Строение и функции
Может показаться, что внешне гиенодоны несколько напоминают современных представителей семейства псовых, однако на самом деле они далеко не являются их аналогами. Данный род объединяет куда более разнообразных животных, занимавших разнообразные ниши сухопутных хищников. Однако вне зависимости от местообитаний, образа жизни и величины, всех гиенодонов роднит ряд общих, свойственных только им морфологических черт.

Сравнительные размеры некоторых видов гиенодонов, слева направо: Hyaenodon mustelinus, Hyaenodon horridus, Hyaenodon gigas, Hyaenodon macrocephalus, Hyaenodon leptorinchus. Автор — Р. Евсеев.
Род гиенодонов представлен довольно широким диапазоном размерных классов. Наиболее мелкие (например, Hyaenodon microdon и Hyaenodon mustelinus) достигали величины домашней кошки: длина их черепов составляла порядка 10-16 см, общая длина — 60-70 см, вес — 5-8 кг. С другой стороны, крупнейшие представители рода по размерам не уступали самому большому тигру: так, длина черепа Hyaenodon megaloides составляла 40 см, а у Hyaenodon gigas, судя по размерам его зубов и фрагментов челюстей, доходила до 60 см и даже более при расчетной общей длине в 350 см и массе в 380 кг. В среднем же большинство гиенодонов достигало роста волка или крупной собаки.

Реконструкция внешнего облика гиенодона (Hyaenodon brevirostrus). Автор — Р. Евсеев.
Череп
Череп гиенодонов длинный, узкий и довольно низкий, с вытянутым лицевым отделом и высоким, напоминающим акулий плавник сагиттальным гребнем. Скуловые дуги низко расположенные, прямые и тонкие. Костные слуховые пузыри иногда имеются. Нижняя челюсть также длинная и тонкая, с очень длинным симфизом. На первый взгляд, подобная черепная морфология особо ничем не отличается от таковой любого другого длинномордого млекопитающего хищника, однако при ближайшем рассмотрении она демонстрирует целый комплекс уникальных черт.
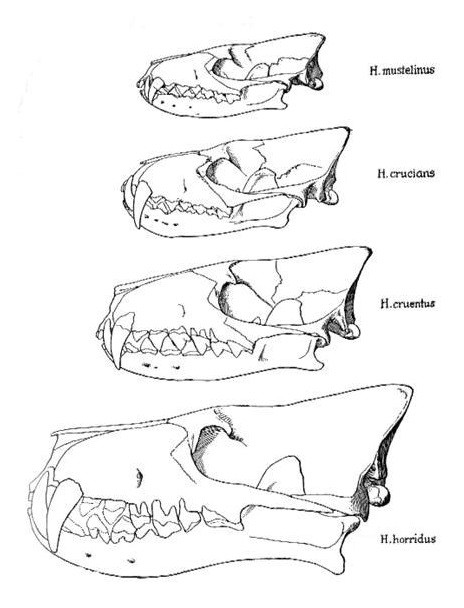
Прорисовки черепов с нижними челюстями различных видов гиенодонов.
Одними из первых на себя обращают внимание глазницы, расположенные далеко позади носовых костей, а спереди и сверху прикрытые костями, образующими полки. По сути, гиенодоны единственные известные наземные хищники, у которых глазницы находятся на боковых поверхностях черепа и ориентируются почти строго по сторонам — к примеру, у Hyaenodon crucians угол расположения глаз по отношению к сагиттальной оси черепа приближалось к 90 °. При таком положении глазниц поле зрения каждого глаза составляло почти 180 °, а оба глаза охватывали 320-340 °. В то же время, зона бинокулярного зрения была необычайно узкой и ограничивалась сектором в 10-30 °. Вероятно, чтобы посмотреть прямо перед собой и увидеть объемную картинку, гиенодонам приходилось поднимать нос вверх и сдвигать вперед и вниз свои крупные, подвижные и выпуклые глаза.






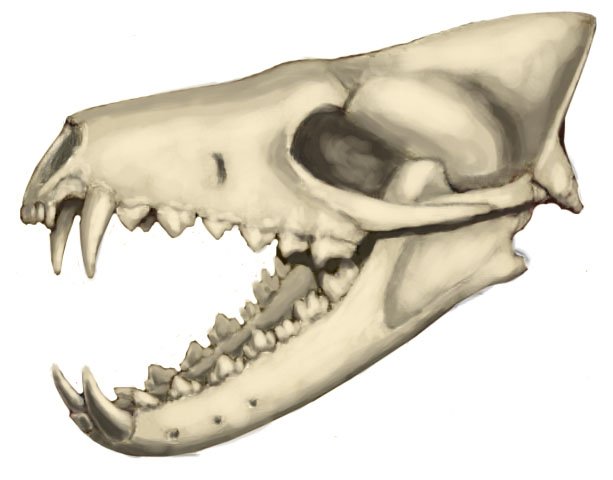
Череп с нижней челюстью гиенодона (Hyaenodon horridus).
Как правило, круговое зрение свойственно растительноядным млекопитающим, у которых такое положение глаз связано с необходимостью обозревать местность, высматривая хищников, но никак не специализированным засадным охотникам (коими и являлись гиенодоны), которым как раз нужен широкий сектор бинокулярного зрения для выслеживания жертвы и оценки расстояния до нее. Впрочем, это было собственным эволюционным приобретением гиенодонов (поскольку у их предков глаза были сдвинуты вперед, как у современных собак) и позволяло как-то по-своему скрадывать добычу.

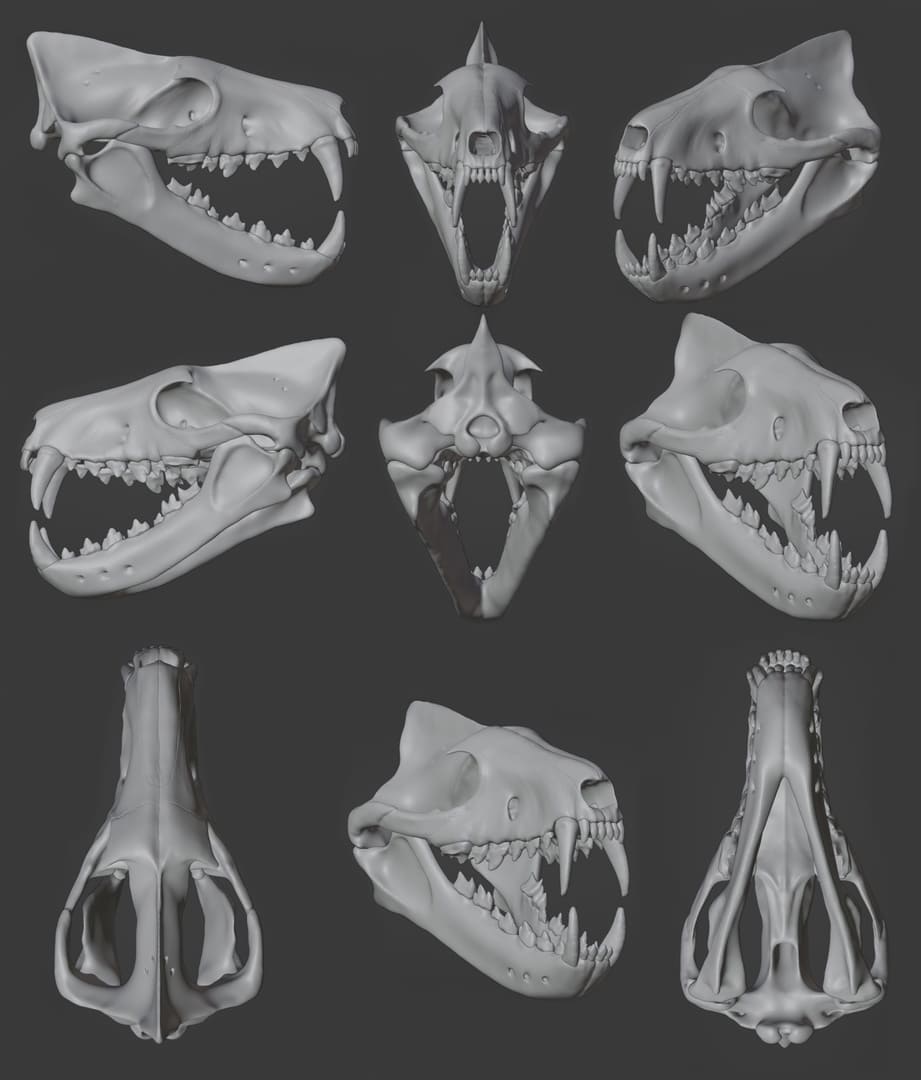
Компьютерные модели черепа с нижней челюстью гиенодона (Hyaenodon horridus). Автор — И. Белоусов.
Любопытно, что после смены зубов черепные кости гиенодонов еще долго продолжали расти, а швы черепа оставались неокостеневшими на протяжении всей жизни, в то время как у Carnivora рост черепа приостанавливается после смены зубов и черепные швы во многих случаях срастаются, препятствуя дальнейшему росту.


Этапы портретной реконструкции гиенодона (Hyaenodon horridus). Автор — Р. Евсеев.
Не менее интересной особенностью черепной морфологии гиенодонов является удлиненное небо. Их небные и крыловидные кости позади коренных зубов загибаются к средней линии черепа, где вполне или почти сходятся и образуют отведенные далеко назад костные носовые каналы. У современных Carnivora ноздри продолжаются за небом в трубке из соединительной ткани, и только у некоторых енотовых (Procyonidae) наблюдается подобное удлинение неба костными носовыми трубками, но оно значительно меньше, чем у гиенодонов. Исходя из этой особенности некоторые ранние авторы предполагали, что гиенодоны могут быть родственны енотам, барсукам, коати и мангустам из-за небольшого небного удлинения, наблюдаемого у этих животных. Также строились предположения о том, что гиенодоны вели водный образ жизни, поскольку подобное небное удлинение имеется у крокодилов и китообразных. Существовало даже мнение о специализации к питанию насекомыми, опять же, из-за наличия костных носовых трубок у муравьедов. Однако причины возникновения этой черты у представителей рода Hyaenodon кроются в ином — скорее всего, такие далеко отведенные назад костные носовые трубки позволяли этим хищникам глубоко захватывать в пасть части тела добычи (например, горло жертвы) или набивать пасть мясом, не боясь задохнуться.

Портретная реконструкция гиенодона (Hyaenodon horridus), демонстрирующая боковую ориентацию глаз. Автор — Р. Евсеев.
У ныне живущих хищных скуловые дуги преимущественно массивные и сильно выгнуты вверх. Подобное их строение напрямую связано с развитой массетерной мышцей, которая верхним концом крепится к скуловой дуге, чья выгнутая аркой кверху структура лучше выдерживает тянущие вниз нагрузки. У гиенодонов скуловые дуги низко опущены, тонкие, почти горизонтальные, и очень высокие сагиттальный и затылочный гребни. Это позволяет сделать вывод, что массетерные мышцы были редуцированы и компенсировались увеличенными крыловидными и височными мышцами. Об этом также свидетельствуют неглубокие массетерные ямки на нижней челюсти. В целом челюстная мускулатура была очень мощной и позволяла гиенодонам управляться даже с крупной добычей.
В отличие от современных Carnivora, у которых сочленение с нижней челюстью расположено почти вровень с уровнем мозговой коробки, суставная ямка гиенодонов находится заметно ниже уровня верхнего зубного ряда, спускаясь на своеобразной ножке. Примечательно, что такое низкое сочленение с нижней челюстью характерно и для махайродонтин (Machairodontinae), по-видимому, из-за сходных функциональных особенностей: саблезубым кошкам необходимо было широко раскрывать пасть, чтобы задействовать длинные клыки, у гиенодонов же это было связано, скорее всего, с необходимостью задействовать расположенные глубоко в пасти главные хищнические зубы, приспособленные для разрезания мяса. Установлено, что пасть этих хищников могла открываться под углом в 75 °.

Портретная реконструкция гиенодона (Hyaenodon horridus). Автор — Р. Евсеев.
Очередными довольно непривычными морфологическими особенностями гиенодонов являются сросшийся нижнечелюстной симфиз и сфероидальный суставный отросток (за исключением эоценовых форм). Эти черты свойственны растительноядным и всеядным млекопитающим, но не специализированным хищникам, у которых симфиз не сросшийся в связи с цилиндрической формой суставного отростка. У ранних, эоценовых гиенодонов имелся как раз цилиндрический мыщелок, обеспечивавший жесткое закрепление челюсти таким образом, что та могла двигаться только в вертикальной плоскости, и это просто необходимо для наилучшего использования режущих зубных рядов. Учитывая данное обстоятельство, вызывает недоумение смена цилиндрического мыщелка у более поздних видов на округлый, обеспечивающий ход челюсти еще и в горизонтальной плоскости. При всем этом, сфероидальный мыщелок суставного отростка создает трудности, отсутствующие у ранних форм с цилиндрическими мыщелками — усилия, прикладываемые вдоль проксимо-дистальной оси челюстей (например, при дроблении), рискуют вывихнуть челюсть. Быстрым решением этой проблемы явилось развитие большого складчатого симфиза и слияния на нем правой и левой ветвей нижней челюсти. Обе половины нижней челюсти в этом случае выступают как единое целое, и проворачивание любой из половин вдоль проксимо-дистальной оси невозможно.
Головы крупных гиенодонов непропорционально большие — длина головы соотносится с общей длиной тела в пропорции примерно 1 : 4. Тяжелую голову поддерживала довольно короткая (короче черепа) и очень сильная шея. Мощная шея была также необходима, чтобы удерживать схваченную зубами и вырывающуюся жертву. Судя по строению шейного отдела позвоночника, у гиенодонов была высоко посаженная голова, то есть их шея располагалась почти вертикально. Вытянутое туловище заканчивается достаточно длинным хвостом.

Череп с нижней челюстью гиенодона (Hyaenodon crucians) из отложений формации Брюл в штате Южная Дакота, США.
Зубы
Зубная формула составляет I3/i3, C1/c1, P4/p4, M2/m3. Резцы и предкоренные представлены в полном числе. Р1-2 сжаты по длине. Р3-4 снабжены внутренними бугорками. Р4 симметричный, с большим срединным внутренним выступом. М1 обычно меньше предкоренных. М2 удлиненный, режущий. М3 отсутствует. Пара- и метакон на М1 слиты почти, на М2 — вполне. Передний внутренний выступ верхних коренных, несущий протокон, рудиментарный или отсутствует, метастиль сильно удлинен. Нижние предкоренные сжатые, с острыми средними буграми. Нижние коренные удлиненные, с добавочными острыми буграми. m1-2 с очень маленьким талонидом, m3 — с зачаточным или без него. Метаконида на коренных нет. Параконид m3 значительно короче протоконида и иногда значительно ниже его.
В отличие от большинства современных наземных Carnivora, обладающих только одним набором режущих (т. н. хищнических) зубов (P4/m1), гиенодоны имели целый ряд зубов, участвующих в разделке животного корма, начиная с молочных, которые в дальнейшем передавали эту функцию постоянным. У этих хищников общая длина режущего зубного ряда (т. е. зубов, участвующих в разделке корма) всегда составляет в среднем около 30 % от общей длины челюсти, независимо от размера и возраста животного. Коренные зубы гиенодонов имеют длинные режущие лезвия, образовавшиеся за счет слияния пара- и метаконов, увеличения метастиля на верхних коренных зубах и редукции метаконида и талонида на нижних.
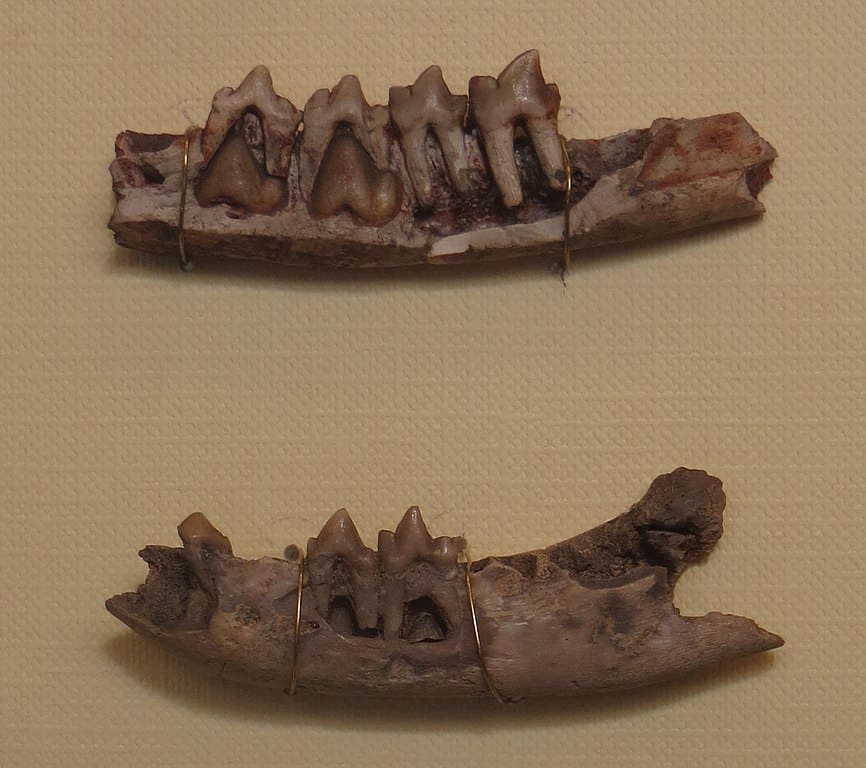
Фрагменты нижних челюстей гиенодона (Hyaenodon filholi, = Hyaenodon vulpinus) с молочными и постоянными щечными зубами.
Гиенодоны обладали массивными предкоренными и специализированными к разрезанию мяса коренными M2/m3, аналогичными хищническим зубам гиен (которые появились лишь в миоцене). Лезвиеподобные структуры тригонидов коренных зубов, образованные паракристидами, расширились за счет утраты метаконидов. Позднеэоценовые птеродоны (Pterodon) также приобрели режущие хищнические зубы подобным образом, и по этой причине долгое время считались состоящими в тесном родстве с гиенодонами. Однако в дальнейшем было установлено, что утрата метаконида у птеродонов произошла независимо.
У Carnivora апикобазальная ось режущего края верхнего хищнического зуба (P4) остается неизменной на протяжении всей жизни животного. У гиенодонов же апикобазальные оси М1-2 в течение жизни проворачивались лингвально, так что у старых особей их режущие края проходят почти под прямым углом к корням, а не параллельно им, как у современных хищных; эмалевые коронки при этом практически полностью стерты. Благодаря такой особенности хищнические зубы, стачиваясь, постоянно соприкасались и самозатачивались, что обеспечивало сохранение их остроты. Эта специализация, очевидно, также способствовала увеличению средней продолжительности жизни гиенодонов. Подобная особенность также характерна для Patriofelis и Hemipsalodon.
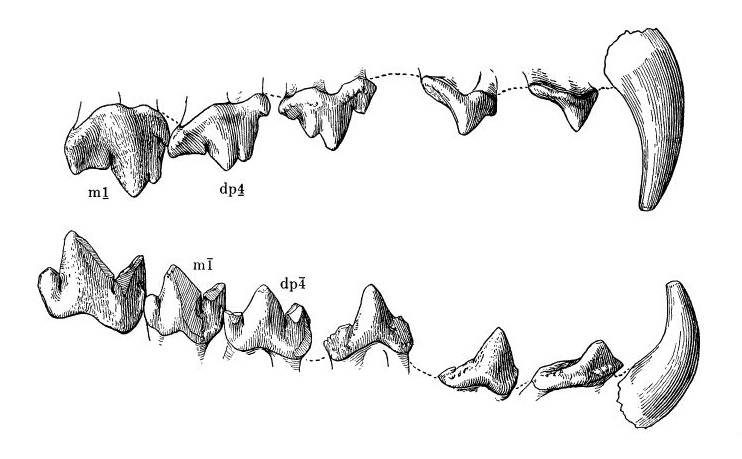
Прорисовки верхнего и нижнего правых зубных рядов молодой особи гиенодона (Hyaenodon cruentus).
У современных Carnivora постоянным хищническим зубам P4/m1 соответствуют молочные DP3/dp4. У них существует большая разница в размерах молочных и постоянных зубов. С другой стороны, у гиенодонов уже ювенильные особи обладали набором молочных зубов и постоянными m1. Для эффективного функционирования молочного хищнического зуба DP4 возникла необходимость его смыкания с нижним зубом, что привело к прорезыванию постоянного m1 одновременно с молочными в самом раннем возрасте. Как правило, у гиенодонов этот зуб сильно истерт или преждевременно утерян. При снашивании молочных зубов и увеличении длины челюсти смена зубов у гиенодонов начиналась с прорезывания M1/m2 и передачи режущих функций зубам, находящимся ближе к челюстному суставу. С прорезыванием M2 и m3 молочные DP3-4 и dp4 утрачивались полностью. M2 и m3 — самые крупные коренные, и если бы они прорезались одновременно, соотношение длины режущего зубного ряда и длины челюсти было бы нарушено.
При стирании M1 его режущий край уменьшался в длину, но компенсировался в начале апикальной части M2 в зубном ряду. Дистальная грань M2 расположена под углом и соответствует проксимальной грани M1, базальной к лезвию метастиля. При дальнейшем прорезывании M2, М1 продолжал стираться, пока у взрослого животного метастиль на этом коренном не утрачивался полностью и основная часть режущей функции возлагалась на увеличенный M2. Нижний коренной m3 гиенодонов способствовал постепенной аккумуляции режущей силы, проворачиваясь при прорезывании вдоль букколингвальной оси. Сначала прорезывалась задняя, а затем и передняя часть зуба. Проворачивание продолжалось до полного прорезывания коронки и выхода ее в окклюзионную позицию. Как и у передних зубов, расположение выемки и гребня довершает надлежащий вид этого зуба, по форме напоминающего m1 кошек.
Отношение между длиной зубного ряда и суставной длиной обусловливает необычный порядок прорезывания Р3/р3. Оба этих зуба проксимодистально удлинены и резко добавляют около 10 % к общей длине зубного ряда. Возможно, если бы эти зубы прорезывались по всей длине, нарушалась бы механика закрывания челюстей. Входя под углом в зубной ряд, а затем проворачиваясь вдоль своих апикобазальных осей, эти зубы были предназначены для дробления и удержания пищи. Европейская линия гиенодонов отличается от североамериканской главным образом порядком смены зубов. У североамериканских представителей рода Р1 прорезывается раньше М1, тогда как европейские формы демонстрируют более раннее прорезывание М1. p3 у европейских форм прорезался одновременно с m3 и перед Р4. У североамериканских видов p4 прорезывался ранее p3. Молодым особям требовалось порядка 3-4 лет, чтобы их коренные полностью прорезались. Европейские виды начинали рано использовать клыки, прорезающиеся примерно в одно время с p2, что также резко контрастирует с порядком смены зубов видов из Северной Америки, у которых клыки сменялись в последнюю очередь. Это свидетельствует о широком разделении линий развития европейских и североамериканских представителей рода.
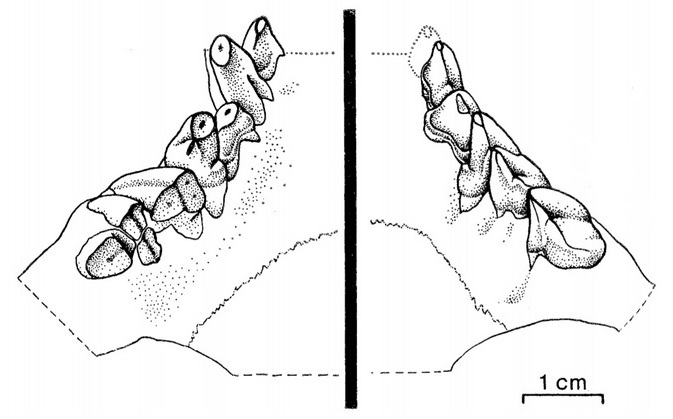
Прорисовки жевательных поверхностей верхних щечных зубов гиенодона (Hyaenodon crucians): слева — старое животное, у которого режущая кромка проходит по корням зубов, справа — молодое животное.
Таким образом, в эволюции линии рода Hyaenodon имела место цепь последовательных изменений в черепной и зубной морфологии, которая в конечном итоге привела к увеличению размеров, появлению более эффективных режущих зубов и возможности широко раскрывать пасть. Предполагается, что удлинение основных лезвий на хищнических зубах, метакрист (постметакрист) на верхних и паракристидов на нижних коренных, а также переориентация этих гребней от наклонных до почти мезиодистальных, дали гиенодонам возможность справляться с большой добычей, что в т. ч. привело к общему увеличению размеров их тела. Благодаря утрате M3 стало возможным расширение в заднем направлении хищнических зубов M2/m3, столь характерных для гиенодонов, что в конечном итоге привело к изменениям в строении челюстного сустава и всего комплекса жевательной мускулатуры. Эти черты, несомненно, сделали гиенодонов одними из наиболее серьезных хищников своего времени.
Для гиенодонов был характерен половой диморфизм, выражавшийся главным образом в более длинных клыках самцов.