Состав
Систематика рода эласмотериев строится на тонких различиях в строении черепа, нижней челюсти, зубов и костей посткраниального скелета. В объем рода включено 5 видов.

Прорисовка типового черепа Elasmotherium primigenium, вписанного в телесные очертания, и реконструкция его внешнего облика. Автор — Чен Ю.
Elasmotherium primigenium Sun, Deng et Jiangzuo, 2021 — вид, известный из позднего миоцена Китая (г. Янцзинь в уезде Динбянь провинции Шаньси).
Таксон установлен по черепу с отломленной передней частью. Из его зубов хорошо сохранился только правый М2; М3 и левый М2 сильно повреждены. Судя по сросшимся швам, образец принадлежит старой особи.

Поврежденный череп Elasmotherium primigenium из провинции Шаньси, Китай: a — сбоку; b — сверху; c — снизу; d — сзади.
Размеры крупные: длина неполного черепа превышает 52 см. Как и у более поздних форм, имеется хорошо выраженное лобное возвышение длиной 24 см, шириной 18 см и высотой 16 см. Возвышение выступает за край глазницы, его передний край находится на уровне М3. Теменные гребни широком разделены. Затылочная поверхность широкая, ориентирована вертикально.
Щечные зубы гипсодонтные, с хорошо развитым цементом и складчатой эмалью. Протокон и гипокон сильно сужены; задняя долина закрытая; криста хорошо развита и раздваивается; кроше отсутствует; складки паракона и метакона отсутствуют; эктолоф прямой; губной и язычный цингулюмы отсутствуют.
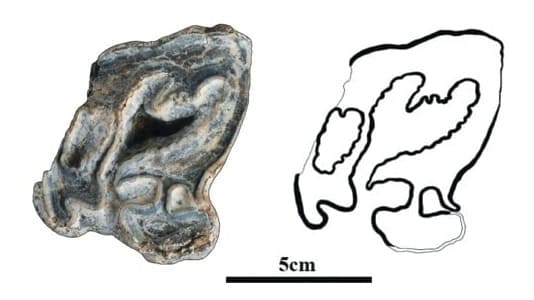
Жевательная поверхность М2 Elasmotherium primigenium из провинции Шаньси, Китай, и ее прорисовка
По своим морфологическим характеристикам данный вид занимает промежуточное положение между филогенетически более архаичным родом Sinotherium и более молодыми представителями рода Elasmotherium. Обращает на себя внимание более прямой угол затылка и менее выраженная складчатость зубной эмали, чем у последних.
Судя по всему, этот носорог обитал на открытых пространствах Восточной Азии, где в то время доминировали трехпалые формы лошадей, такие как Hipparion.
Хапровский эласмотерий (Elasmotherium chaprovicum Shvyreva, 2004) — вид, известный из раннего плейстоцена (гелазий, первая половина среднего акчагыла, средний виллафранк, MN17) Молдавии (с. Новые Танатары, с. Фарланданы) и России (Ростовская область: хут. Хапры, хут. Ливенцовка, хут. Мокрый Чалтырь; Ставропольский край: с. Саблинское). Синонимы: Elasmotherium caucasicum Borissiak, 1914 (частью); Elasmotherium sp. Shvyreva, 1995.
Таксон установлен по правому слабостертому М2 из хапровской песчаной толщи Ливенцовского карьера в северо-восточном Приазовье. Обнаружен фрагментарный материал: обломок черепа, зуб М2, фрагмент нижней челюсти с зубами m2 и m3, лучевая кость, крючковатая кость запястья из Ливенцовского карьера; зубы Р4 и М1 из карьера Мокрый Чалтырь; IV-я пястная кость из карьера Зубов сад в районе с. Саблинского; астрагал из района с. Новые Танатары. Поскольку найденного материала пока слишком мало, дифференцировать данный таксон по морфологическим признакам в настоящее время сложно.
Череп крупный, широкий в скуловых дугах.
Эмаль по контуру зубов простая и грубая. Складчатость эмали верхних щечных зубов хорошо проявляется на стенках, окаймляющих переднюю и заднюю долинки. Криста и антекроше развиты сильно на коренных, на предкоренных отсутствует криста. На сильно стертых коренных зубах развивается кроше. Возможно замыкание передней долинки за счет слияния кроше и антекроше. Эмаль, окаймляющая долинки, сильно складчатая. Щечные зубы демонстрируют тенденцию к замыканию корней. По соотношению ширины и длины нижние щечные зубы отличаются от зубов Elasmotherium peii, Elasmotherium caucasicum и Elasmotherium sibiricum.
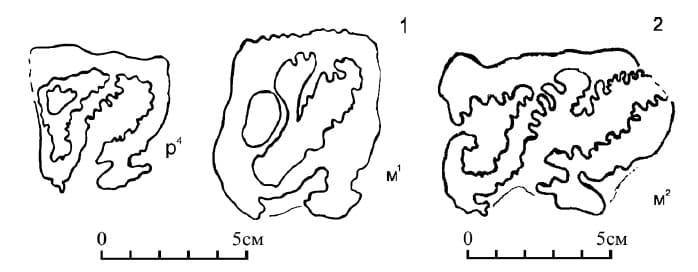
Прорисовки жевательных поверхностей зубов хапровского эласмотерия (Elasmotherium chaprovicum): 1 — Р4 и М1 из карьера Мокрый Чалтырь, Россия; 2 — М2 (голотип) из Ливенцовского карьера, Россия.
Кости кисти массивнее, чем у Elasmotherium caucasicum и Elasmotherium sibiricum. Блок астрагала (надпяточной кости) сильно сдвинут наружу. Нижний медиальный бугор дистальной части кости выражен резко. Шейка тела кости обозначена хорошо. Ямка под блоком четко отделяет его от кромки нижней суставной поверхности для костей заплюсны. На ее дне имеются сосудистые отверстия. Нижняя суставная фасетка на задней поверхности для сочленения с пяточной костью вогнутая, в то время как у Elasmotherium sibiricum она плоская.
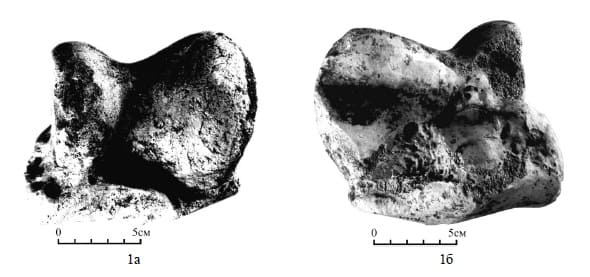
Астрагал хапровского эласмотериея (Elasmotherium chaprovicum) из села Новые Танатары, Молдавия: а — спереди, б — сзади.
Животное входит в состав хапровского фаунистического комплекса Восточной Европы.

Эласмотерий Пэйя (Elasmotherium peii) в естественном окружении. Автор — Р. Евсеев.
Эласмотерий Пэйя (Elasmotherium peii Chow, 1958) — вид, известный из раннего плейстоцена Китая (провинция Шаньси: Сихоуду, Санмень; провинция Хэбэй: Сяшагоу, Шаньшэньмиаозуй и Хэйтугоу в Нихэваньской котловине), а также раннего плейстоцена (поздний акчагыл - первая половина раннего апшерона) Молдавии (село Салчия), Украины (Одесская область: Жевахова гора, Куяльницкий лиман, с. Долинское; Николаевская область: урочище Луговая Балка у города Николаев; Запорожская область: город Токмак) и России (Ставропольский край: станица Расшеватская, станица Кармалиновская, город Новоалександровск; Краснодарский край: река Псекупс). Синонимы: Elasmotherium caucasicum Borissiak, 1914 (частью); Elasmotherium sp. Teilhard de Chardin et Piveteau, 1930; Elasmotherium inexpectatum Chow, 1958; Elasmotherium cf. inexpectatum Chia et Wang, 1958.
Таксон установлен по правому ряду зубов верхней челюсти Р3-М3 и левым DР4, Р3, М2, М3, принадлежавшим одной особи из отложений позднего плиоцена / раннего плейстоцена района Санменьского ущелья в районе деревни Пингло на юге провинции Шаньси. Среди обнаруженного материала: сильно поврежденный череп из г. Токмака; зуб М2 из окрестностей города Одессы; 8 зубов с Жеваховой горы и Куяльницкого лимана; 4 верхних зуба (Р4 и 3 М2 разной степени стертости) из ст. Расшеватской; 2 нижних зуба из Ставрополья; 7 зубов из д. Пингло; III-я пястная кость из ст. Бакинской; левая локтевая кость, левая головчатая кость, правые III-я и IV-я пястные кости, правая большая берцовая кость с частью малой берцовой кости, левая таранная кость, правая кубовидная кость, часть левой IV-й плюсневой кости, дистальная фаланга III-го пальца стопы из Шаньшэньмиаозуй.

Поврежденный череп эласмотерия Пэйя (Elasmotherium peii) из раннего плейстоцена окрестностей города Токмак, Запорожская область, Украина: А — сбоку; Б — сверху; В — снизу; Г — сзади. Национальный научно-природоведческий музей НАН Украины, Киев. Образец сохранился почти полностью, кроме незначительной части переднего отдела и передней части куполообразного возвышения. Значительная стертость зубов и полное срастание швов между костями черепа свидетельствует о принадлежности его взрослому животному. Первоначально этот череп был описан как принадлежащий Elasmotherium caucasicum. Однако по сравнению с последним он обладает более мелкими зубами с более толстой эмалью, простой по периметру. Складки эмали, окаймляющие долинки, крупные и расположены редко. При малой стертости (длина М2 превышает его ширину) щечные зубы имеют замкнутые корни. Кроме того, соотношение длины ряда М1-3 и длины полного зубного ряда (индекс 81,85) отличается от Elasmotherium caucasicum (индекс 85,43) и совпадает с Elasmotherium peii из Китая (индекс 81,92). На основании проведенного сравнения череп из Токмака отнесли к виду Elasmotherium peii.
Размеры крупные, сравнимые с Elasmotherium sibiricum, но уступающие Elasmotherium caucasicum. Длина черепа из Токмака от затылочного мыщелка до переднего края альвеолы Р3 составляет 82 см. Череп крупный и вытянутый. Лицевая часть его незначительно опущена относительно мозговой: касательная к альвеолярному краю челюсти, продолженная назад, проходит несколько ниже мыщелков. Соответственно низко расположена и суставная поверхность для нижней челюсти. Задний отдел черепа удлинен, его длина от задней поверхности затылочных мыщелков до переднего края засуставного отростка составляет около 1/3 расстояния от задней поверхности затылочных мыщелков до начала зубного ряда (Р3). Верхняя линия профиля сильно вогнута в задней части, вблизи затылочного гребня сильно выпукла.
Верхнечелюстные, предчелюстные и носовые кости прочно сросшиеся. Очень иростые носовые кости суживаются к переднему концу клином, слегка наклонены вниз, в боковом направлении выпуклые, края закругленные. Костная перегородка своим задним концом входит в подкупольную часть. Лобное вздутие у черепа из Токмака длиной 38 см, шириной 26,5 см и высотой 17,3 см. Его верхняя поверхность не несет ругозистости. Однако шероховатость хорошо видна на боковых и задней стенке купола, усиливаясь к его основанию. Крупная глазница хорошо очерчена. Она расположена почти на середине высоты черепа. Ее передний край находится на уровне заднего края альвеолы М3. Передние стороны глазниц ограничены большими мрщными выростами, как бы прикрываюзими их спереди. Скуловая дуга относительно короткая и толстая, особенно в переднем и заднем краях, в средней части слабо изогнута и уплощена, лежит низко (чуть выше альвеолярного края). Суставная поверхность для нижней челюсти выступает внутрь за уровень засуставного отростка. Засуставной отросток короткий, массивный, спереди уплощен и несет фасетку для бугра нижней челюсти. Яремный отросток выражен слабо, далеко отстоит от затылочных мыщелков. Височная кость имеет удлиненную, вогнутую сильно и шероховатую чешую, что указывает на крепление здесь сильного мускула.
Теменные кости сильно вогнуты в передне-заднем направлении, имеют седловидную форму, резко поднимаясь к затылочному гребню. Боковые теменные гребни широко расставлены, идут почти параллельно друг другу, однако вблизи затылочного гребня они расходятся в стороны. Затылок назад не отклонен. Затылочная кость низкая, широкая. На черепе из Токмака угол, образуемый вертикальной остью большого затылочного отверстия и небом, равен 105 °, а угол затылочного гребня — 57 °. Затылочные бугры развиты сильно, но не нависают над затылочным отверстием. На основной затылочной кости (по средней линии) имеется сильно вогнутая впадина, разделяющая кость на 2 части, верхняя из которых заканчивается 2 мощными буграми. Гребни затылочной области очень сильно развиты. Посредине гребня затылочной кости имеется значительная впадина треугольной формы. Выйная впадина небольшая, посередине нее имеется невысокий гребень. Сосцевидные части сильно развиты в стороны и выступают за уровень скуловой дуги и глазницы. Большое затылочное отверстие субккруглое (высота несколько больше ширины). Засуставный отросток отходит от внутреннего края суставной впадины, прилегая изнутри вплотную своей уплощенной поверхностью к выросту — бугру нижней челюсти, и препятствует смещению нижней челюсти внутрь.
Суставная поверхность в месте сочленения с нижней челюстью немного выпуклая посередине, сильно выпуклая в ширину (индекс передне-заднего поперечника составляет 40 %). Затылочные мыщелки крупные, овальные, сильно вытянутые (индекс их поперечников порядка 73 %) с выпрямленным передненижним краем, в передненижней части очень выпуклые и уплощенные, относительно вертикальной оси затылочного отверстия поставлены косо вверх и наружу, от затылочной кости со всех сторон они отделены желобком. Основная часть затылочной кости короткая, широкая, утолщенная, в передней части несет крупные овальные глоточные бугры. Базисфеноид имеет форму удлиненного конуса. Твердое небо узкое, с вогнутой поверхностью. На небном отростке предчелюстной кости имеется 2 пары небных щелей. Небная вырезка открывается на уровне переднего края альвеолы М3. Строение других элементов черепа из-за значительных повреждений описать трудно.

Щечные зубы эласмотерия Пэйя (Elasmotherium peii) из станицы Расшеватская, Ставропольский край, Россия:1 — m2 (а — сверху, б — губная поверхность, в — языковая поверхность); 2 — m3 (а — сверху, б — языковая поверхность, в — прикорневая часть зуба).
Ряды предкоренных и коренных образуют дугу с резко выраженным изломом в области Р4. Имеется 3 верхних предкоренных зуба. Ряд предкоренных вдвое короче ряда коренных зубов. Верхние сильностертые коренные зубы имеют хорошо отличимые корень и коронку. Задняя долинка длительно существует открытой. Корни замыкаются до ее закрытия. На задней стенке всех зубов на ранних стадиях стирания есть эмалевый воротничок в виде тонкой полоски. Парастильная складка выражена нечетко. На сильно стертых зубах гипокон выступает внутрь (лингвально) за уровень протокона. Зубы обладают четкостью строения элементов жевательной поверхности. Поперечные гребни протолоф и металоф узкие, их лингвальные концы сильно отогнуты назад. На ранних стадиях стирания синусы, выделяющие гипокон, отсутствуют. Складчатость эмали развита в области долинок. Заметалофная часть эктолофа М3 длинная. Нижние коренные зубы широкие (индекс ширины 59,65), с толстой слабо извитой эмалью внутренних стенок полулуний. Эмаль наружных стенок простая, нескладчатая. Характерно образование переднего и заднего корней.

Прорисовки жевательных поверхностей зубов эласмотерия Пэйя (Elasmotherium peii): 1, 1а,1б — М2 и 5 — Р 4 из станицы Расшеватская Ставропольского края, Россия; 2 и 3 — М 2 и 4 — Р4 из провинции Шаньси, Китай; 6 — Р4 из окрестностей города Одесса, Украина.
М2 Elasmotherium peii отличаются от такового Elasmotherium chaprovicum из Ливенцовки по соотношению ширины и длины, более изящным строением элементов жевательной поверхности, более узкими лофами и более коротким протоконом по отношению к длине М2 (у Elasmotherium peii индекс 26,5; у Elasmotherium chaprovicum — 28,4). В отличие от Elasmotherium caucasicum, М2 Elasmotherium peii более мелкий и более узкий. Также этот зуб отличается наличием воротничка на задней стенке зуба. Складки эмали, окаймляющие переднюю и заднюю долинки, глубокие. Расположены они более упорядочено. Задняя долинка более длительно остается открытой. На слабо и умеренно изношенных зубах М2 и М3 заметалофная часть эктолофа длиннее чем у Elasmotherium caucasicum.
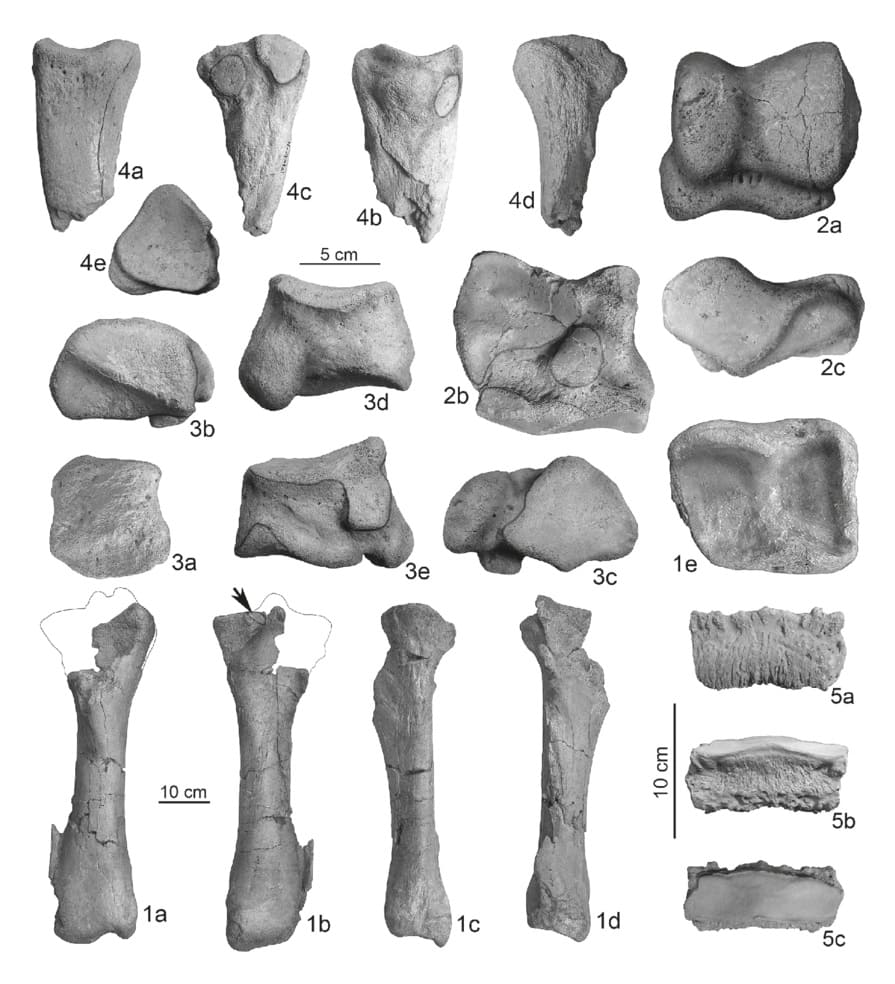
Кости задней конечности эласмотерия Пэйя (Elasmotherium peii) из Шаньшэньмиаозуй, провинция Хэбэй, Китай: 1 — правая большая берцовая кость с частью малой берцовой кости; 2 — левая таранная кость; 3 — правая кубовидная кость; 4 — часть левой IV-й плюсневой кости; 5 — дистальная фаланга III-го пальца стопы.
По сравнению с Elasmotherium sibiricum верхние коренные зубы Elasmotherium peii более узкие. Кроме того, они отличаются простой или слегка волнистой эмалью на периферических стенках лофов; наличием открытой или закрытой задней долинки даже на очень стертых зубах; обозначением на поздних стадиях стирания коронки и корня; ранним замыканием корней. Нижние коренные зубы Elasmotherium peii ýже, чем у Elasmotherium chaprovicum, но шире чем у Elasmotherium сaucasicum и Elasmotherium sibiricum. По сравнению с Elasmotherium sibiricum зубы имеют более грубую, простую эмаль на наружных стенках полулуний. Эмаль металофида и гиполофида на стенках, окаймляющих переднюю и заднюю внутренние долинки, более толстая, чем у Elasmotherium sibiricum, складки ее располагаются более упорядочено.
Животное входит в состав псекупского фаунистического комплекса Восточной Европы и нихэваньской фауны Северного Китая.

Кавказский эласмотерий (Elasmotherium caucasicum) в естественном окружении. Автор — Р. Евсеев.
Кавказский эласмотерий (Elasmotherium caucasicum Borissiak, 1914) — вид, известный из конца раннего плейстоцена (вторая половина раннего - поздний апшерон) Украины (Одесская область: село Крыжановка; Херсонская область: город Геническ; Запорожская область: город Бердянск, Приморск и Мариуполь), России (Краснодарский край: овраг Синяя Балка у станицы Ахтанизовской, Цимбал у поселка Сенной, станица Фонталовская; Ростовская область: село Самарское). Синоним: Elasmotherium sibiricum Fischer von Waldheim, 1808 (частью).
В качестве лектотипа выбраны фрагменты верхних челюстей с правым и левым зубными рядами Р4-М3 из отложений среднего и позднего апшерона оврага Синяя Балка в 2-3 км к северу от станицы Ахтанизовской на Таманском полуострове. Среди обнаруженного материала: неполный череп, фрагмент правой верхней челюсти с рядом Р4-М3, несколько нижних челюстей, более 10 нижнечелюстных ветвей, большая серия верхних и нижних зубов, позвонки, кости кисти, 4 астрагала из Синей Балки; нижняя челюсть и фрагмент лучевой кости из города Бердянска; VI-й шейный позвонок из города Мариуполя; фрагмент лопатки из Северного Причерноморья; левая плечевая кость из села Самарское. Полные скелеты пока не обнаружены.



Реконструированный скелет кавказского эласмотерия (Elasmotherium caucasicum). Азовский историко-археологический и палеонтологический музей-заповедник.
Самый крупный представитель рода Elasmotherium. Скелет, реконструированнный по остаткам разных особей в Азовском историко-археологическом и палеонтологическом музее-заповеднике, имеет 4,2 м в длину и 2,4 м в холке. Однако, принимая во внимание, что размеры некоторых изолированных щечных зубов, хранящихся в коллекциях, значительно превосходят размеры зубов, известных по верхним и нижним челюстям данного вида, можно полагать, что длина тела достигала, по меньшей мере, 5-5,2 м. Поврежденный череп очень крупной особи из Синей Балки имеет в длину более 1 м, а его полная длина оценивается примерно в 1,2 м. Это дает рост около 3 м в холке. Вес животного оценивается в 4-5,5 т и более.
Горизонтальная ветвь нижней челюсти низкая и длинная (около 73 см). Симфиз лопатообразный, короткий (39,53 % длины зубного ряда) и лопатообразно расширенный (индекс ширины 116,61), уплощенный и гладкий сверху, расположен почти горизонтально. Задний край его находится на уровне р3. На нижне-передней поверхности симфиза имеются 3 пары сосудистых отверстий. Подбородочное отверстие у молодого животного смещено вниз и лежит на уровне переднего края р4, а у взрослого — под задней половиной р3. Диастема короткая, ее длина от переднего края симфиза до альвеолы р2 составляет около 9 см (31,41 % длины зубного ряда). Альвеолярный край челюсти горизонтален. Вентральный край плавно поднимается к симфизу. Восходящая ветвь широкая.
Нижняя челюсть Elasmotherium саucasicum ниже, чем у Elasmotherium sibiricum. Горизонтальная ветвь менее быстро суживается к переднему концу, резкий подъем начинается на уровне p4. Нижний край ее и симфизный отдел расположены более горизонтально, образуя более пологую дугу. Симфизный отдел более широкий, уплощенный и короткий по сравнению с Elasmotherium sibiricum. Диастема короче, чем у Elasmotherium sibiricum.

Зубы верхней челюсти и коренной зуб кавказского эласмотерия (Elasmotherium caucasicum) из Приазовья, Россия. Палеонтологический музей им. Ю.А. Орлова РАН, Москва.
В каждой половине обеих челюстей полувзрослых и взрослых особей имеется по 3 предкоренных и 3 коренных зуба. Верхние коренные зубы призмовидные, коронка от корневой части почти неотличима, размеры крупные. Складчатость эмали развита только на внутренних стенках лофов, окаймляющих переднюю долинку. Замыкание задней долинки совпадает с началом образования корней. На сильно стертых зубах задняя долинка исчезает, развивается слабый кроше, возможно замыкание передней долинки в результате слияния протокона и кроше. Корни таких зубов замкнуты. Нижние зубы крупные. Складчатость интенсивнее развита на внутренних стенках полулуний. Наружная стенка металофида уплощенная, у гиполофида — выпуклая. Эмаль здесь простая и толстая. На сильно стертых зубах прослеживается замыкание задней долинки за счет сближения и слияния металофида и гиполофида.

Неполная нижняя челюсть кавказского эласмотерия (Elasmotherium caucasicum) из города Бердянск, Запорожская область, Украина. Национальный научно-природоведческий музей НАН Украины, Киев.
Главное отличие зубной системы Elasmotherium саucasicum от таковой Elasmotherium sibiricum — наличие 3 предкоренных в верхней и нижней челюстях. Верхние щечные зубы отличаются более слабой складчатостью эмали, меньшим изгибом по высоте, более длительным существованием задней долинки, замыканием корней на поздних стадиях стирания. М2 Elasmotherium caucasicum сходны с зубами Elasmotherium рeii и Elasmotherium chaprovicum в развитии складчатости, оттянутости вперед паракона и отличаются от них более крупными размерами, отсутствием воротничка на задней поверхности верхнекоренных зубов, более поздним замыканием корней, более сложной складчатостью эмали. Они более широкие, чем у Elasmotherium peii и Elasmotherium chaprovicum. Нижние щечные зубы его более узкие, чем у Elasmotherium реii и Elasmotherium chaprovicum, но шире, чем у Elasmotherium sibiricum. Зубам Elasmotherium caucasicum присущи такие признаки архаичности как развитие кроше в зрелом и пожилом возрасте, замыкание передней долинки на верхних и задней долинки на нижних зубах, чего никогда не встречается у Elasmotherium sibiricum. Наличие кроше и замкнутых долинок, по всей видимости, следует считать возрастным признаком.

Прорисовки жевательных поверхностей зубов кавказского эласмотерия (Elasmotherium caucasicum) из оврага Синяя Балка, Краснодарский край, Россия.
Полный посткраниальный скелет Elasmotherium caucasicum не описан. Телосложение несколько более тяжелое, чем у типового вида.
VI-й шейный позвонок имеет носорожий тип строения. Тело позвонка выпукло-вогнутое, сжато с боков. Головка овально-выпуклая, ямка сильно вогнутая. Основания остистого, нижних суставных и собственно реберных отростковх толстые и достаточно длинные. Отверстие позвоночного канала низкое, треугольно-овальное. Поперечные отверстия овальные, крупные. Позвоночная арка массивная, несколько наклонена вперед. Верхние суставные отростки крупные. Передние расставлены шире задних.
Лопатка крупная и широкая. Ость проходит почти посередине кости. Она простирается вдоль лопатки вентрально, сходит на нет, не достигнув суставной впадины. Поверхность заостной ямки сильно вогнута, а предостной — более уплощенная, кость здесь более тонкая. На медиальной поверхности кости развита глубокая подлопаточная ямка, наибольшая глубина ее лежит против вентального конца ости. Заямочная часть ее по сравнению с предъямочной более выпуклая и широкая. Лопаточный бугор массивный, шероховатый, округлый. Коракоидный отросток не развит. Шейка кости широкая и короткая. Суставная впадина неправильно-овальная, продолговатая, вогнута в обоих направлениях, но в передне-заднем — больше. На латеральной поверхности лопатки между суставной впадиной и лопаточным бугром находится шероховатое возвышение. Сильное развитие лопаточного бугра, шероховатостей, предостной, заостной и подлопаточной ямок свидетельствует о значительном развитии мускулатуры лопаточно-плечевого сустава и лопаточно-туловищных мышц. Они приводят лопатку к туловищу, поднимают и тянут ее вперед и назад.

Часть лопатки кавказского эласмотерия (Elasmotherium caucasicum) из Северного Причерноморья, Украина. Национальный научно-природоведческий музей НАН Украины, Киев.
Левая плечевая кость мощная, с укороченным диафизом треугольного сечения. Она несколько короче, чем у Elasmotherium sibiricum. Суставнаят головка немного шире, чем у последнего. Наименьшая ширина диафиза, ширина верхнего конца кости с дельтовидной бугристостью и нижнего конца с латеральным надмыщелком меньше, чем у Elasmotherium sibiricum. Диафиз плечевой кости Elasmotherium caucasicum за счет более мощного развития гребня плечевой кости толще, чем у Elasmotherium sibiricum (соотношение наименьшей ширины диафиза и поперечника там же у Elasmotherium caucasicum 116,9 %, а у Elasmotherium sibiricum — около 101,1 %).
Астрагал крупный, высокий и широкий, с узким блоком (индекс 97,87). Ширина суставного блока меньше ширины дистального отдела кости. Нижний медиальный бугор его развит сильно за счет смещения блока в латеральном направлении. Ниже суставного блока прослеживается шейка кости. Задняя поверхность кости несет 3 фасетки для сочленения с пяточной костью. Они не соприкасаются друг с другом. Астрагал относительно больше и стройнее, чем у других видов. Блок астрагала узкий по сравнению с Elasmotherium sibiricum и Elasmotherium chaprovicum. Относительная ширина его дистального отдела меньше, чем у Elasmotherium sibiricum и Elasmotherium chaprovicum.
Животное является одним из характернейших элементов таманского фаунистического комплекса Восточной Европы. Считается, что Elasmotherium caucasicum более примитивен, чем Elasmotherium sibiricum, и, вероятно, был для него предковым.
Изучение остатков Elasmotherium caucasicum позволяет высказать некоторые замечания об экологии этого вида. Известно, что у животных, питающихся мягкими кормами, нижняя челюсть испытывает меньшую нагрузку в вертикальном отношении и поэтому она низкая. Нижняя челюсть Elasmotherium caucasicum ниже, чем у Elasmotherium sibiricum. Это дает возможность предположить, что пища Elasmotherium caucasicum была не такой жесткой, как у Elasmotherium sibiricum. В пользу этого говорит и более простое строение эмали его щечных зубов. Кроме того, анализ сопутствующей ему фауны показывает широкое распространение лесных и лесостепных видов животных, что свидетельствует о хотя бы частичной залесенности территории Таманского полуострова в период обитания здесь Elasmotherium саucasicum. Распространение лесов сопровождается увлажнением климата, что способствует развитию растительности гумидного типа, которая и могла составлять пищу Elasmotherium caucasicum.