Состав
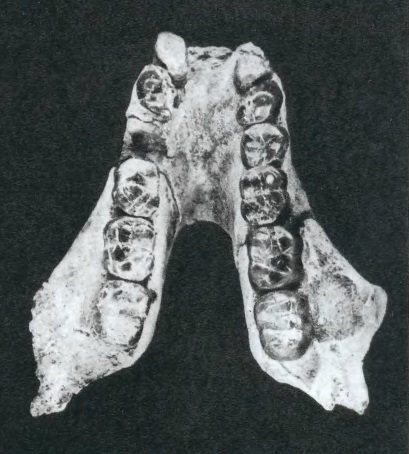
Нижняя челюсть гигантопитека (Gigantopithecus giganteus) из Хариталиангара, Индия.
Gigantopithecus giganteus (Pilgrim, 1915) Kay, 1982 — вид, известный из позднего миоцена (порядка 8,6-6,3 млн. лет назад) Сиваликских холмов на севере Индии (штат Химачал-Прадеш; местонахождение Хариталиангар) и Пакистана (плато Потвар, местонахождения Сети-Нагри и Алипур). Синонимы: Sivapithecus indicus Pilgrim, 1910 (частью); Dryopithecus giganteus Pilgrim, 1915; Indopithecus giganteus von Koenigsvald, 1950; Gigantopithecus bilaspurensis Simons et Chopra, 1969; Dryopithecus indicus Simons et Pilbeam, 1971 (частью). Лучше всего изучена неполная нижняя челюсть с большинством зубов и без восходящих ветвей, обнаруженная к северу от Хариталиангара в отложениях, коррелирующих с пакистанской формацией Дхок-Патхан и вначале описанная в качестве Gigantopithecus bilaspurensis. Из Сети-Нагри происходит дистальный фрагмент плечевой кости, по величине и морфологически сходный с соответствующей частью плеча самки гориллы. Это единственный известный на сегодняшний день элемент посткраниального скелета гигантопитека, хотя его идентификация остается сомнительной. Размеры найденных остатков, а также их возраст и морфология позволяют предполагать, что Gigantopithecus giganteus примерно вдвое уступал по величине Gigantopithecus blacki и, вероятно, являлся его непосредственным предком.
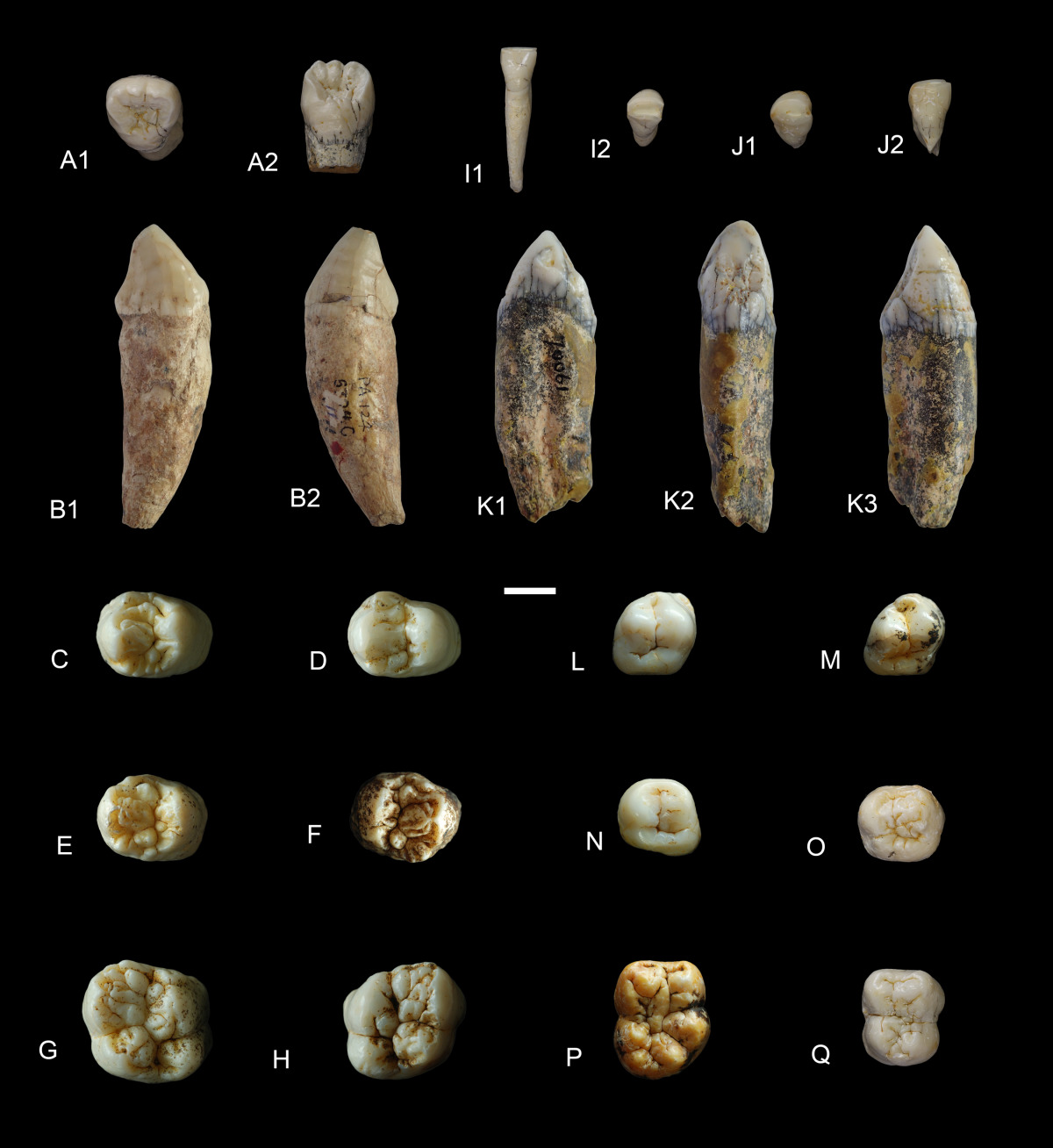
Изолированные верхние (A-H) и нижние (I-Q) зубы гигантопитека (Gigantopithecus blacki) из Китая: A — левый I1 из пещеры Яньлян, жевательная (1) и язычная (2) поверхности; B — правый C из пещеры Сяоянь, жевательная (1) и язычная (2) поверхности; C — правый P3 из пещеры Саньхэ; D — правый P3 из пещеры Саньхэ; E — правый P4 из пещеры Саньхэ; F — левый P4 из пещеры Саньхэ; G — правый M1/2 из пещеры Саньхэ; H — левый M1/2 из пещеры Саньхэ; I — левый i1 из пещеры Яньлян, жевательная (1) и язычная (2) поверхности; J — левый i2 из пещеры Яньлян, жевательная (1) и язычная (2) поверхности; K — правый с из пещеры Сяоянь, дистальная (1), язычная (2) и мезиальная (3) поверхности; L — правый р3 из пещеры Хэцзян; M — правый р3 из пещеры Саньхэ; N — левый р4 из пещеры Бойю; O — левый р4 из пещеры Яньлян; Р — правый m1/2 из пещеры Бойю; Q — левый m1 из пещеры Яньлян. Длина масштабной линейки — 10 мм.
Gigantopithecus blacki von Koenigswald, 1935 (тип) — типовой вид, известный из раннего - среднего плейстоцена (примерно 2,2-0,2 млн. лет назад) центрального и южного Китая (Гуанси-Чжуанский автономный район, провинции Хубэй, Гуйчжоу и Хайнань, а также муниципалитет Чунцин; пещеры Лунгудонг (33 зуба, ок. 2,1-1,8 млн. л. н.), Байконг (214 зубов, ок. 2 млн. л. н.), Чуйфэн (92 зуба, ок. 2-1,2 млн. л. н.), Лунгупо (16 зубов, ок. 1,96-1,4 млн. л. н.), Мохуэй (16 зубов, ок. 1,9-1,2 млн. л. н.), Бойю (43 зуба, ок. 1,8 млн. л. н.), Саньхэ (74 зуба, ок. 1,6-0,7 млн. л. н.), Сяоянь (1006 зубов и 3 частичные нижние челюсти, ок. 1,2-0,94 млн. л. н.), Цюэцюэ (22 зуба, ок. 1 млн. л. н.), Баэрян (5 зубов, ранний плейстоцен), Яньлян (22 зуба и 1 частичная нижняя челюсть, ранний плейстоцен), Булалишань (12 зубов, ок. 0,75-0,48 млн. л. н.), Синьчун (1 зуб, 0,6-0,3 млн. л. н.), Хэцзян (17 зубов, 0,4-0,32 млн. л. н.), Хэй (3 зуба, 0,38-0,31 млн. л. н.), Номошань (1 зуб, средний плейстоцен), Шуангтан (средний плейстоцен)), а также, по-видимому, севера Вьетнама (провинция Лангшон; пещера Там-Хуен (1 зуб, ок. 0,6-0,35 млн. л. н.)), Лаоса (пещера Там-Ганг), севера Таиланда (пещеры Пха-Бонг (1 зуб, средний плейстоцен) и Хад-Пу-Дай (средний плейстоцен)) и острова Ява в Индонезии (местонахождение Семоно (2 нижнечелюстных фрагмента с зубами). Синонимы: Gigantanthropus blacki Weidenreich, 1946; Giganthropus blacki Weinert, 1950. Более молодой и крупный представитель рода.
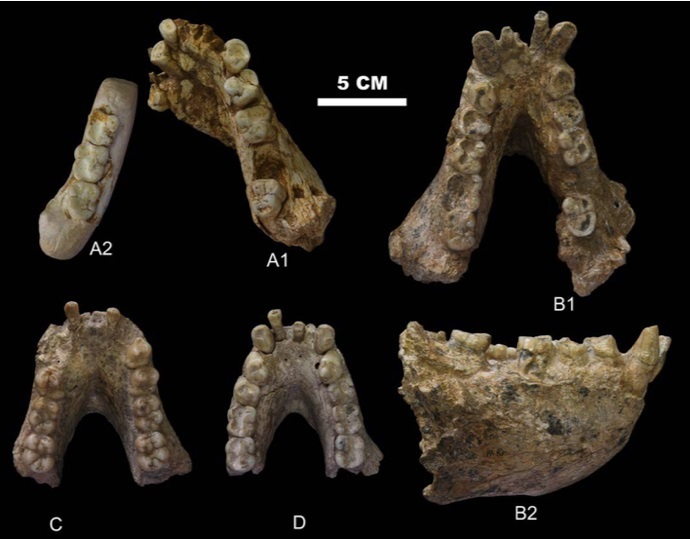
Частичные нижние челюсти гигантопитека (Gigantopithecus blacki) из Китая: A1 — челюсть с правыми р3-m1, m3 и левыми c-p4 из пещеры Яньлянь; A2 — фрагмент левого тела челюсти с m1-m3 той же особи, что и A1; B — челюсть III из пещеры Сяоянь; C — челюсть II из пещеры Сяоянь; D — челюсть I из пещеры Сяоянь.
Строение и функции
Поскольку имеющийся в наличии ископаемый материал довольно скуден (почти исключительно отдельные зубы и нижние челюсти без восходящих ветвей), реконструкция внешнего облика гигантопитеков является весьма нелегкой задачей. Конечно же, прежде всего бросаются в глаза значительные размеры сохранившихся остатков. Однако точно определить по ним величину их обладателей далеко не просто. К примеру, известно, что у австралопитеков (Australopithecus) коренные зубы достигали значительной величины, но чересчур высоким ростом они не отличались — не более 1,7 м в высоту. Таким образом, при реконструкции необходимо учитывать, что рост гоминоидов нередко мало коррелирует с размерами их зубов. Поэтому после первых находок зубов исследователи благоразумно предположили, что несмотря на очень массивные челюсти гигантопитеки были не крупнее современных горилл (Gorilla).
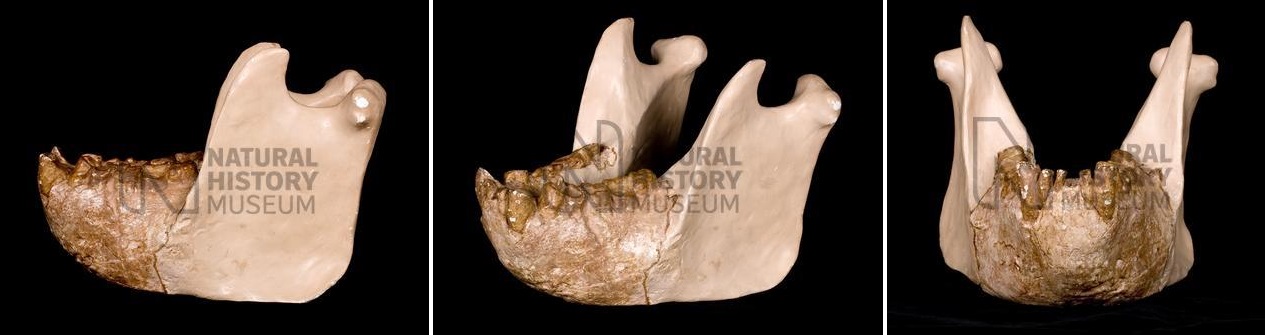
Реконструированная нижняя челюсть гигантопитека (Gigantopithecus blacki) из пещеры Сяоянь, Китай.
Когда были обнаружены челюсти гигантопитеков, ситуация несколько прояснилась. Было предложено множество расчетов предполагаемых габаритов этих обезьян. Очевидно, что использование величины зубов в качестве показателя массы тела у ископаемых приматов проблематично, поскольку размер зубов значительно варьирует в пределах любого размерного класса в результате воздействия множества функциональных и филогенетических факторов. Тем не менее, из того, что известно об их анатомии, можно сделать разумные доводы в пользу того, что гигантопитеки, вероятно, являлись относительно мегадонтными гоминоидами, и что любой прогноз массы тела, основанный на размере коренных зубов, вероятно, будет завышенным.
Согласно одной из умеренных оценок, исходя из размеров самой крупной нижней челюсти Gigantopithecus blacki (высота горизонтальной ветви — 184 мм, общая ширина — 104 мм, высота в области симфиза — 67 мм), при сходных с гориллой пропорциях рост ее обладателя должен был достигать 2,3 м, а вес составлял примерно 280 кг (для сравнения: взрослый самец гориллы имеет рост около 1,7 м и весит порядка 170 кг). Что касается расчетной высоты черепа самца Gigantopithecus blacki, то она оценивается в 45 см от края нижней челюсти до наиболее высокой точки сагиттального гребня (у самца гориллы — соответственно 25 см).
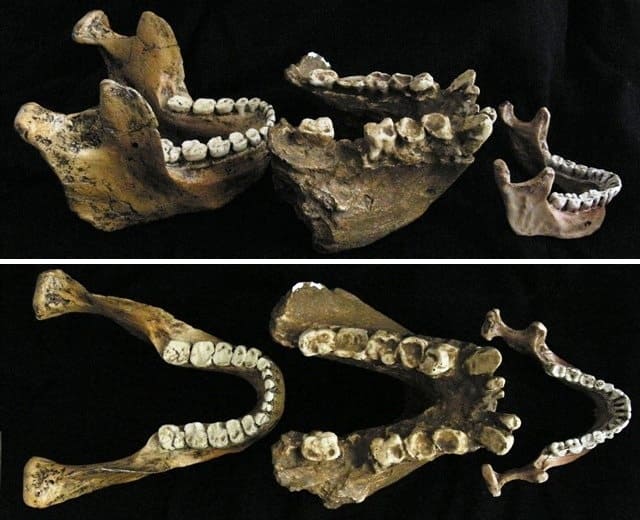
Относительные размеры нижних челюстей небольшой церкопитекоидной обезьяны, человека разумного (Homo sapiens), неандертальца (Homo neanderthalensis) и самца гигантопитека (Gigantopithecus blacki).
Разумеется, пока не будут найдены другие кости, всего этого с уверенностью нельзя ни утверждать, ни отрицать. Относительные размеры челюсти и зубов гигантопитеков и горилл представляются довольно надежным мерилом, однако известно немало случаев, когда подобного рода расчеты оказались далекими от истины. Поэтому стоит ограничиться утверждением, что Gigantopithecus blacki был крупнее гориллы и является самым большим известным на сегодняшний день гоминоидом и приматом в целом. Необходимо также отметить, что, судя по размерам найденных зубов, позднемиоценовые гигантопитеки были гораздо меньше плейстоценовых (причем среднеплейстоценовые экземпляры несколько крупнее раннеплейстоценовых). Таким образом, у этих обезьян четко прослеживается прогрессивное увеличение размеров тела на протяжении всей их эволюционной истории.
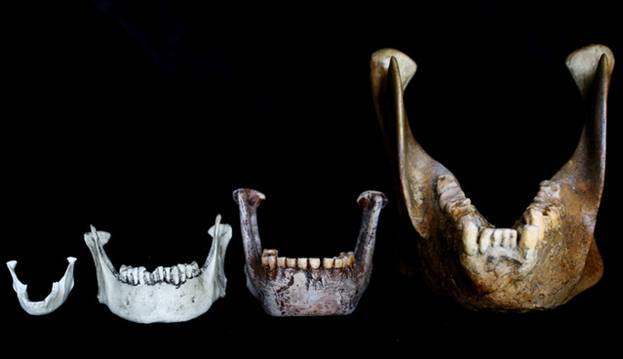
Относительные размеры нижних челюстей небольшой церкопитекоидной обезьяны, человека разумного (Homo sapiens), неандертальца (Homo neanderthalensis) и самца гигантопитека (Gigantopithecus blacki).
Нижняя челюсть гигантопитеков очень массивна. Для ее тела характерны относительная укороченность и большая высота, которая увеличивается кзади и достигает максимума на уровне m3. Наружная и внутренняя поверхности тела гладкие, отличаясь этим от человека и вполне сближаясь с человекообразными обезьянами. Лишь невысокий и гладкий валик проходит наискось вдоль наружной боковой поверхности, понижаясь вперед; наибольшей высоты он достигает по направлению к заднему коренному. Подобный валик, или гребень, часто встречается на нижних челюстях современных и ископаемых человекообразных обезьян. Ему соответствует валик, проходящий на внутренней стороне и направляющийся к симфизу. Его нижний край образует язычно-подъязычную линию. Сильное развитие зубной системы сопровождается низко расположенными на каждой стороне челюсти одним-двумя крупными и открывающимися вперед подбородочными отверстиями. Надостное отверстие также расположено низко.

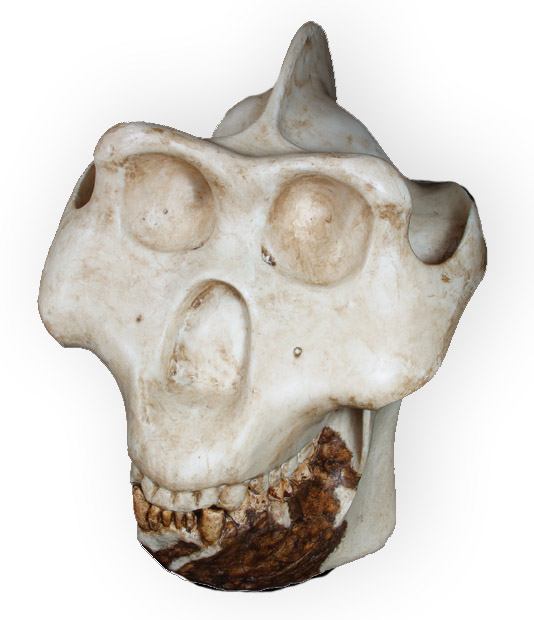
Реконструированный череп самца гигантопитека (Gigantopithecus blacki).
Симфиз сильно укреплен. Передняя поверхность симфиза обычно довольно крутая и наклонена примерно на 10 ° по вертикали. Ее профиль в боковой проекции направлен дугообразно вниз и назад, потом загибается в горизонтальном направлении, переходя в нижнюю сторону широкой базальной пластинки. С внутренней стороны область симфиза ясно разделяется на альвеолярную (верхнюю) и базальную (нижнюю) части. Первая на большем протяжении гладкая, будучи снизу ограничена заметным валиком, составляющим продолжение язычно-подъязычной линии. Это — верхний поперечный валик, развитый у крупных человекообразных обезьян. Под ним расположена глубокая ямка — место прикрепления подбородочно-язычной и подбородочно-подъязычной мышц (у человека для этого служат подбородочные ости). Дно этой ямки образовано верхней стенкой базальной пластинки ("обезьяньей полки"). Эта пластинка у гигантопитеков достигает больших размеров. В то время как у шимпанзе ее задний край доходит до уровня клыка, у горилл — до p3, у гигантопитеков она заходит значительно дальше назад — до уровня m1 или m2. Нижний край симфизной части сильно утолщен с образованием выдающегося поперечного валика на ее внутренней поверхности. В целом толщина симфиза у гигантопитеков превосходит все, что наблюдается у родственных форм.
Зубные ряды гигантопитеков хорошо известны по большой выборке разрозненных зубов. Частичные нижние челюсти также предоставляют полезную информацию о пропорциях нижнего зубного ряда. В высшей степени узкоспециализированная зубная система с уникальной комбинацией морфологических признаков четко отличает гигантопитеков от всех других ископаемых и современных гоминоидов.
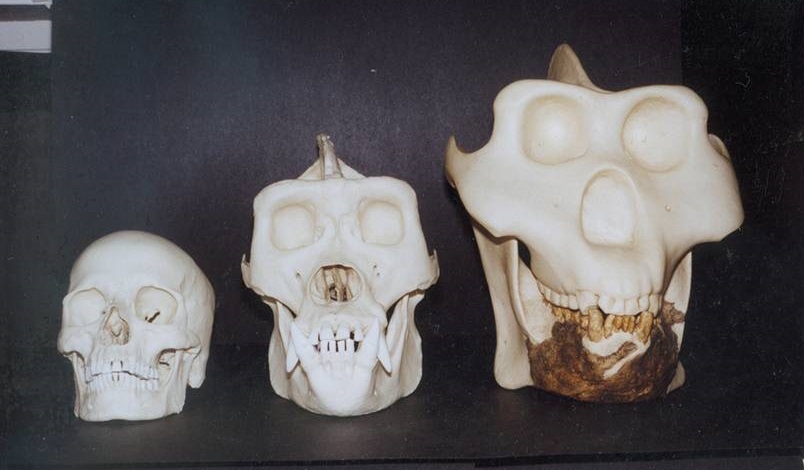
Относительные размеры черепов человека разумного (Homo sapiens), гориллы (Gorilla gorilla) и гигантопитека (Gigantopithecus blacki).
Нижний зубной ряд относительно короткий и слабо U-образный, его половины расходятся назад под углом около 25 °. Индекс длины к ширине альвеолярной дуги близок к таковому у орангутанов (Pongo). Диастемы на нижней челюсти отсутствуют или очень маленькие. Большая часть сохранившихся зубов частично стерта, и характер стертости позволяет установить тип соприкосновения нижних зубов с верхними. Износ нижних резцов сначала происходит на их апикальном крае, а затем часто захватывает язычную поверхность коронки. При сильном износе истертость распространяется по язычной поверхности далеко вниз и даже достигает корней. Это позволяет предполагать, что режущий край верхних резцов выступал над нижними резцами несколько вперед, т. е. прикус имел форму, чаще всего встречающуюся у современного человека. Верхний клык соприкасается с латеральной стороной нижнего клыка и с проксимальной р3. На этих местах заметна стертая поверхность скольжения. Износ нижнего клыка обычно изначально сосредоточен на вершине, но на более поздних стадиях распространяется вниз по дистальной поверхности к основанию коронки. На верхних коренных зубах язычные бугорки стирались сильнее, чем щечные, а на нижних — наоборот, что с течением времени приводило к появлению больших участков обнаженного дентина на соответствующих сторонах жевательных поверхностей. В целом верхние зубы несколько наклонены наружу, а нижние — внутрь, и верхняя зубная дуга несколько шире нижней. Такое соотношение характерно и для современных крупных человекообразных обезьян.
Резцы небольшие и плотно посаженные, сходные с таковыми орангутанов, но без морщинистости. Верхние резцы относительно массивнее нижних. Внутренние верхние резцы много больше наружных; они толсты и на язычнной стороне имеют хорошо развитый бугорок, растущий по направлению к середине их лопатообразной поверхности; щечная сторона несколько выпукла. Корень конический. Наружные верхние резцы достигают почти такой же высоты коронки, но много уже внутренних и более развиты в щечно-язычном направлении, чем в мезио-дистальном (передне-заднем). Язычный бугорок выступает в форме валика. Наружные верхние резцы характеризуются большей изменчивостью, чем внутренние, по размерам и некоторым чертам строения. Нижние резцы, как у многих высших приматов, — самые мелкие из зубов гигантопитеков. Они расположены вертикально, обладают более низкими коронками и очень длинными корнями. Внутренние нижние резцы при этом меньше наружных (также они относительно намного меньше, чем у современных человекообразных обезьян и миоценовых азиатских гоминоидов). Кроме размеров, внутренние и наружные резцы различаются еще толщиной корня, который, будучи сжат в мезио-дистальном направлении, очень утолщен в язычно-щечном. Размер и форма корней наружных нижних резцов мало отличаются от корней резцов внутренних.

Относительные размеры черепов человека разумного (Homo sapiens), гориллы (Gorilla gorilla) и гигантопитека (Gigantopithecus blacki).
Клыки слабо выступают над уровнем соседних зубов, тоньше, чем предкоренные, конические, с тупой вершиной. Коронки клыков относительно невелики, но при этом довольно мощные, широкие и плоские. Корни очень длинные — по меньшей мере, вдвое длиннее коронок. Один из крупных верхних клыков достигает во всю длину 59,7 мм, из которых на долю корня приходится 40,4 мм. Щечная поверхность верхних клыков заметно выпукла, язычная — несколько вогнута; с дистальной стороны коронка острая, с мезиальной — тупая. У основания коронки заметен цингулюм. Нижние клыки, будучи очень массивными, отличаются от верхних несколько меньшими диаметрами — мезио-дистальным и щечно-язычным; при этом, как правило, последний несколько превышает первый. Мезиальная и дистальная поверхности коронки клинообразны; на губной проходит продольный валик; язычная слегка выпукла. Поясок у основания коронки охватывает ее со всех сторон. Хотя на первый взгляд клыки гигантопитеков и напоминают человеческие, характер стертости их коронок свидетельствует о функциональной близости скорее к предкоренным и коренным, чем к резцам, как у людей. Клыки у самок не столь массивны, как у самцов; по своей форме у представителей различных полов они практически не отличаются. Между нижним клыком и р3 имеется небольшая диастема (у других человекообразных обезьян она, как правило, ярко выражена).
Щечные зубы Gigantopithecus blacki достигают максимальной среди приматов величины. Высота коронки левого m3, описанного фон Кенигсвальдом, равна 12,5 мм (самец гориллы — максимум 8,2 мм, среднее для современного человека — 6,6 мм). Длина коронки — 22,3 мм (горилла — 18 мм, человек — 10,7 мм), наибольшая ширина — 17 мм (горилла — 15,1 мм, человек — 10,8 мм). Площадь жевательной поверхности m3 составляет 363 кв. мм, тогда как у самца гориллы она достигает 312 кв. мм, у мегантропа — 220 кв. мм, у современного человека — 110 кв. мм. По объему коронки (4420 куб. мм) m3 гигантопитека превосходит почти вдвое коренной зуб крупнейшего самца гориллы (2356 куб. мм) и в шесть раз зуб современного человека (среднее 723 куб. мм). Коронки щечных зубов высокие, широкие и относительно плоские, с толстой эмалью и добавочными бугорками. Их корни намного длиннее, чем у современных человекообразных обезьян, соответствующие высокому телу нижней челюсти. Изменения подобного рода в строении щечных зубов у древних гоминид (например, у австралопитеков) свидетельствуют об адаптации к питанию растительной пищей.
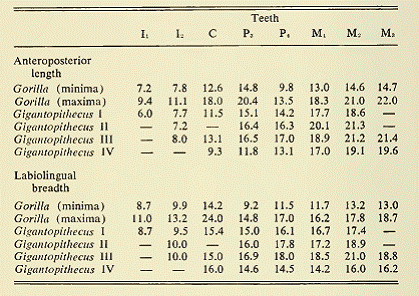
Промеры зубов Gigantopithecus blacki (I, II, III) и Gigantopithecus giganteus (IV) в сравнении с Gorilla.
Верхние предкоренные обладают некоторыми общими чертами: контуры их многоугольны (или трапециевидны — для Р3); они несут по два бугорка, из которых щечной паракон больше и выше язычного протокона; снабжены крупным и массивным корнем, дающим два корешка в щечном и один в язычном направлении. Оба бугорка Р3 отделены бороздкой, проксимальная часть жевательной поверхности короче дистальной и более округлена по краю. Коронка P3 немного больше, чем P4. Коронка Р4 скорее эллиптической формы, разделяющая бугорки бороздка проходит по середине жевательной поверхности и делит ее на две почти равные половины. Цингулюм умеренно развит на Р3 и слабо развит или отсутствует на Р4.
Нижние предкоренные различны. У р3 коронка сжата и удлинена, вытянутая сильнее не в мезиальном, а в щечном направлении. На жевательной поверхности имеется два бугорка, из которых щечной (талонид) хорошо развит, язычный (тригонид) же сильно ему уступает, присутствует небольшой мезиальный фланец. Со всех сторон коронку окружает базальный цингулюм, более выступающий с щечной поверхности. Задняя половина коронки больше передней и отделяется от нее бороздкой. Обе ямочки — передняя и задняя — выражены. Зуб имеет двураздельный корень. р4 испытывает моляризацию. Его коронка сжата в мезио-дистальном направлении, так что ее ширина несколько превышает длину и жевательная поверхность имеет неправильную квадратную форму. Оба ее бугорка смещены в мезиальном направлении, так что явно выступает талонид, который ниже большого тригонида. Корень расчленен на переднюю и заднюю ветви.
Коренные зубы очень велики. M1 меньше, чем M2, но в остальном они очень сходны. Коронки верхних коренных настолько высоки, что по отношению к длине и к толщине они превосходят коронки прочих высших приматов и уступают только человеческим зубам. В то время как щечная поверхность коронок направлена вертикально, их язычная сторона отклоняется под углом в 65 ° по направлению наружу. Проксимальная же и дистальная поверхности лежат в вертикальных плоскостях. Жевательная поверхность квадратно-ромбической формы и несет четыре хорошо развитых, но низких бугорка. Бугорки имеют характер "глыбок", т. е. у них округленные очертания, тесное расположение и разделены они очень узкими и глубокими бороздками. Из четырех бугорков наибольшего развития достигает протокон. По краю коронки между параконом и протоконом лежит небольшой дополнительный бугорок-протоконуль. На щечной и язычной сторонах коронки проходят заметные перпендикулярные борозды, из которых щечная отделяет паракон от метакона, а язычная — протокон от гипокона. Наиболее мощный валик, хотя и пересеченный тонкой бороздкой, соединяет протокон с метаконом. Другой, более тонкий валик проходит от протокона к паракону. М3 отличается несколько меньшими размерами, чем М2. Особенному сокращению подвергся его талон, а гипокон меньше метакона.
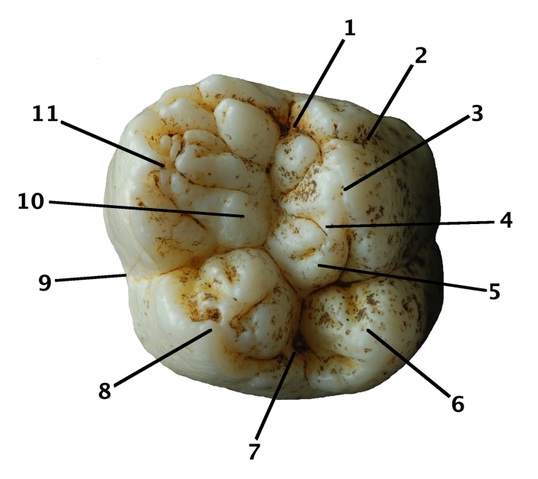
Жевательная поверхность правого M1 или m2 гигантопитека (Gigantopithecus blacki) из пещеры Саньхэ, Китай: 1 — мезиальная ямка; 2 — цингулюм; 3 — протокон; 4 — гипопротокриста; 5 — crista obliqua; 6 — гипокон; 7 — дистальная ямка; 8 — метакон; 9 — щечно-язычный перехват; 10 — бассейн тригона; 11 — паракон.
m1 меньше, чем m2, а в остальном они весьма сходны и в некоторых случаях очень трудноразличимы. Крупные коронки этих зубов несколько удлинены и с сильнее выраженным сужением в средней части, но не так высоки, как у верхних коренных. Бугорки имеют ту же форму "глыбок", что и на верхних коренных; щечная и язычная стороны коронки выпуклы, цингулюм развит слабо и более заметен на щечной стороне. Здесь почти вся высота коронки пересечена перпендикулярной бороздой, отделяющей протоконид от гипоконида. Подобная бороздка на язычной стороне отделяет метаконид от энтоконида. Кроме этих четырех бугорков, всегда развит мезоконид, положение которого не отличается постоянством; он может быть целиком смещен на щечную сторону, но может располагаться и центрально. Примерно в 25 % случаев имеется также VI-й бугорок; нередко, кроме того, присутствуют добавочные бугорки: внутренний добавочный между энто- и метаконидом. Самым крупным обычно является метаконид, превосходящий остальные бугорки в продольном и в поперечном направлении; он соприкасается с гипоконидом на значительном протяжении. Как последнее обстоятельство факт, так и постоянное присутствие мезоконида обусловливают хорошо выраженный дриопитековый узор нижних коренных. Бугорки тригонида, прото- и метаконид несколько выше остальных, да и сам тригонид в большинстве случаев несколько шире талонида. Гипоконулид на m1 и m2 выражен очень слабо. Эти зубы обладают хорошо выраженными корнями, мезиальным и дистальным; сужения в области шейки не заметно. m3 лишь в среднем несколько меньше m2, в отдельных случаях его размеры очень невелики. В отличие от М3 на m3 трудно указать какие-либо признаки редукции. Тригонид на нем слегка превышает по ширине талонид.
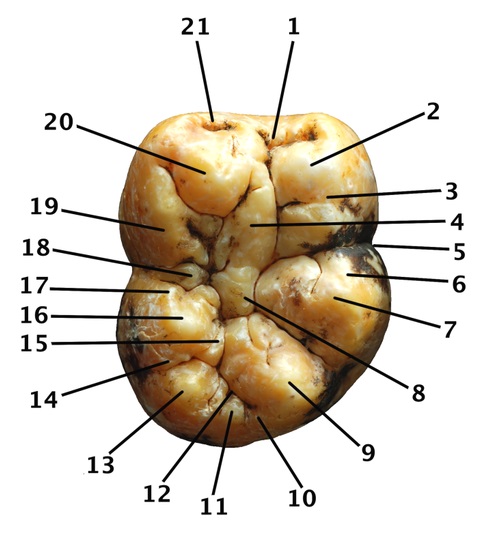
Жевательная поверхность правого m1 или m2 гигантопитека (Gigantopithecus blacki) из пещеры Бойю, Китай: 1 — мезиальная ямка; 2 — протоконид; 3 — постпротокристид; 4 — бугорок талонида; 5 — щечно-язычный перехват; 6 — прегипокристид; 7 — гипоконид; 8 — бассейн талонида; 9 — гипоконулид; 10 — дистальный краевой гребень; 11 — дистальный добавочный бугорок; 12 — дистальная ямка; 13 — tuberculum sextum; 14 — постентокристид; 15 — гипоэнтокристид; 16 — энтоконид; 17 — преэнтокристид; 18 — добавочный язычный бугорок; 19 — постметакристид; 20 — метаконид; 21 — мезиальный краевой гребень.
Зубная полость коренных расширяется к базальной части коронки и продолжается в сравнительно толстые и сближенные корни. Нижнечелюстной фрагмент из пещеры Яньлян демонстрирует доказательства потери правого m2 до прорезывания соседнего m3, растущего наклонно. Это позволяет предположить, что данная особь смогла выживать в течение длительного времени, несмотря на нарушение жевательной функции.
Зубная эмаль Gigantopithecus blacki в абсолютном выражении является самой мощной среди приматов, достигая на коренных зубах в среднем 2,5-2,9 мм, а максимально — более 6 мм. Однако, что касается относительной величины зубов, толщина эмали гигантопитеков перекрывается с таковой Homo и частично Pongo, но меньше, чем у Graecopithecus, Lufengpithecus и Australopithecus. При этом у Gigantopithecus blacki эмаль толще на стороне коронки, больше подверженной износу — т. е. на язычных буграх верхних коренных и щечных буграх нижних коренных зубов. Подобно орангутанам и, возможно, все понгинам, коренные зубы обладают относительно короткими дентиновыми рогами, выступающими в верхний слой эмали (у гоминин они относительно гораздо длиннее). Коренные зубы гигантопитеков являются наиболее гипсодонтными (с эмалью, оканчивающейся за пределами десен) среди всех обезьян.
Исследование m3 показало, что формирование бугорковой эмали было длительным (600-800 дней), периодичность образование штрихов Ретциуса составляла 11 дней, а формирование коронки длилось 4,09 года. Такие результаты указывают на относительно долгий период развития зубов по сравнению с современными человекообразными обезьянами. Секвенирование белков эмали Gigantopithecus blacki выявило наличие альфа-2-HS-гликопротеина, который у современных обезьян играет важную роль в минерализации костей и дентина. Поскольку он был обнаружен в эмали, а не в дентине, альфа-2-HS-гликопротеин мог являться у гигантопитеков дополнительным компонентом, способствовшим биоминерализации эмали во время ее роста. Подобно многим другим обезьянам, скорость образования эмали вблизи границы с дентином начиналась примерно с 4 мкм в день.
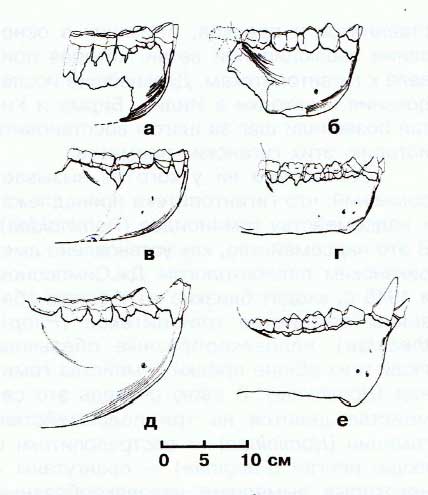
Прорисовки передних частей нижних челюстей Gigantopithecus blacki (а, б, д), Gigantopithecus giganteus (в), а также самки (г) и самца (е) Gorilla beringei.
Молочные зубы гигантопитеков сохранились в ничтожном числе. Можно лишь сказать, что они отличались меньшими размерами (хотя и среди них выделяются более крупные и более мелкие), а предкоренные — расходящимися корнями. Тригонид у этих зубов выше, но уже талонида, а морщинки на жевательной поверхности выражены яснее, чем на зубах постоянных.
Анализ нескольких сотен разрозненных зубов из ряда пещер позволил установить, что на протяжении раннего и среднего плейстоцена у Gigantopithecus blacki происходило постепенное, но в итоге весьма значительное увеличение размера зубов. Кроме того, возможно, наблюдается сдвиг в сторону усложнения жевательной поверхности щечных зубов — по крайней мере, в одной конечной популяции. Так, разрозненные зубы из пещеры Хэцзян на юго-востоке Китая, датируемые возрастом 400-320 тыс. лет (средний плейстоцен), демонстрируют ряд выразительных отличий от материала из раннеплейстоценовых местонахождений. На них имеются дополнительные гребни, исходящие от паракона и протокона ближе к средней линии (медиально), а основные гребни более острые. Вместе с тем, в отличие от верхних предкоренных других популяций, на зубах из пещеры Хэцзян отсутствует связь между препротокристой и парастилярным гребнем на P3 или с препаракристой на P4, в результате чего борозда прерывает мезиальный краевой гребень. Таким образом, относительно поздно существовавший хэцзянский гигантопитек может являться особой формой, приспосабливающейся к меняющимся пищевым ресурсам.
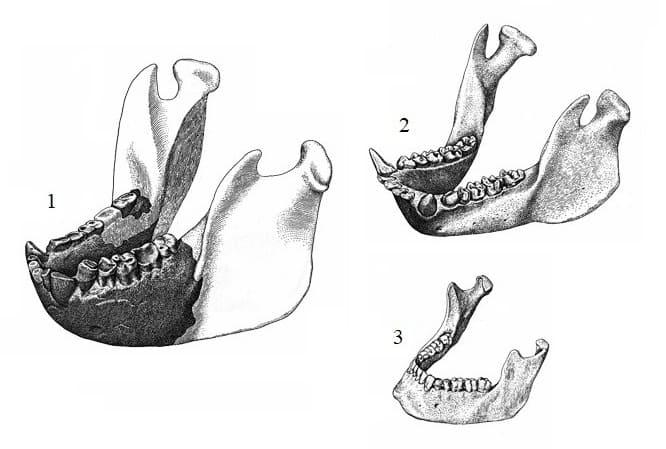
Прорисовки реконструированной крупнейшей нижней челюсти гигантопитека (Gigantopithecus blacki) (1) в сравнении с нижними челюстями гориллы (Gorilla gorilla) (2) и человека (Homo sapiens) (3).
У Gigantopithecus giganteus из позднего миоцена Сивалика зубы гораздо больше, чем у самого крупного вида Sivapithecus, а тело нижней челюсти высокое и крепкое. В то же время зубы и нижняя челюсть намного меньше, чем у Gigantopithecus blacki, что вполне ожидаемо для предшественника. Нижние резцы не сохранились, но пространство между клыками узкое, что подразумевает относительно небольшие и тесно расположенные резцы, как у Gigantopithecus blacki. Нижний клык относительно невелик по сравнению с заклыковым зубным рядом. Нижние щечные зубы устроены проще, чем у Gigantopithecus blacki, и больше напоминают таковые Sivapithecus. р3 однобугорчатый, с короткой передне-щечной площадкой для окклюзии с небольшим верхним клыком. р4 относительно большой, но мало моляризованный. Коренные имеют менее гипсодонтную коронку без выраженного щечно-язычного сужения и сложной жевательной поверхности, характерных для Gigantopithecus blacki.
Форма челюстей и малый размер резцов свидетельствуют о том, что гигантопитеки не могли отщипывать и отрывать кусочки пищи передними зубами, что характерно для современных человекообразных обезьян. Огромная высота нижней челюсти и выступающий вперед передний край восходящей ветви многократно увеличивают силу раздавливания пищи. Массивные симфиз и нижнечелюстные ветви свидетельствуют о способности гигантопитеков к мощному сжатию челюстей. Кроме того, задняя часть горизонтальной ветви нижней челюсти немного отклонена наружу, что, по всей вероятности, еще больше увеличивало силу сжимания челюстей. Особенности нижнечелюстного строения и характер износа зубов подразумевают жевательные движения челюсти из стороны в сторону (боковой экскурс). Без сомненья, сверхмассивные челюсти гигантопитеков поддерживала чрезвычайно мощная мускулатура.
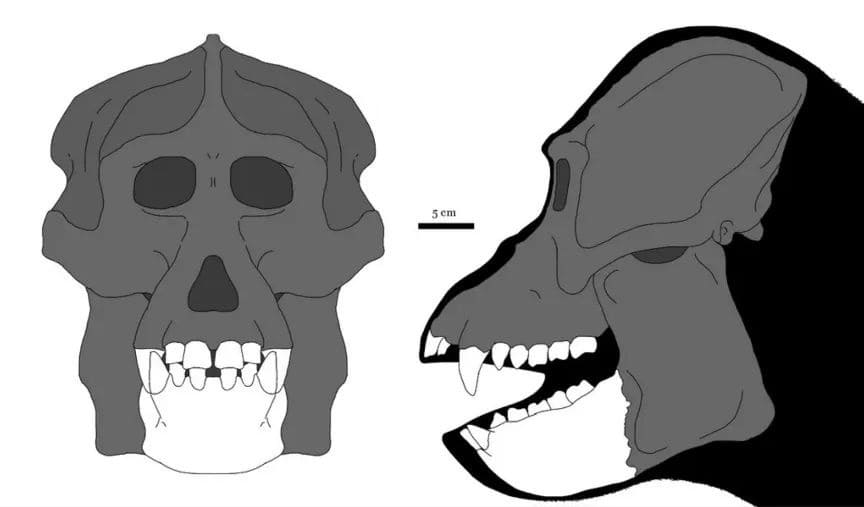
Прорисовки реконструированного черепа с нижней челюстью гигантопитека (Gigantopithecus blacki). Известный материал выделен белым, серая область восстановлена по краниальному скелету Sivapithecus, Pongo и Gorilla. Автор — М. Уиттон.
Для гигантопитеков, как и для современных человекообразных обезьян, характерен половой диморфизм. Предполагается, что самки были намного меньше самцов, уступая им в массе примерно вдвое. Исходя из этого допущения, из обнаруженных в гроте Сяоянь челюстей Gigantopithecus blacki одна крупная нижняя челюсть (II) с еще непрорезавшимся m3 и малостертыми бугорками, скорее всего, принадлежала молодому самцу 14-15 лет, наиболее крупная челюсть (III) — взрослому самцу, а самая мелкая челюсть (I) с утраченным m3 — взрослой самке. При этом высота тела нижней челюсти на уровне m1 взрослого самца почти на 40 % превосходит таковую взрослой самки. Подобно другим человекообразным обезьянам, наибольшие половые различия наблюдаются в высоте тела нижней челюсти на уровне симфиза и на уровне m2. Слабее выражены различия в некоторых толстотных размерах (например, толщина на уровне коренных). Из этого следует, что между самцами и самками существовала значительная разница в размерах черепа и, предположительно, массе тела. Половые различия в высоте тела нижней челюсти у гигантопитеков проявлены значительно сильнее, чем у современных человекообразных обезьян (так, у самцов горилл данный показатель в среднем лишь на 16-20 % превышает таковой у самок). Возможно, что это связано с их более крупными размерами.
Найденные отдельно зубы гигантопитеков также достаточно отчетливо распадаются на 2 размерные категории: крупные и мелкие. Поскольку основные морфологические особенности тех и других идентичны, можно полагать, что в различии размеров сказывается половая дифференцировка гигантопитеков. Близкую аналогию этому дают крупные человекообразные обезьяны — гориллы и орангутаны. Так, высота коронки верхних клыков, принадлежащих к крупному типу, колеблется между 21 мм и 29 мм; длина — от 16,8 до 21,2 мм; толщина, которая обычно (но не всегда) превышает длину, варьирует в пределах от 18,8 до 23,9 мм. У зубов мелкого типа длина коронки бывает от 14,1 до 17,7 мм, а толщина — от 13 до 17,4 мм. Среди современных человекообразных обезьян более выраженный диморфизм по размеру клыков наблюдается лишь у горилл, а по относительной величине нижней челюсти у разных полов гигантопитекам равных нет.

Возможные размеры тела самца Gigantopithecus blacki в сравнении с крупными самцами Pongo pygmaeus, Gorilla gorilla, Pan troglodytes и высоким мужчиной Homo sapiens. Автор — Р. Учитель.
Как и у современных человекообразных обезьян, передние конечности гигантопитеков явно были гораздо длиннее задних. Предполагаемый интермембральный индекс составляет 108, однако необходимо учитывать, что вычислен он полностью умозрительно. Точно так же пока ничем не подтверждено предполагаемое соотношение высоты черепа к общей длине тела гигантопитеков как 1 к 6,5. Отчетливее судить о телесных пропорциях этих приматов представляется возможным лишь после обнаружения их более полных ископаемых остатков.