Происхождение и родственные связи
Своеобразие и гигантизм гигантопитеков, а также фрагментарность их ископаемых остатков долгое время затрудняли определение места этих обезьян на эволюционном древе приматов. Новые находки и методы исследований позволили внести некоторую ясность в этот вопрос.
На сегодняшний день установлено, что гигантопитеки входят в подсемейство понгин (Ponginae) семейства гоминид (Hominidae). Время отделения этой группы от основного ствола человекообразных обезьян восходит к середине миоценовой эпохи. Гоминиды, видимо, появились в Африке и расселились сначала в Европе, а затем и в Азии. В Африке и Европе они вымерли в конце миоцена, а в Азии продолжают существовать и по сей день в лице своих последних представителей — орангутанов (Pongo). Среди вымерших гоминид имелись как небольшие, так и среднего роста или крупные обезьяны. Для них были характерны укороченный (по сравнению с другими человекообразными обезьянами) лицевой отдел черепа и в большей степени V-образная (а не U-образная) зубная дуга, небольшие резцы и крупные щечные зубы. Одна из морфологических особенностей гоминид — толстая, складчатая на жевательной поверхности эмаль.
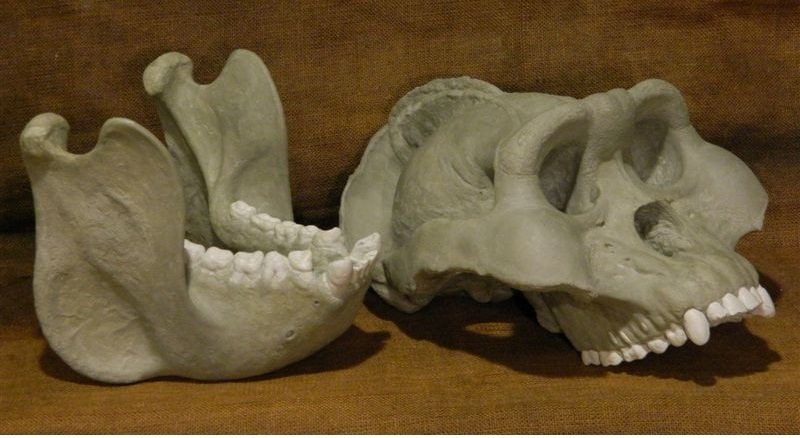

Реконструированные череп и нижняя челюсть гигантопитека (Gigantopithecus blacki).
Строение зубочелюстного аппарата гигантопитеков очень своебразно. Оно в определенной степени имитирует таковое гоминин (Homininae), в особенности массивных форм австралопитеков (Australopithecus). Это выражается прежде всего в массивной, высокой и относительно короткой нижней челюсти, узких диастемах, небольших вертикально посаженных резцах, уменьшенных коронках клыков, моляризованных предкоренных, а также крупных коренных с низкими округлыми бугорками и толстой эмалью. Все перечисленные специализации могут быть выведены в качестве эволюционных приобретений гигантопитеков, хотя увеличенные коренные с относительно короткими дентиновыми рогами и мощным слоем эмали с неглубокими впадинами и низким окклюзионным рельефом, возможно, представляют собой исходное состояние гоминидов (например, присутствующее у их среднемиоценовых родов Griphopithecus и Kenyapithecus), впоследствии претерпевшее у понгин прогрессивное развитие.
Благодаря сходным морфологическим чертам гигантопитеков в свое время даже сближали с австралопитеками и прочими гомининами. Однако, несмотря на поразительное сходство, при рассмотрении каждого конкретного признака становится ясно, что оно носит чисто поверхностный характер. Вместо указания на тесную родственную связь с гомининами, общие с ними черты строения правильнее интерпретировать как конвергентные приобретения в ответ на структурно-функциональные требования к зубочелюстной системе в похожих жизненных условиях. Формирование укороченной и массивной нижней части лицевого отдела, уменьшение размеров резцов и клыков, моляризация предкоренных и увеличение коренных зубов, по-видимому, являлось общей тенденцией среди позднемиоценовых гоминоидов и, очевидно, развивалось независимо в нескольких линиях. По меньшей мере 2 такие линии дожили до плио-плейстоцена и дали начало Homo и Gigantopithecus соответственно.

Портретная реконструкция самца гигантопитека (Gigantopithecus blacki).
В настоящее время исследователи располагают достоверными сведениями о том, что у некоторых миоценовых человекообразных обезьян в связи с переходом к наземному образу жизни произошли значительные изменения в строении зубной системы и всего скелета. Видимо, эти ветви эволюционировали параллельно по пути "очеловечивания". Большинство из них все более специализировались и вымерли, а другие во времена плиоцена "встали на ноги", что лишь у одной из групп африканских гоминид приобрело принципиальное значение (при сборе корма передними конечностями и дальнейшем использовании естественных и искусственных орудий) и привело к появлению разумных существ.
Известно, что эволюция гоминид была связана с постепенным приспособлением к жизни в саваннах и лесостепях (подтверждением тому служат и некоторые признаки скелета верхних и нижних конечностей) и питанию сухой и грубой пищей. В конце миоцена произошло сокращение площадей тропических лесов, и в основном как раз в тех районах, где в условиях жесткой конкуренции обитали 2 группы человекообразных обезьян — понгины и куда более многочисленная полиморфная группа гоминид типа дриопитеков (Dryopithecus). Именно конкуренцией и объясняется постепенный переход понгин в экологические ниши, нетипичные для большинства других человекообразных обезьян. Вершин специализации к новым условиям достигли именно гигантопитеки.

Реконструкция внешнего облика гигантопитека (Gigantopithecus blacki). Автор — Concavenator.
В качестве непосредственных предшественников и наиболее вероятных предков гигантопитеков рассматриваются сивапитеки (Sivapithecus) — успешный род средних и крупных понгин из среднего и позднего миоцена Южной и Восточной Азии. Скорее всего, от сивапитеков произошли и очень похожие на них орангутаны, впоследствии сумевшие приспособиться к сугубо древесному образу жизни в тропических лесах. Впрочем, нельзя также исключать более теснуюю связь гигантопитеков с другим родом понгин из позднего миоцена Китая — небольшими люфенгпитеками (Lufengpithecus). Протеомный анализ дентирна и эмали коренного зуба Gigantopithecus blacki из пещеры Чуйфэн возрастом 1,9 млн. лет подтвердил, что этот примат родственен орангутанам и их эволюционные пути разошлись около 12-10 млн. лет назад.
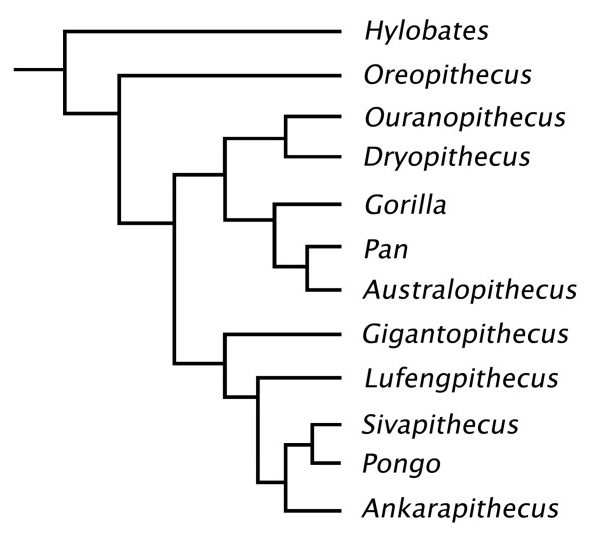
Кладограмма, отображающая возможные родственные связи Gigantopithecus (по Бегуну, 2007).
Не исключено, что гигантопитеки представляют собой один из нереализовавшихся путей гоминизации. Препятствием для их "параочеловечивания" явилась, видимо, специализация к растительноядности (что следует из строения зубов несмотря на предположения о периодическом потреблении мяса) и гигантские размеры тела. У столь крупных приматов практически не было естественных врагов. А безопасность расслабляет, не давая стимулов для развития интеллекта.
Тафономия и палеоэкология
На сегодняшний день остатки Gigantopithecus blacki известны из 16 китайских местонахождений. Наиболее северные из них — Лунгупо и Лунгудонг — расположены к югу от течения реки Янцзы, а самое южное находится на острове Хайнань в Южно-Китайском море. Одни из древнейших известных остатков происходят из пещеры Байконг и датируются возрастом 2 млн. лет, а достоверные наиболее молодые найдены в пещере Хэй и имеют возраст 380-310 тыс. лет. Gigantopithecus blacki был характерным представителем раннеплейстоценового гигантопитекового и среднеплейстоценового пандо-стегодонового фаунистических комплексов, имевших распространение на юге Китая и в близлежащих областях. Северная граница распространения Gigantopithecus blacki примерно совпадала с таковой плейстоценовых орангутанов.
Для юго-востока Азии обычен рельеф с круто возносязимися известняковыми останцами, пронизанными карстовыми пещерами и вымоинами. В этих пещерах и обнаружены остатки Gigantopithecus blacki, а также богатая сопутствующая фауна позвоночных. Челюсти и зубы гигантской обезьны обычно залегают в слоях желтой песчано-глинистой известковой брекчии — типе пещерных отложений, в которых рыхлые породы и камни сцементированы кальцитом. Происхождение наслоений, по-видимому, связано с чередованием влажных и сухих периодов, в связи с чем в пещеры проникало то большее, то меньшее количество влаги с расстворенными известковыми солями. Что касается совместного залегания в них фрагментарного материала гигантопитеков и других крупных "непещерных" млекопитающих, то он в большинстве случаев представляет собой остатки добычи хищников (например, гиен и крупных кошек), притаскивающих в пещеры части туш. На это указывает сам набор костей: они принадлежат либо молодым, либо старым животным, т. е. таким, которые легче могли стать жертвой.

Гора Динху в провинции Гуандун на юге Китая — ныне существующий вероятный аналог раннеплейстоценовых метообитаний гигантопитека (Gigantopithecus blacki).
Кости, оставшиеся от пиршества плотоядных, зачастую обгрызались дикобразами (Hystrix spp.), обычными в плейстоценовых фаунах Китая. Из костной ткани они получают минеральные вещества, необходимые для роста защитных игл. Эти грызуны способны затаскивать элементы скелета в свои подземные логова и потреблять их целиком, за исключением твердых, покрытых эмалью зубных коронок. Это может объяснить, почему зубы обычно встречаются в большом количестве, а другие остатки пропадаются так редко. Помимо заноса в подземные полости хищниками и дикобразами, кости и зубы могут находиться во вторичном залегании, смываясь с поверхности земли водными потоками и попадая в карстовые трещины в результате эрозии почвы. Вкупе эти факторы совсем не благоприятствовали целостности остатков Gigantopithecus blacki гигантопитеков, но поспособствовали их сохранению хотя бы в фрагментарном виде. Стоит отметить, что орангутаны, встречающиеся в плейстоценовых фаунах южного Китая почти вдвое чаще, представлены исключительно зубным материалом.
Территория южного Китая в раннем и среднем плейстоцене представляла собой пересеченную невысокими горами равнину — травянисто-кустарниковую саванну. Горы и горные долины покрывали широколиственные леса субтропического типа. На Хайнани, в Индокитае, на Малаккском полуострове и Яве имелись тропические муссонные леса. Состав сопутствующей фауны, палинологические данные, а также количественный анализ содержания изотопов углерода и кислорода в эмали зубов гигантопитеков раннеплейстоценового возраста свидетельствуют, что они населяли хорошо увлажненные широколиственные леса с закрытым пологом и густым подлеском из кустарников и травянистых растений. Подходящим современным аналогом с аналогичным изотопным профилем углерода является биосферный заповедник Динхушань в провинции Гуандун на юге Китая, который представляет собой субтропический муссонный вечнозеленый широколиственный лес со средней годовой температурой 21 °C и среднегодовым количеством осадков 1900 мм.

Животно-растительное сообщество, существовавшее в среднем плейстоцене на территории Северного Вьетнама. На переднем плане — первобытные люди Homo еrectus, на противоположном берегу — группа Gigantopithecus blacki. Автор — Дж. Сиббик.
Палинологический анализ раннеплейстоценовых слоев пещеры Цюэцюэ в Гуанси-Чжуанском автономном районе, содержащих остатки Gigantopithecus blacki, указывает на климат от теплого умеренного до субтропического. В окрестностях пещеры произрастали смешанные лиственные и вечнозеленые широколиственные леса с преобладанием березы, дуба и каштанника, а также нижний ярус из различных кустарников и травянистых растений, включая папоротники, травы и двудольные.
Хотя северная граница субтропической зоны на протяжении плейстоцена могла пульсировать в ответ на ледниковые циклы, вполне вероятно, что ее растительность и климатический режим были аналогичны существующим сегодня. В пещерных местонахождениях южного Китая и близлежащих областей преобладают таксоны индо-малайской фауны, тогда как фауна более северных местонахождений имеет переходной характер с примесью палеарктических элементов. Весьма разнообразное сообщество приматов, существовавшее в плейстоцене на юге Китая рядом с Gigantopithecus blacki, включает в себя представителей родов Macaca (несколько видов), Rhinopithecus, Pygathrix, Trachypithecus, Nomascus и Pongo. Хотя в современной фауне аналогов этому сообществу приматов нет, наиболее близко соответствующее ему обитает в тропических лесах на севере Вьетнама.
Образ жизни и поведение
Ввиду значительной фрагментарности находок предполагаемый способ передвижения гигантопитеков реконструируется путем логических умозаключений. Само собой разумеется, что крупные размеры препятствовали лазанию по деревьям или сильно ограничивали его. К тому же, строение зубов и челюстей гигантопитеков свидетельствует о питании отнюдь не древесным кормом. Не могли эти приматы и передвигаться на задних конечностях — их лодыжка и стопа хватательного типа попросту не выдержали бы настолько значительных нагрузок. Таким образом, напрашивается вывод о квадропедальном (четвероногом) типе передвижения гигантопитеков, при котором масса распределялась на все четыре конечности. Вероятно, на задние конечности гигантопитеки поднимались нечасто (например, чтобы дотянуться до пищи или отпугнуть врага) и оставались в таком положении недолго. Вероятно, на деревья крупные особи не забирались, поскольку ветви просто не выдержали бы их огромного веса.

Реконструкция внешнего облика гигантопитека (Gigantopithecus blacki). Автор — З. Буриан.
Учитывая специфику строения зубочелюстного аппарата, существует значительный интерес к реконструкции пищевого поведения гигантопитеков. Был использован ряд методов и подходов, каждый из которых дает важные ключи к разгадке возможных диетических предпочтений Gigantopithecus blacki, но трудность заключается в том, чтобы интегрировать и согласовать результаты иследований в единую функционально-поведенческую модель, согласующуюся с различными линиями доказательств.
Массивная нижняя челюсть с мощным симфизом, моляризованные предкоренные и огромные коренные зубы с низкими закругленными бугорками и длинными корнями свидетельствуют о их ярко выраженной жевательной и дробящей специализации, которую можно встретить у животных, потребляющих объемные массы твердого и волокнистого растительного материала: семян, стеблей, жестких побегов бамбука, клубней и т. д. Имеющийся на щечных зубах толстый слой эмали и характер его износа подразумевает попадание вместе с пищей абразивных частиц — например, прилипших к побегам бамбука песчинок. В то же время, низкий рельеф и слабое развитие эмалевых гребней свидетельствует о плохой приспособленности щечных зубов к эффективной обработке листьев и, в особенности, травинок. Резцы кажутся слишком мелкими для обработки относительно крупных объектов (например, плодов), но характер износа режущего края подразумевает их интенсивное использование. Подобным строением нижней челюсти и зубов обладает почти исключитетельно бамбукоядная большая панда (Ailuropoda melanoleuca), что породило предположения о схожем рационе у гигантопитеков. Приматы столь значительных размеров нуждались в обильном источнике питательных веществ, на роль которого как раз хорошо подходит бамбук. Однако сходное строение зубочелюстного аппарата характерно и для многих современных обезьян, питающихся в основном мягкими листьями или семенами. Кроме того, более толстая эмаль и гипсодонтия у гигантопитеков может указывать на иную по сравнению с большой пандой функциональность зубов.

Портретная реконструкция гигантопитека (Gigantopithecus blacki). Автор — З. Буриан.
На зубной эмали обезьян имеются весьма характерные царапины и повреждения, образующиеся при употреблении жесткого корма, насыщенного фитолитами — микроскопическими кремневыми включениями, отлагающимися в тканях многих растений. У Gigantopithecus blacki количество микроповреждений эмали и их размеры заметно меньше, чем у орангутанов, и примерно такие же, как у шимпанзе, что может указывать на столь же разнообразное питание. Изучение содержания в зубной эмали доли углерода-13 свидетельствует о наличии в рационе плодов, листьев, молодых побегов и другой лесной растительности. Количественный анализ изотопов кислорода позволяет сделать вывод, что гигантопитеки потребляли относительно больше низкорасположенной растительности, такой как стебли, корни и травы, чем орангутаны.
Особый интерес представляет изучение самих фитолитов, выявленных на некоторых зубах Gigantopithecus blacki. Они сосредоточены преимущественно на жевательных поверхностях коренных и порой сохранились достаточно хорошо для того, чтобы их можно было таксономически идентифицировать. Некоторые из фитолитов, по-видимому, происходят из вегетативных частей трав (например, побегов бамбука), в то время как большинство напоминает таковые из волосковых клеток плодов тутовых (Moraceae). Последние представляют собой семейство кустарников и деревьев, в основном произрастающих в субтропических и тропических регионах и дают большое количество съедобных плодов, таких как шелковица, инжир, хлебное дерево, дуриан и баньян. Очевидно, Gigantopithecus blacki с удовольствием лакомился сезонными фруктами в пору их созревания. Можно предположить, что плоды были такого размера и типа, которые позволяли эффективно пережевывать их массивными щечными зубами с незначительной предварительной обработкой небольшими резцами или вообще без нее. Наличие в зубном камне крахмальных зерен указывает на то, что могли поедаться и клубни. Бамбук, по-видимому, не являлся первостепенным компонентом рациона, а саванные травы не употреблялись в пищу совсем. Таким образом, гигантопитеки питались довольно разнообразным растительным кормом, сосредоточенном в нижнем ярусе леса, а также упавшими с деревьев плодами. Помимо покрытосеменных, на лесной подстилке произрастают многочисленные съедобные грибы и папоротники, которые также могли поедаться гигантопитеками. Судя по всему, они представляли собой своего рода азиатский аналог горилл.

Слева: фитолит, содержавшийся в растительной пище гигантопитека (Gigantopithecus blacki) и застрявший в его зубной эмали.
Справа: реконструкция способа питания гигантопитека (Gigantopithecus blacki).
Дополнительным подтверждением того, что Gigantopithecus blacki являлся преимущественно вегетарианцем служит и то, что часть его обнаруженных зубов (11,5 %) сильно поражена кариесом, который мог возникнуть из-за содержания в пище большого количества крахмала и недостатка кальция и фосфора. Интересно, что похожая ситуация наблюдается и у большой панды. Впрочем, она могла быть вызвана и высоким содержанием сахара в потребляемых гигантопитеками плодах подобно тому, как это происходит у шимпанзе. У других ископаемых приматов и древних людей кариес редок. Также у Gigantopithecus blacki фиксируется относительно высокая частота заболеваемостью гипоплазией, проявляющейся в неравномерном развитии слоя зубной эмали. Это может быть обусловлено периодическим недоеданием, хотя другие факторы также способны вызывать подобное состояние. Высказывалось предположение, что у гигантопитеков гипоплазия была связана с недостаточным питанием во время нехватки основного корма — бамбука, для которого характерно периодическое массовое усыхание. С подобной проблемой в 1970-х гг. столкнулись большие панды — этот неблагоприятный экологический фактор сократил их и так редкую популяцию до менее 1000 особей.
Крупные размеры гигантопитеков и их видимое сходство с предками людей наводили ранних исследователей на мысль, что эти огромные обезьяны не могли прокормиться исключительно растительностью. Действительно, некоторые факты косвенно указывают как минимум на их всеядность. Об этом в частности может свидетельствовать нахождение остатков Gigantopithecus blacki в сопровождении многочисленных костей копытных в пещерах, расположенных высоко на крутых склонах (например, в гроте Сяоянь на горе Лянь Чи-Шань). Предполагается, что они были недоступны для хищников (что, впрочем, сомнительно в отношении представителей семейства кошачьих) и туши травоядных сюда доставлялись именно гигантопитеками. Более того, иногда в а общих с Gigantopithecus blacki слоях обнаруживаются очень большие каменные орудия, возможно, принадлежавшие этим приматам. Судя по размерам челюстей, их головной мозг был заметно больше, чем у горилл, и находился в пределах значений человека выпрямленного (Homo erectus). И все же гигантопитеки являлись преимущественно растительноядными, а в труднодоступные пещеры их остатки почти наверняка попали в результате деятельности хищников.

Группа кормящихся гигантопитеков (Gigantopithecus blacki). Автор — М. Антон.
Представляется вероятным, что у столь крупных животных, как гигантопитеки, испытывавших ежедневную потребность в значительном количестве пищи, образование больших групп было невозможно. Скорее всего, они, подобно современным горным гориллам, жили небольшими семейными группами численностью в 5-10 особей. Вероятно, большую часть времени гигантопитеки бродили по зарослям в поисках пищи, а насытившись — отдыхали. Представляется вероятным, что ели они сидя, перебирая пищу и отправляя ее в рот руками или пригибая к себе стебли растений, как это делают гориллы.
Анализ ископаемых остатков показывает, что количественное соотношение особей различных полов в группе было примерно одинаковым. Из 88 особей Gigantopithecus blacki, чьи остатки были собраны в пещерах южного Китая, 41 оказались самцами, а 47 — самками. Подобное соотношение самцов и самок довольно обычно для крупных современных приматов и достоверно установлено, например, для горной гориллы (Gorilla beringei). Однако конкурировали ли между собой самцы за обладание несколькими самками, точно неизвестно. Можно судить и о возрастном составе погибшей популяции, в которой взрослые (но не старые) особи составляли примерно 56 %, молодые неполовозрелые — 24 %, детеныши — 6 %, очень старые особи — 15 %. Такой возрастной состав погибших животных нетипичен для нормально существующей популяции млекопитающих, обычно процент погибших взрослых особей всегда меньше.

Самец гигантопитека (Gigantopithecus blacki) отпугивает от своей семейной группы случайно встретившуюся пантеру Зданского (Panthera zdanskyi). Автор — В. Симеоновски.
Очевидно, гигантопитеки были довольно медлительными и флегматичными животными. Скорее всего, хищники не часто осмеливались нападать на взрослых обезьян, особенно на громадных самцов, которые благодаря габаритам и недюжинной силе вполне могли постоять за себя и за своих сородичей. Высокая степень полового диморфизма подразумевает относительно интенсивную конкуренцию между самцами, но учитывая относительно небольшие размеры клыков у обоих полов можно предположить, что их демонстрация не играла большого значения при выяснении иерархических отношений.
Вымирание: сроки и причины
Долгое время считалось, что более поздний представитель рода — Gigantopithecus blacki — вымер порядка 400 тыс. лет назад. Однако по последним результатам датировки, включающей электронный спиновый резонанс и измерение относительного содержания долгоживущих изотопов урана-238, он окончательно исчезает лишь к концу среднего плейстоцена, где-то между 295 и 215 тыс. лет назад. Приблизительно в то же время существенно сокращается ареал большой панды. Это животное исчезает с большей части занимаемой им до этого территории, в частности перестает встречаться на островах Юго-Восточной Азии. В дальнейшем область ее распространения вообще ограничивается труднодоступными высокогорными районами китайской провинции Сычуань. На сегодняшний день большая панда находится на грани исчезновения. Со значительной долей уверенности можно утверждать, что основным неблагоприятным фактором для выживания данного вида являлось прямое преследование со стороны человека.
Но что же привело Gigantopithecus blacki к гибели? Этот вопрос остается открытым. Ранее считалось, что главная причина кроется в высокой степени специализации этого примата. Так как высокоспециализированные формы зачастую утрачивают способность приспосабливаться к изменяющимся условиям, они попадают в эволюционный тупик и рано или поздно прекращают свое существование. В процессе эволюции у многих групп млекопитающих (копытных, хоботных и др.) наблюдалась тенденция к постепенному увеличению размеров тела, а иногда и появлению гигантизма. Как правило, это связано с односторонней адаптацией — пассивным приспособлением к внешним условиям. Хотя увеличение размеров тела дает животным биологические преимущества при конкуренции с другими видами, в частности в борьбе с хищниками, но часто оказывается одной из главных причин исчезновения при существенных изменениях окружающей среды. Известно немало примеров того, как виды, становясь гигантами, оказываются на грани вымирания.
Среди главных причин исчезновения Gigantopithecus blacki предполагались также конкуренция за источники питания с большой пандой и палеолитическим человеком. Все трое с удовольствием употребляли в пищу молодые ростки бамбука, что в принципе могло привести к уменьшению площадей, занимаемых этими растениями. Кроме того, необходимо принимать во внимание периодически случающуюся массовую гибель бамбука под влиянием невыясненных до конца факторов. Если гигантопитеки в самом деле были столь зависимы от бамбука (что сегодня подвергается сомнению), то они действительно могли не выдержать возросшей конкуренции со стороны других видов за сократившиеся основные пищевые ресурсы. Что касается панд, то они были гораздо мельче гигантопитеков и поэтому имели больше шансов выжить. В свете этого предположения примечательно, что в Китае ископаемые остатки древнего человека и большой панды нередко сопровождают остатки гигантопитека, нашли их рядом и в Там-Хуен во Вьетнаме (сейчас большая панда на территории этой страны не встречается).
На протяжении всего среднего плейстоцена вследствие увеличения сезонности и силы муссонов, а также тенденции к похолоданию, леса в Юго-Восточной Азии отступали к югу, так что преобладающим ландшафтом к позднему плейстоцену здесь становится саванна. Увеличение размера зубов Gigantopithecus blacki с течением времени, а также появление морфологических изменений зубных рядов у поздней популяции из пещеры Хэцзян может указывать на то, что этот примат приспосабливался к новому рациону из более грубой растительности в ответ на меняющиеся условия окружающей среды. Однако убедительных доказательств экологического сдвига, который привел Gigantopithecus blacki к вымиранию, нет. Судя по всему, другие его современники из числа приматов в значительной степени не пострадали и перешли в поздний плейстоцен. При этом нельзя исключать того, что общие экологические изменения являлись незначительными и определенные аспекты экосистемы были затронуты с однозначно неблагоприятными последствиями лишь для Gigantopithecus blacki.
Наконец, не исключается возможность того, что древние люди охотились на гигантопитеков. В Гуанси-Чжуанском автономном районе и провинции Хубэй в общих слоях с Gigantopithecus blacki присутствие Homo фиксируется в виде его костных остатков и каменных орудий. Кроме того, разрозненные зубы Homo еrectus могут сопровождаться остатками гигантопитека во вьетнамской пещере Там-Хуен. Таким образом, доподлинно известно, что на протяжении длительного времени Gigantopithecus blacki жил по соседству с древним человеком. Также установлено, что его численность существенно сократилась одновременно с распространением в регионе человека прямоходящего около 1-0,9 млн. лет назад, хотя существенных изменений климатической обстановки и характера растительности на юго-востоке Азии в этот период не происходило (примечательно, что Африке приблизительно в то же время исчезают массивные австралопитеки). Очевидно, мощь гигантской обезьяны не являлась достаточным аргументом против стремительно совершенствующихся орудий и способов охоты двуногого хищника. Крупные размеры и наземный образ жизни Gigantopithecus blacki делали его желанной и легкой добычей. С другой стороны, следы человеческого присутствия на юге Китая в раннем и среднем плейстоцене являются достаточно скудными. Кроме того, первобытный человек предпочитал селиться в саванне, тогда как гигантопитек обитал в лесу и какие-либо археологические свидетельства охоты на него отсутствуют.

Столкновение группы азиатских представителей вида человека прямоходящего (Homo еrectus) с гигантопитеком (Gigantopithecus blacki). Автор — З. Буриан.
Сегодня представляется наиболее вероятным, что вымирание Gigantopithecus blacki было вызвано сочетанием целого ряда неблагоприятных факторов. По-видимому, важнейшим из них было изменение климата, вызвавшее критическое сокращение предпочитаемых местообитаний к концу среднего плейстоцена. На протяжении всего среднего плейстоцена вследствие увеличения сезонности и силы муссонов, а также тенденции к похолоданию, леса в Юго-Восточной Азии отступали к югу, так что доминирующим ландшафтом к позднему плейстоцену здесь становится саванна. На таком фоне деятельность древнего человека могла стать последней каплей, окончательно нарушившей шаткое равновесие существования огромной обезьяны. К примеру, если люди активно потребляли те же плоды, что и гигантопитеки, во время периодического отмирания бамбука они просто могли оставить их без пищи. Это в сочетании с конкуренцией со стороны больших панд и спорадических охот могло уменьшить численность гигантопитеков ниже уровня, необходимого для выживания их популяции.
Домыслы и спекуляции
По степени внимания, которое уделяли находкам гигантопитеков журналисты, они могут сравниться, пожалуй, лишь с древнейшими предшественниками человека, остатки которых были обнаружены в Восточной и Южной Африке. Мысль о гигантских обезьянах неожиданно вдохновила кинорежиссеров, создавших серию фильмов о чудовищных обезьянах-монстрах, доживших до наших дней в джунглях на островах Юго-Восточной Азии.
Однако больше всего гигантопитеки "прославились" тем, что с ними часто связывают многочисленные сообщения о т. н. снежном человеке, с которым эти обезьяны на самом деле не имеют ничего общего. Несмотря на это, зачастую утверждается, что гигантопитеки не вымерли и продолжают существовать в труднодоступных районах Азии (йети, алмас и т. п.) и других потаенных уголках нашей планеты. Также было высказано ничем не обоснованное предположение, что гигантопитеки в свое время пересекли Берингов перешеек и проникли в Новый Свет, где сегодня известны как бигфуты, саскватчи и др. Несмотря на наличие значительного количества наблюдений и отпечатков следов (часто очень различающихся между собой), на сегодняшний день бесспорных доказательств существования снежного человека нет. Несомненно, он является интереснейшим феноменом, который должен находиться в ведении сравнительной мифологии, культурной антропологии и психологии, но, вероятно, никоим образом не естественных наук. И дело не только в том, что ученые пока не получили в свои руки живого гоминоида, его труп или хотя бы четко идентифицированные волосы. Не мифическая, а реально существующая жизнеспособная популяция крупных приматов непременно должна воздействовать на окружающую среду, чего, однако, не наблюдается в случае со снежным человеком.