расход кислорода в состоянии спячки составляет 0,02-0,03 мл/г/час (в активном состоянии — порядка 0,5 мл/г/час), выделение азота — 40-50 мг/кг/сут. Потеря веса в среднем достигает 2,07 г/кг/сут. В целом интенсивность обмена веществ может сократиться до 1/8 от летней нормы. Наоборот, во время коротких зимних пробуждений она возрастает в 9 раз по сравнению с летним покоем.
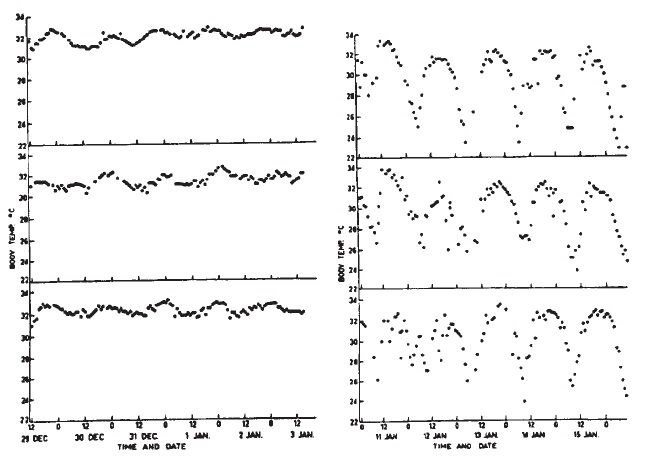
Показатели температуры тела 3 австралийских ехидн (Tachyglossus aculeatus), измеренные радиотелеметрией в ходе лабораторных исследований. Животные содержались при постоянной температуре 25 °C (слева), а затем 10 °C (справа). Изменения телесной температуры коррелируют с паттернами активности. Температура тела начинает снижаться после того, как животные становятся неактивными и растет по после того, как они начинают проявлять активность.
Состояние оцепенения может быть вызвано искусственно в любое время года путем лишения животного пищи и содержания его при температуре среды 5 °C. Мелкие особи становятся неактивными в течение 3-9 дней после принятия этих мер. Для более крупных требуется больше времени, свыше 32 дней. Если находящуюся при 5 °C неактивную особь слегка потревожить, можно вызвать постепенное повышение температуры ее тела от 6 °C до нормальной, которая при окружающей температуре 5-6 °C будет сохраняться на протяжении 20-часового периода пробуждения. Способность пробуждать от оцепенения утрачивается после многократной длительной выдержке при температуре среды 5 °C, но пробуждения всегда можно добиться, повысив эту температуру. Если она резко повышается с 5 °C до 25 °C, температура тела может увеличиться до 28 °C за 11 часов. Как уже было упомянуто, способность входить в оцепенение связана с деятельностью надпочечников.
Ехидны, встречающиеся на острове Кенгуру и принадлежащие к подвиду Tachyglossus aculeatus multiaculeatus, тоже впадают в зимнее оцепенение. Благодаря использованию радиослежения стало возможным отыскивать их и фиксировать физиологические показатели. Одно такое животное, весившее 2,6 кг, при повторной поимке имело температуру тела 13,6 °C. После помещения его в теплые условия (18-20 °C) в течение 22 часов после начала пробуждения температура тела менялась следующим образом: 13,5, 16,0, 17,5, 24,0 и 26,0 °С. При последнем замере температуры животное уже являлось достаточно оживленным. На острове Кенгуру также наблюдали, как находившиеся во вялом состоянии ехидны принимают солнечные ванны.
Согласно расчетам, средний уровень обмена веществ у ехидны равен 2,3 Вт/кг. Это составляет всего около 30 % от интенсивности метаболизма у большинства плацентарных и 60 % у сумчатых, что делает ехидну наименее "энергозатратным" млекопитающим. Указанная цифра близка к соответствующим показателям других мирмекофагов. Для роющих животных обычно также характерен пониженный обмен веществ. Это подтверждают результаты экспериментов с использованием беговой дорожки, согласно которым ехидна во время передвижения затрачивает на единицу веса энергии меньше, чем плацентарные млекопитающие. При этом потребление организмом кислорода возрастает линейно по отношению к скорости передвижения. Полевые данные показывают, что как кормящие, так и нелактирующие самки имеют примерно равные показатели интенсивности метаболизма и потребления кислорода.
Продолжительность жизни
Максимальная зарегистрированная продолжительность жизни ехидны составляет свыше 56 лет, что примерно в 4 раза больше, чем можно было бы ожидать от млекопитающего ее величины.
Известно, что долгоживущим млекопитающим свойственны устойчивые к окислению клеточные мембраны, регулирующие содержание полиненасыщенных и мононенасыщенных липидов. В мембранах ехидны количество полиненасыщенных липидов ниже, а мононенасыщенных выше ожидаемой. Такое их соотношение указывает на особенно высокую устойчивость клеточных мембран к окислению. Продолжительность жизни также связана с образованием свободных радикалов, которая пропорциональна интенсивности метаболизма. У ехидны значительно понижена скорость обмена веществ, за исключением периодов пробуждения от зимнего оцепенения. Во время этих эпизодов активности метаболизм значительно увеличивается, а производство свободных радикалов достигает своего максимума.
В связи с этим считается, что ехидна обладает хорошо развитыми уравновешивающими механизмами, способствующими высокой продолжительности ее жизни. По-видимому, в этот процесс вовлечен крупный и сложно структурированный мозг. Подобные его характеристики у других млекопитающих зачастую сопутствуют таким чертам жизнедеятельности, как медленное созревание и единственный детеныш в помете. Эти признаки, в свою очередь, коррелируют с высокой продолжительностью жизни.
Генетика
Согласно первоначальной оценке, геном ехидны меньше чем у утконоса. В кариотипе имеется 63 (самец) или 64 (самка) хромосомы. Подобно другим однопроходным, наличествуют множественные половые хромосомы. Они объединены в комплекс и в мейозе ведут себя как единое целое. У самца образуются сперматозоиды, имеющие цепочки X1X2X3X4X5 и Y1Y2Y3Y4. Когда сперматозоид X1X2X3X4X5 оплодотворяет яйцеклетку X1X2X3X4X5, рождается детеныш женского пола, если сперматозоид Y1Y2Y3Y4 — мужского. Таким образом, комбинация половых хромосом X1X1X2X2X3X3X4X4X5X5 дает самку, а комбинация X1Y1X2Y2X3Y3X4Y4X5 — самца.
Исследования выявили у ехидны и утконоса гомологию половых хромосом X1Y1, X2Y2, X3Y3 и X4Y5, однако у ехидны Y4 и Х5 гомологичны аутосомам утконоса I2 и Y4. Кроме того, порядок расположения половых хромосом в мейотической цепи ехидны отличается от таковой утконоса. Примечательно, что эти половые хромосомы не гомологичны таковым териевых млекопитающих, но гомологичны Z-хромосоме птиц. Это объясняет отсутствие у однопроходных гена SRY, определяющего пол у териев. Как и у утконоса, пол определяется геном антимюллерова гормона (АМГ) на самой старой Y-хромосоме.