Функции
Пищевое поведение саблезубых хищников до сих пор является одним из самых интригующих вопросов палеозоологии. Ввиду остутствия современных плотоядных с такой адаптацией лишь понимание функциональной анатомии позволяет построить вероятные гипотезы о методах использования удлиненных верхних клыков. Основополагающим для понимания способа нападения саблезубых хищников признается строение сосцевидной области черепа.

Прорисовка черепа с нижней челюстью и портретная реконструкция гомотерия (Homotherium crenatidens). Автор — М. Антон.
Сосцевидная область махайродонтов и "обычных" кошек несколько отличается. Сосцевидный отросток у Smilodon и Homotherium более развит; затылочный выступ у некоторых махайродонтов относительно небольшой, в результате чего кончик сосцевидного отростка направлен вниз в большей степени, чем затылочный выступ. Сосцевидный отросток отличается большей бугристостью, чем у современных кошачьих. В большинстве случаев кончик сосцевидного отростка пропорционально более широкий, чем у нынешних кошек. Сосцевидный гребень обычно более развит, с большим боковым расширением, в результате чего поверхность непосредственно сосцевидного отростка демонстрирует тенденцию располагаться далее книзу, кзади и не столь выдвинута в сторону, как у современных кошачьих.
Была предложена гипотеза, согласно которой саблезубы использовали сгибание шеи и сокращение мускулатуры, чтобы нанести нисходящий удар, позволяющий верхним клыкам вонзиться в плоть. Нижняя челюсть фактически не играла никакой роли при таком способе нападения, и явная адаптация к широкому открытию пасти предназначалась только для отведения нижней челюсти назад. Данная схема подразумевала гипертрофию мышц, опускающих голову и шею и делающими возможное успешное нанесению удара. Как доказательство такой адаптации, упоминалось увеличение поперечных отростков шейных позвонков, служащих для прикрепления группы мышц, сгибающих шею, и увеличенный сосцевидный отросток, который, в свою очередь, имеет следы прикрепления группы грудино-ключично-сосцевидных мышц, опускающих голову вниз. Также было предположено, что инерция тела хищника, движущегося по направлению к добыче, дополнительно помогала ударному действию верхних клыков. Однако другие специалисты утверждали, что длинные верхние клыки саблезубых кошек слишком хрупки, чтобы выдерживать подобное воздействие.
Большинство проблем этой теории было решено в т. н. гипотезе "рубящего удара", согласно которой саблезубые кошки умервщляли жертву посредством другого типа укуса. В этом сценарии поднимающие нижнюю челюсть мышцы играют поддерживающую роль, а главную силу для удара верхними клыками обеспечивало опускающее голову действие атланто-сосцевидной мускулатуры. В процессе рубящего удара нижняя челюсть противопоставляется действию верхних клыков, и анатомические особенности широкого открытия пасти рассматриваются как адаптация черепа и нижней челюсти к такому типу укуса. Кроме того, было показано, что строение шеи саблезубов отражает приспособление к мощным движениям шейных мускулов, включая дополнительное боковое вращение, что кажется необходимым условием при рубящем ударе.
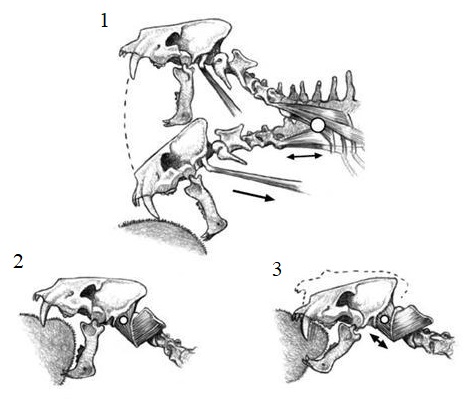
Схематическое изображение черепа и шейных позвонков Homotherium при нанесении гипотетического ударного (1) и рубящего (2, 3) укуса. В первом случае главное ударное движение происходит вокруг шейно-грудной области (большой белый круг), тогда как во втором основное вращение происходит в атланто-затылочном суставе (маленький белый круг). В первом способе тяга брахиоцефальных мышц (одна стрелка) и вес (двойная стрелка) обеспечивает основную силу удара. Во втором тяга атланто-сосцевидной мышцы (короткая двунаправленная стрелка) является наиболее важной силой для проникновения верхних клыков.
Основное различие между двумя гипотезами состоит в том, что первая подчеркивает важность наличия массивных грудино-шейных и задних шейных межпозвонковых суставов, тогда как вторая предполагает большую важность во вращении головы в атланто-затылочном суставе. Гипотеза "рубящего удара" основывалась на анатомии большой панды (Ailuropoda melanoleuca). Такой выбор был сделан в связи с общими представлениями об аналогичном расположении мышц, однако вызывал неуверенность в связи с их функциональным соответствием. В итоге сложилась неудовлетворительная ситуация, которая сохраняется по сей день.
Судя по всему, Homotherium наносил укус сверху вниз в соответствие с осью верхнего клыка. В результате появлялась открытая колото-резаная рана, равная по ширине этому зубу. Когда клык выдергивался из плоти, он своими зазубренными краями увеличивал рану еще больше. Исследование показало, что Homotherium обладал укусом, приспособленным для зажима и удержания мягких тканей при нанесении повреждений клыками. Функционально он походил не на удар верхними клуками Smilodon, а на укус Panthera leo из-за большего количества трабекулярной костной ткани, входящей в состав черепа этого рода. Различие в способах нанесения укуса свидетельствует о различных экологических адаптациях указанных махайродонтов.
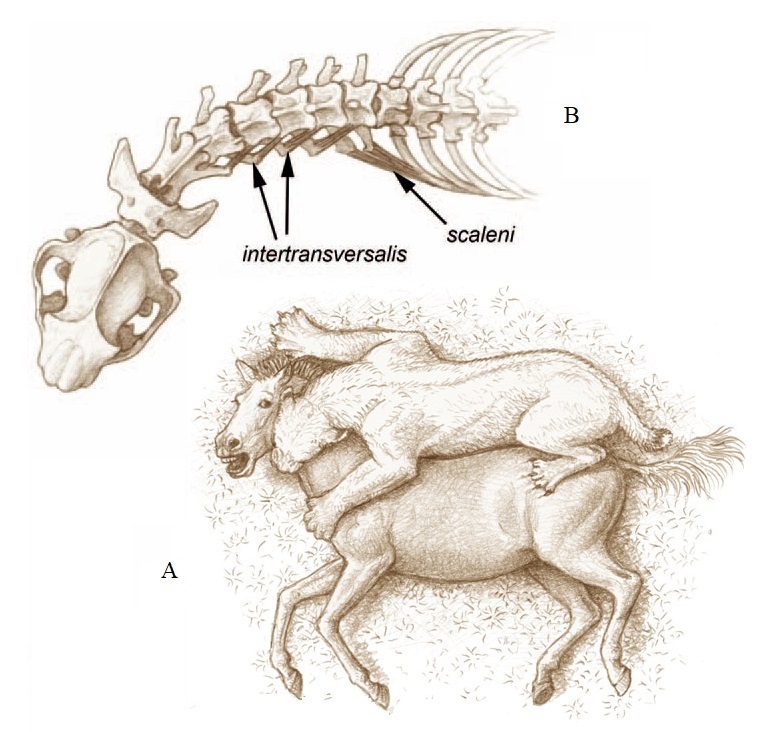
Реконструкция гомотерия (Homotherium latidens), применяющего укус клыками, сдвигаясь к горлу лошади (Equus sp.), одновременно прижимая ее к земле: А — вид сверху на жертву и хищника, демонстрирующий угол между центральной осью ее тела и головой; B — рисунок черепа и передней части позвоночника, иллюстрирующий боковое сгибание шеи и траектории волокон некоторых мышц, отвечающие за это сгибание. Автор — М. Антон.
Помимо верхних клыков, немаловажная роль в зубном аппарате принадлежала резцам. Все они, как верхние, так и нижние, а также резцоподобные нижние клыки, составляют как бы единый мощный выдвинутый вперед аппарат, состоящий из отдельных конусообразных, незначительно загнутых назад и плотно смыкающихся зубов. Судя по всему, этот аппарат усиливал способность челюстей захватывать и цепко удерживать добычу, а при надобности отрывать от нее куски. Сильно развитый сагиттальный гребень указывает на значительный объем височной мускулатуры большой и, следовательно, на большую силу укуса. Затылочная часть черепа скошена — предполагается, что данная особенность увеличивает вектор приложения сил, прилагаемых к передним зубам. Подобные тенденции будут вполне понятны, если учесть, что саблезубы в основном охотились на крупную дичь (им нередко приходилось, вероятно, удерживать в зубах и перетаскивать жертву в 2-3 раза крупнее самого хищника). Впрочем, согласно биомеханическим исследованиям, сила сжатия челюстей Homotherium была относительно небольшой, примерно в 2-3 раза уступая таковой соразмерных представителей рода Panthera и находясь на уровне гораздо более мелкой Puma.

Средний плейстоцен Европы. Гомотерии (Homotherium latidens) убивают лошадь (Equus sp.). Автор — М. Антон.
Относительно длинная шея Homotherium увеличивала мобильность головы во время нанесения смертельного укуса в горло жертвы, зафиксированной лапами и туловищем хищника. Homotherium имеет короткий поясничный отдел с выступающими в стороны поперечными отростками, отклоняясь, таким образом, от модели примитивных кошачьих, наблюдаемой у таких ранних махайродонтов как Nimravides и у всех ныне живущих фелин, у которых поясничный отдел длинный, а поперечные отростки более отчетливо вытягиваются в переднее-нижнем направлении. Похожий сдвиг наблюдается у медвежьих и гиеновых, и, конечно, в более выраженной форме — у копытных, делая спину этих животных более жесткой и сильной. Из этого следовала двойная выгода. Во-первых, короткая и жесткая спина действовала как пассивный передатчик движущей силы, генерируемой во время передвижения задними конечностями, экономя значительное количество энергии.
Во-вторых, у хищников, которые роют землю или переворачивают большие камни, негибкая спина лучше стабилизирована во время таких действий. Примерами такой адаптации могут служить барсуки и медведи. Типичные кошачьи, наоборот, приобрели удлиненную и гибкую спину. Это удлиняет прыжок и увеличивает абсолютную скорость и способность к взрывному ускорению, но ведет к большим энергетическим затратам. Такие животные исключительно быстро достигают высокой максимальной скорости броска, но утрачивают выносливость. Для саблезубых важным преимуществом жесткого, негибкого позвоночника должна была стать дополнительная сила противостоять боковым нагрузкам, вызванных яростным сопротивлением жертвы, удерживаемой передними лапами находящегося в полувертикальном положении хищника. Некоторые адаптации задних конечностей, возможно, также связаны с этой фазой охоты.

Поздний плейстоцен Северной Америки. Охота гомотериев (Homotherium serum) на молодого мамонта Колумба (Mammuthus columbi). Автор — М. Антон.
Стройная плечевая кость Homotherium указывает на адаптацию к быстрому бегу. Приподнятость большого бугорка дает преимущество (с точки зрения биомеханики) в работе надостной мышце, в то время как мускулы, обеспечивающие захват и удержание жертвы, такие как дельтовидная и мышцы, разгибающие пальцы, развиты весьма умеренно (судя по местам прикрепления мускулатуры ― дельтовидной бугристости, надмыщелку). С другой стороны, скорее боковое (как у Panthera), чем центральное (как у Acynonyx) положение большого бугорка указывает на то, что совместное отводящее действие подостной, большой круглой и надостной мышц, по-прежнему было очень важным. Относительно глубокая ямка локтевого отростка указывает на обычно выпрямленное положение локтевого сустава (характерный признак животных, приспособленных к быстрому бегу, таких как Acinonyx jubatus и Panthera leo). Кроме того, вертикальная ориентация ямки больше походит на таковую у Acinonyx jubatus, чем у пантеровых кошек, также указывая на способность к быстрому бегу.
Сохранившиеся травмы подтверждают, что действие мышц передней конечности было важным для поимки добычи. Так, правая плечевая кость Homotherium crenatidens из Сенез демонстрирует патологию развития сходную с той, которая выявлена у Smilodon fatalis с Ранчо Ла-Бреа — дельтовидную бугристость, протянувшуюся в задне-боковом направлении. Это предполагает наличие повторяющихся перегрузок дельтовидной мышцы во время сгибания конечности, что должно было происходить, когда хищник пытался удержать лапой сопротивляющуюся жертву. Изредка встречаются патологические костные изменения в области суставов, о чем свидетельствует лопатка Homotherium latidens из Шенингена. Они могут объясняться травмами или остеоартритом. Заболевания суставов передней конечности, в особенности локтевого, довольно широко распространены среди домашних кошек и являются следствием интенсивного использования передних ног.
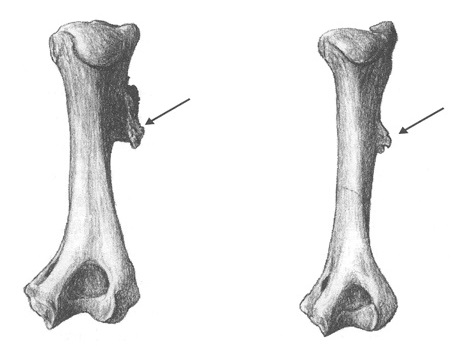
Прорисовки плечевых костей с костным разрастанием в месте прикрепления мышц смилодона (Smilodon fatalis) с Ранчо Ла-Бреа, США (слева), и гомотерия (Homotherium crenatidens) из Сенез, Франция (справа). Автор — М. Антон.
В пропорциях длин костей конечностей Homotherium демонстрирует наиболее разительные отличия от остальных кошачьих. Этот факт, вероятно, отражен как в значениях массы тела, так и в специфической функциональной конвергенции, спровоцированной подчеркнутой курсориальностью по сравнению с другими современными кошками. Частные значения длины плечевой кости к ее диаметру у родов Epicyon и Homotherium так же, как у современных гиеновых, находятся ниже 95 %-го интервала значений для псовых, указывая, что их плечевые кости были значительно более массивными, чем у последних. Фактически, плечевые кости Epicyon, Homotherium и современных гиен являются столь же массивными, как у большинства кошек, но дельтовидные гребни Epicyon и гиеновых выгляделили значительно короче, чем у кошачьих. Epicyon, Homotherium и представители семейства гиеновых оценочно достигали максимального веса тела на 50-70 % больше чем псовые сравнимых линейных размеров.
Грацильность лучевой кости наиболее заметна в узком дистальном конце, что также является одним из элементов приспособления к быстрому бегу. Это вступает в противоречие с необходимостью хватать и удерживать крупную добычу. Морфологическое подобие, предполагающее сходство в функционировании, между короткой и квадратной гороховидной костью Homotherium и гороховидной костью Acinonyx контрастирует с длинной гороховидной костью Panthera. Это, в свою очередь, должно было быть связано с действием локтевого разгибателя запястья.
Морфология средних фаланг пальцев кисти с их мало выраженной ассиметрией предполагает, что когти у Homotherium втягивались хуже, чем у пантеровых кошек, и в этом отношении он занимал промежуточное положение между ними и Acinonyx jubatus. Однако, II-й палец, по-видимому, сохранял повышенную хватательную функцию, как это следует из увеличенного выступа дистальной головки средней фаланги и по наклону ее дистальной суставной поверхности. Значительная редукция в размере когтевых фаланг II-V-го пальцев предполагает, что находящиеся на них когти выполняли в большей степени локомоторную функцию, улучшая сцепление, как это имеет место у Acinonyx jubatus. Как и у последнего, основную хватательную функцию выполнял I-й палец. Тем не менее, загнутая форма и сдавленность с боков когтевых фаланг II-V-го пальцев Homotherium предполагают, что находящиеся на них когти все еще могли играть роль в удержании жертвы, несмотря на их небольшие размеры.

Когтевая фаланга I-го пальца кисти гомотерия (Homotherium crenatidens).
Таранная кость Homotherium из Меальяды в Португалии достаточно четко отличается от таковой фелин, поскольку является гораздо более низкой, массивной, с меньшим большеберцовым блоком и выступом ладьевидной кости, свидетельствуя о другой линии развития, чем у остальных кошачьих. Такая же особенность фиксируется у барбурофелиевого Sansanosmilus. Ископаемые остатки из Меальяды довольно хорошо изучены и установлено, что морфологически они аналогичны образцам Homotherium crenatidens из Сенез во Франции и Homotherium latidens из Хандсхейма в Австрии. Сравнение с Homotherium crenatidens привело к заключению, что развитый передне-задний большеберцовый блок образца из Сенез подразумевает больший диапазон движения нижних сегментов конечности. Таким образом, остатки Homotherium из Меальяды соответствуют более поздней стадии на пути к стопохождению, что и ожидается исходя из их более молодого геологического возраста. При их сравнении с таранной костью из Хандсхейма заметной разницы не обнаружено.
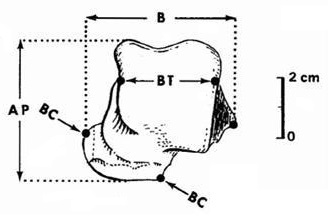
Прорисовка таранной кости гомотерия (Homotherium crenatidens): AP — передне-задний размер; B — ширина кости; BT — ширина большеберцового блока; BC — ширина выступов для ладьевидной кости.
У пантеровых кошек отношение между шириной таранной кости и шириной ее блока всегда выше даже с учетом низких значений у Pantera onca (конвергентно эволюционировавшем в направлении, аналогичном Homotherium) и Pantera spelaea (в данном случае, без сомнения, в связи с крупными размерами). Другими словами, у пантеровых кошек и большинства других фелин ширина таранной кости и блока меньше, а выступ для сочленения с ладьевидной костью расположен выше. Также наблюдаются различия в соотношениях ширины и высоты таранной кости. У пантеровых кошек эти значения больше (за исключением сравнения с ранними видами Homotherium), так как выступ ладьевидной кости играет большую роль в специализации таранной кости к передаче вертикального усилия, как у животных с явно выраженной тенденцией к стопохождению.
Укороченная пяточная кость и неглубокий блок таранной кости являются характерными чертами, которые у хищников обычно связывают со стопохождением. По этой причине Homotherium иногда так и реконструировался (т. е. с задней конечностью, опирающейся на стопу, а не на пальцы). Однако плюсневые кости расположены сравнительно плотно друг к другу, на что указывает верхняя выпуклость II-й плюсневой кости, вместо вывернутых наружу, как у медведей и других стопоходящих животных. Другие характерные особенности задних конечностей, включающие форму суставных поверхностей кубовидной и ладьевидной кости, указывают на то, что животное все-таки опиралось на пальцы.

Прорисовки реконструированных скелетов и реконструкции внешнего облика Homotherium с пальцеходящим (вверху) и стопоходящим (внизу) положением задних конечностей. Автор — М. Антон.
Относительно неглубокий блок таранной кости может являться адаптацией к борьбе с сопротивляющейся жертвой. Когда современные кошки имеют дело с крупными добычей, они стараются сохранять контакт своих задних лап с грунтом, часто опираясь на всю стопу, чтобы лучше контролировать ситуацию при яростных рывках добычи. У махайродонтов, которым необходимо было удерживать крупных жертв, сустав, соединяющий большую берцовую и таранную кости, вероятно, подвергался большим боковым нагрузкам, но стопа должна была оставаться на земле. Тогда это могло бы дать преимущество, обеспечивая некоторую дополнительную степень боковой мобильности и уменьшая риск потерять равновесие при отрыве стопы от земли.
Укороченные пяточная и плюсневые кости наряду со сниженным крестцом, вероятно, были связаны с необходимостью обеспечить большую устойчивость. В любом случае строение задней конечности говорит о силе, а не о высоких скоростных характеристиках. Задние лапы могли использоваться для мощного усилия, чтобы, к примеру, лишить добычу равновесия. Благодаря такой их способности крупные махайродонты, предположительно, были способны охотиться даже на молодых хоботных — мамонтов, лесных слонов и мастодонтов.
Таким образом, с функциональной точки зрения пропорции конечностей Homotherium сложны для понимания. Тенденции, наблюдаемые у Homotherium crenatidens из виллафранка Европы, доходят до крайности у Homotherium serum из позднеплейстоценовой пещеры Фризенхан в Северной Америке. Это наводит на мысль, что они давали реальное преимущество в борьбе за существование. Результаты изучения предполагают для Homotherium более курсориальный образ жизни, чем медленное скрадывание или засадная охота в лесистой среде обитания. Строение суставов и относительные пропорции конечностей указывают на довольно высокие способности к бегу по сравнению с другими махайродонтами. Об этом также может свидетельствовать уменьшение мышечной массы конечностей и снижение супинаторных возможностей, влияющих на способность к захвату добычи.

Прорисовка реконструированного скелета бегущего гомотерия (Homotherium crenatidens). Автор — М. Антон.
Удлиненная шея, длинные передние конечности, высоко расположенная лопатка и слегка скошенная спина с сокращенной поясничной областью придавали силуэту Homotherium определенное сходство с гиеной. Аналогия усиливалась укороченной пяточной костью, подразумевающей уменьшенную способность к прыжкам. В сочетании со стройной, явно пальцеходящей передней конечностью, бег Homotherium должен был выглядеть весьма необычно. В отношении локомоторных характеристик предполагается его промежуточное положение между пантеровыми кошками и гиенами. Однако Homotherium демонстрирует и очевидные анатомические отличия от гиены. Хотя он обладает такой же конструктивно мощной спиной, его конечности гораздо грацильнее, передняя конечность более удлиненная, а когти, скорее всего, относительно меньше. Такой экстерьер подразумевает адаптацию к умеренно быстрому преследованию добычи на длинной дистанции на манер гиен или собак.
Относительно короткая и жестккая спина Homotherium в меньшей степени, чем спина пантеровых кошек, способствовала стремительному ускорению, но, вероятно, обеспечивала большую выносливость во время бега (но все же значительно меньшую, чем у собак или гиен). Укороченные большая берцовая кость и стопа Homotherium подразумевают меньшие способности к прыжкам в сравнении с пантеровыми кошками, а удлиненные передние конечности, включая высоко расположенную лопатку, позволяли Homotherium бежать легким галопом. Этот тип бега на длинной дистанции энергетически являлся более эффективным, чем другие. Высказано предположение, что уменьшенные когти Homotherium были связаны со спринтерским бегом, как у Acinonyx, поскольку из современных кошек только он демонстрирует схожую степень редукции когтей.

Гомотерий (Homotherium crenatidens), бегущий галопом. Автор — М. Антон.
Однако, поскольку пропорции тела Homotherium не были хорошо приспособлены для спринта (особенно из-за укороченной спины и задних конечностей), то более вероятно, что редукция когтей являлась частью адаптации к улучшению сцепления с субстратом при беге на длинные дистанции со средней скоростью, как это в настоящее время происходит у псовых и гиеновых. Такие локомоторные способности указывают на предпочтение Homotherium открытых ландшафтов, а также позволяют предположить, что он должен был в поисках пропитания преодолевать значительные расстояния.
Еще одна примечательная особенность строения Homotherium и большинства других махайродонтов — короткий хвост. Эта часть тела используется современными хищниками для поддержания равновесия при прыжках или управления во время бега. Таким образом, предполагается, что саблезубые кошки не являлись хорошими бегунами.
Генетические исследования остатков с территории Юкон в Канаде подтверждают предположение, что Homotherium был хорошо приспособлен к образу жизни хищника-преследователя. В частности, у него выявлены вариации генов, которые повышали выносливость сердца и легких. Необычно большое квадратное носовое отверстие, сходное с таковым у Acynonix, предположительно способствовало быстрому насыщению крови кислородом, а также служило для охлаждения мозга во время интенсивного бега. Однако Homotherium наверняка не являлся столь быстрым бегуном, как Acinonyx, поскольку у него были совершенно иные пропорции тела. Судя по облегченному телосложению, слабым когтям и крупным хоанам, этот махайродонт, вероятно, был достаточно выносливым животным (особенно по меркам кошачьих) и был приспособлен к относительно длительному бегу и дальним переходам.

Портретная реконструкция гомотерия (Homotherium crenatidens). Автор — М. Антон.
Кроме того, у Homotherium обнаружены гены, повышающие зоркость глаз в светлое время суток и поддерживающие циркадный ритм. Его глазницы относительно крупнее, а участок коры головного мозга, отвечающий за обработку зрительной информации, больше и сложнее, чем у других махайродонтов, приближаясь к таковому у Acinonyx. Это подчеркивает способность хорошо видеть в светлое время суток, а не только ночью, как большинство кошек. Вместе с тем Homotherium сохранял превосходное ночное зрение, что подтверждается необходимостью охотиться полярной ночью в арктических регионах, где найдено много его остатков. Наконец, в геноме были выявлены мутации, связанные с развитием сложных форм социального поведения. Все вышесказанное, как считают палеогенетики, говорит о том, что Homotherium был дневным хищником, который специализировался на групповой охоте в угон на крупную добычу.