Происхождение и эволюция
В эволюции крупных кошек прослеживается ряд общих тенденций. Во-первых, существенное увеличение размеров тела и рост разнообразия в процессе развития. Второе: изменения в строении зубов, которые привели к выделению двух доминантных линий крупных кошачьих: пантеровых кошек с конической формой клыков и махайродонтов с сабельными верхними клыками. Последние со временем достигли существенных размеров, также раделившись на несколько основных линий.
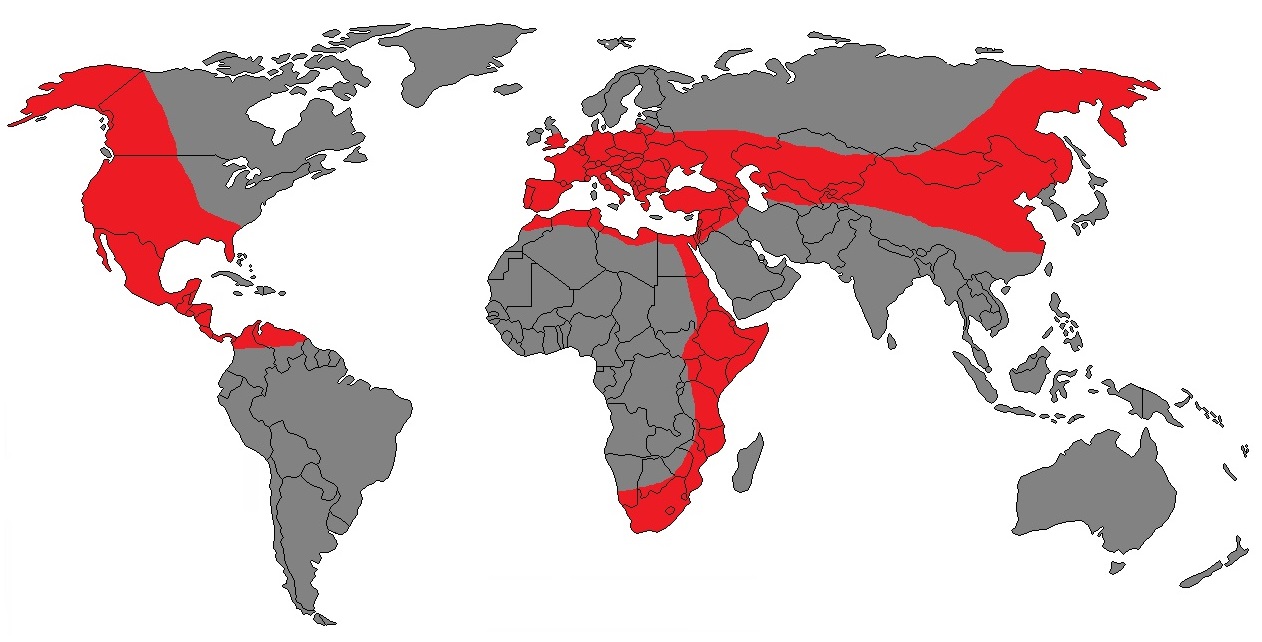
Область распространения ископаемых остатков Homotherium.
Род
Homotherium венчает собой одну из наиболее прогрессивных линий развития саблезубых кошек. Все они имели общую характерную особенность: мощное развитие передней части тела и относительная укороченность задней, что, как полагают, позволяло при нападении лишить добычу равновесия, повалить ее на землю и удерживать там до нанесения умерщвляющего укуса во избежание риска повердить длинные и относительно хрупкие верхние клыки. Помимо этого, для линии
Homotherium характерно удлинение и грацилизация передних конечностей, что вкупе с короткой и покатой спиной придавало этим животным особенно необычный для кошек экстерьер, несколько напоминающий облик гиен. Но ярче всего линию
Homotherium характеризует особая специализация зубо-челюстного аппарата.

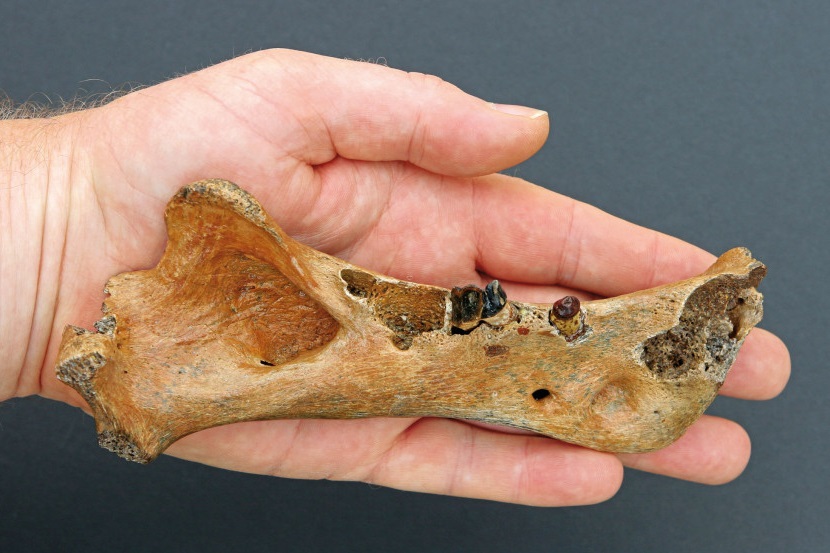
Часть правой ветви нижней челюсти гомотерия (
Homotherium sp.) (образец KNM-ER 931) из Кооби-Форы, Кения; наружная поверхность.
Уже первые представители рода Homotherium хорошо отличаются от таких своих миоценовых родственников, как Machairodus. По сравнению с последним, у Homotherium относительно узкий череп с менее расставленными скуловыми дугами, более крупные и выдающиеся вперед резцы, более изогнутые и сплющенные с боков верхние клыки, сильнее редуцированные предкоренные, более лезвиеподобные хищнические зубы с полностью утраченным язычным бугорком, суставной отросток для причленения нижней челюсти сориентирован более вентрально, более низкий венечный отросток, очень глубокий симфиз и высокое положение резцовой части нижней челюсти по отношению к альвеолярному краю щечных зубов. Кроме того, у Homotherium шея и передние конечности (в особенности предплечье) относительно длиннее, а поясница и хвост относительно короче, чем у Machairodus. Однако между указанными родами имеется и немало общего. Это, прежде всего, сходные пропорции зазубренных верхних клыков, выраженная редукция предкоренных зубов и большая длина нижней заклыковой диастемы. Таким образом, Homotherium демонстрирует своеобразное развитие морфологического комплекса, наблюдаемого у Machairodus.
Кости конечности гомотерия (Homotherium crenatidens) из Одесских катакомб, Украина.
Самые ранние ископаемые остатки, не определенные до вида и обозначенные просто Homotherium spp., найдены в отложениях позднемиоценового и раннеплиоценового возраста (около 7-4 млн. лет назад) на востоке, западе и юге Африки — в Эфиопии, Кении, Уганде, Сенегале, ЮАР. Впоследствии различные представители рода широко расселились в Африке, Европе, Азии, Европе и обеих Америках, просуществовав здесь на протяжении длительного времени. Реконструируемый ареал Homotherium — самый большой известный среди всех махайродонтов.
Нижняя челюсть гомотерия (Homotherium crenatidens) из Ливенцовки, Россия.
Из Африки описаны виды Homotherium ethiopicum, Homotherium africanum, Homotherium hadarensis и Homotherium problematicus, а также ряд форм, установленных лишь до рода. Их возраст охватывает временной промежуток от позднего миоцена до середины раннего плейстоцена (приблизительно от 7 до 1,5 млн. лет назад). Большинство находок имеет позднеплиоценовый и раннеплейстоценовый возраст. Homotherium hadarensis из позднего плиоцена Эфиопии некоторое время синонимизировался с Homotherium crenatidens, но в итоге все же был признан валидным. Родственные связи указанных видов не установлены. Таксономический статус других африканских таксонов ввиду фрагментарности материала остается неясным, однако понятно, что от евроазиатских форм они отличаются незначительно. В целом, гомотерии — одни из самых распространенных хищников в африканском плио-плейстоцене, хотя их окаменелости редко находят в достаточном количестве.
Череп с нижней челюстью гомотерия (Homotherium crenatidens) из Дманиси, Грузия.
В Европе рассматриваемый род впервые появляется в позднем плиоцене (примерно 3 млн. лет назад), будучи представлен видом Homotherium crenatidens. Древнейшие его остатки происходят из фаунистического комплекса Одесских катакомб на юге Украины, откуда также известен Megantereon sp. Раннеплейстоценовое присутствие Homotherium crenatidens отмечено в составе хапровской фауны местонахождения Ливенцовка в российском Приазовье, а также в целом ряде западноевропейских местонахождений виллафранкского возраста: Виллафранка-д'Асти, Оливола, Верхнее Вальдарно, Фарнета, Фоджа в Италии; Вильярроя в Испании; Лез-Этуар, Рокканейра, Рош-Ламберт, Сен-Валье, Купе, Сенезе во Франции; Нойленген в Германии; Грэнчану в Румынии; во многих из них обнаружен и Megantereon cultridens.
Череп Homotherium из Нихэваньской котловины, Китай.
На территории Азии Homotherium crenatidens известен из раннего плейстоцена Грузии (Дманиси, Ахалкалаки), Турции и Таджикистана (Куруксай). Из пинджорских отложений Сивалика в Пакистане, по возрасту соответствующих виллафранку Европы, происходит Homotherium palaeindicus (= Megantereon palaeindicus), соседствующий здесь с Megantereon falkoneri. Сравнение сиваликского гомотерия с другими евроазиатскими формами затруднено из-за фрагментарности материала, но скорее всего он является младшим синонимом Homotherium crenatidens. Остатки, принадлежащие последнему, также найдены в Женьцзыдуне на востоке и в Нихэвани на севере Китая. Фаунистические комплексы из этих местонахождения коррелируют с виллафранкскими фаунами Европы. На острове Ява из слоев, одновозрастных с поздневиллафранкскими, описан Homotherium zwierzyckii (= Epimachairodus zwierzyckii, Hemimachairodus zwierzyckii). Сегодня этот вид обычно синонимизируют с Homotherium crenatidens.

Череп с нижней челюстью гомотерия (Homotherium crenatidens) из Китая.
Таким образом в виллафранке крупный и стройный Homotherium практически на всем протяжении ареала сосуществовал с меньшим по размерам, коренастым Megantereon. Обе эти кошки к тому времени уже заметно отличались от своих миоценовых предков. Они имели удлиненную шею, сокращенную поясничный отдел позвоночника и редуцированный хвост. Однако тогда как Megantereon пропорционально стал очень массивным и обладал короткими конечностями приблизительно равной длины, длинные задние ноги Homotherium контрастировали с еще более длинными передними.
Череп с нижней челюстью гомотерия (Homotherium crenatidens).
В общем контексте евроазиатские гомотерии виллафранка представляют собой довольно однородную группу. Характерным их признаком является сильная редукция предкоренных зубов. Р3 и, особенно, р3 практически перестают функционировать, поэтому р3 на всех известных образцах почти не затронут стиранием, размеры и количество дополнительных бугорков на этом зубе сильно варьируют, часто он не имеет постоянного места в нижней челюсти, а иногда его альвеола вообще зарастает. Тем не менее, в большинстве случаев р3 — это маленький, однокорневой и одновершинный зуб, расположенный близко к р4 и не отделенный от него диастемой. По-видимому, у самых древних представителей рода следовало бы ожидать меньшей редукции предкоренных, чем у более поздних форм. Однако, как показал анализ материала, этот признак у гомотериев прослеживается в целом как у группы, хотя имеет достаточно высокую степень изменчивости. Так, наиболее ранний Homotherium crenatidens из Одессы характеризуется относительно малой редукцией р3. У Homotherium crenatidens из виллафранкских отложений Евразии степень редукции р3 гораздо выше, но даже среди поздневиллафранкских форм встречаются экземпляры с относительно мало редуцированным р3.
Череп с нижней челюстью гомотерия (Homotherium crenatidens).
В Евразии Homotherium сrenatidens был довольно широко распространен до конца раннего плейстоцена. Остатки этого возраста, определенные как Homotherium sp., происходят из Таджикистана (Лахути), Юго-Западной Сибири (Новосергеевский карьер) и Забайкалья (Кудун). В начале среднего плейстоцена Homotherium сrenatidens уступает место своему преемнику Homotherium latidens, а вымерших Megantereon cultridens и Megantereon falkoneri сменяют крупные представители рода Panthera — в Европе львинообразные, а в Восточной Азии тигрообразные формы.
В среднем плейстоцене евразийский ареал рода существенно смещается: он заметно сократился на юге (находки Homotherium latidens в Средиземноморье довольно скудны), но захватил Северо-Западную Сибирь, вплоть до бассейн рек Яны (Адыча) и Колымы. Нижние челюсти из Сибири обладают всеми морфологическими чертами, характерными для других поствиллафранкских форм Евразии, демонстрируя особенно большое сходство с находками из Чжоукоудяня в Китае. Установленное присутствие Homotherium в евразийской Берингии важно для понимания процессов плейстоценового фаунистического обмена между Евразией и Северной Америкой.
Череп гомотерия (Homotherium latidens) из Испании.
Представления о сроках вымирания данного рода в Европе со временем менялись. На протяжении большей части XX в. полагали, что остатки Homotherium latidens из пещеры Робина Гуда и Кентской пещеры указывают на присутствие этой кошки в позднем плейстоцене на территории Англии. Однако, новая интерпретация возраста пещерных отложений и геологический контекст, сопровождающий образцы, отодвинули их датировки до 0,5 млн. лет назад, т. е. до среднего плейстоцена. Относительно изолированного клыка из Кентской пещеры даже высказано предположение, что он был подобран верхнепалеолитическим человеком и использовался в качестве талисмана.
Были известны и несколько более поздние местонахождения Homotherium latidens — например, его остаткам из Артенака во Франциии и Атапуэрки в Испании примерно 0,4 млн. лет, а из Штейнгейма в Англии и Шенингена в Германии — около 0,3 млн. лет (примечательно, что во всех этих местонахождениях также выявлены следы присутствия Homo heidelbergensis, а обломок плечевой кости Homotherium latidens из Шенингена даже несет многочисленные царапины, что указывает на его использования в качестве орудия). На основании приведнных датировок считалось, что Homotherium исчез в Евразии до конца среднего плейстоцена. В связи с этим большой неожиданностью стала часть его нижней челюсти, в 2000 г. поднятая рыболовецкой сетью с шельфа южной части Северного моря вблизи побережья Нидерландов. С помощью радиоуглеродного анализа (С14) был установлен возраст находки — 28 тыс. лет, пик вюрмского оледенения. В то время данная область представляла собой низменную тундростепь, пронизанную реками.
Часть плечевой кости и зубы гомотерия (Homotherium latidens) из Шенингена, Германия.
Теоретически, кости, долгое время находившиеся в воде, могут насыщаться вторичным карбонатом, и в таком случае радиоуглеродная датировка будет омоложена. Однако если полученная датировка действительно верна, ей существует по меньшей мере два приемлемых объяснения. Первое подразумевает гораздо более длительное присутствие Homotherium в Евразии после его, казалось бы, очевидного исчезновения в среднем плейстоцене, вследствие чего на несколько сотен тысяч лет увеличивается период сосуществования Homotherium latidens и Panthera spelaea. Второе объяснение состоит в относительно позднем повторном заселении этого континента через Берингию из Северной Америки, где Homotherium непрерывно присутствовал на протяжении всего плейстоцена. Известно, что многие другие крупные млекопитающие в позднем плейстоцене распространились таким образом в Голарктике.
Хотя челюсть по региону обнаружения и была определена как принадлежащая Homotherium latidens, морфологически она неотличима от челюсти Homotherium serum из Северной Америки. Сравнение генетического материала, добытого из указанной челюсти и остатков 2 особей Homotherium serum возрастом около 50 тыс. лет с территории Юкон в Канаде показало практически полную его идентичность и принадлежность этих находок одному виду. Таким образом, подтверждается точка зрения, согласно которой Homotherium serum в позднем плейстоцене вселился в Старый Свет. С данной гипотезой вполне согласуется и обнаружение в 2020 г. на реке Бадьяриха в Якутии естественной мумии 3-недельного детеныша гомотерия, возраст которого оценивается приблизительно в 36 тыс. лет. Ясно, однако, что это животное оставалось в Старом Свете очень редким, тогда как Panthera spelaea был доминирующей крупной кошкой на большей части Европы и Северной Азии.
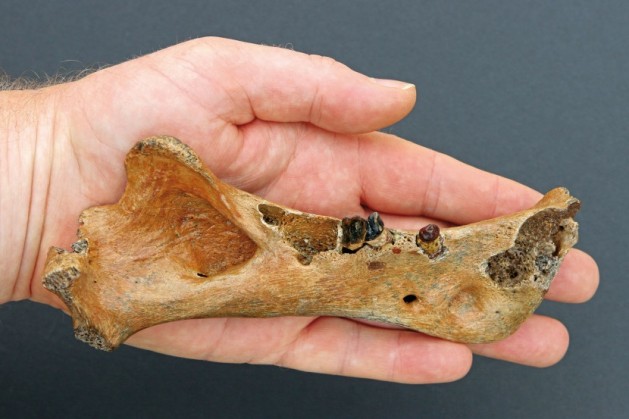
Правая нижнечелюстная ветвь гомотерия (Homotherium latidens) из шельфовых отложений Северного моря.
Анатомические различия между видами Homotherium обычно небольшие, поэтому разные исследователи принимают неодинаковое их число. Вдобавок высокая степень фрагментарности материала в сочетании с широким географическим распространением привела к интерпретации различных морфологических характеристик в качестве диагностических признаков самостоятельных таксонов и, в итоге, к описанию многочисленных видов, хотя валидность многих из них является сомнительной.
В настоящее время все плиоценовые и раннеплейстоценовые европейские виды Homotherium синонимизируют с Homotherium crenatidens, объясняя мелкие различия между ними половым диморфизмом и индивидуальной изменчивостью в пределах одного вида. Азиатские формы очень близки к европейским и также рассматриваются в рамках видов Homotherium crenatidens или Homotherium latidens. Анализ евроазиатского материала показывает, что изменчивость у Homotherium наблюдается в размерах черепа, верхнего клыка, величине верхней и нижней заклыковых диастем и степени развития предкоренных зубов. Эволюционные преобразования проявляются в упрощении структуры предкоренных и тенденции к их редукции.
Евроазиатский материал демонстрирует различия в длине верхнего клыка и залыковых диастем. Столь же варьирует степень редукции р3 и его положение в челюсти, а также наличие либо отсутствие диастемы между р3 и р4. В большинстве случаев эти изменения, по-видимому, подразумевают возрастно-половой диморфизм. Однако тенденция к упрощению структуры и редукции предкоренных зубов характеризует изменения на протяжении всей эволюции рода Homotherium: р3 значительно изменяется в размерах и по расположению, а р4 постепенно становится короче относительно m1. Это позволяет отличать русциний-ранневиллафранкские образцы от более поздних.
Поствиллафранкский Homotherium latidens разделяет с Homotherium crenatidens следующие морфологические особенности: крупные размеры; четко обозначенную зубную арку, образованную резцами и клыками; небольшую нижнечелюстную ямку; умеренно развитый подбородочный гребень, размещенный как раз перед р3; ясно отделенный передний бугорок р4. В качестве отличий Homotherium latidens можно отметить более изогнутые клыки, более редуцированный Р3 и нижнечелюстную ямку, расширенную до заднего края m1.
Череп с нижней челюстью гомотерия (Homotherium crenatidens).
В Европе самые полные находки ископаемых остатков Homotherium известны из середины виллафранка; западноевропейские формы раннего виллафранка в сравнительном анализе из-за недостатка данных используются редко. Более ранняя форма Homotherium crenatidens лучше представлена в восточноевропейской фауне Одесских катакомб на Украине. Ее отличительными чертами являются средние размеры тела, редуцированный p3 и m1, имеющий чуть большую длину, чем p4. Те же самые особенности наблюдаются у Homotherium из местонахождения Удунга в Забайкалье.
Согласно Фиккарелли (1979 г.), разделение между Homotherium crenatidens и Homotherium latidens основывается на меньшем размере и большем изгибе верхнего клыка последнего. Эти признаки первоначально использовались в диагнозе Homotherium latidens. Однако их высокая вариативность наблюдается у образцов из таких местонахождений Евразии, как Куруксай, Ливенцовка, Унтермасфельд, Семибалки-3 и ряд других. К примеру, верхний клык из Унтермасфельда сопоставим в размере с клыком крупнейшей виллафранкской формы Homotherium nestianus из Рокканейры во Франции, тогда как длина верхнего клыка Homotherium crenatidens из Куруксая равна экземпляру Homotherium latidens из Кентской пещеры в Англии. С другой стороны, проведенный анализ позволяет выделить различия межвидового уровня, применимые к виллафранкским и более поздним формам.
Путаница, возникшая в таксономии виллафранкских и среднее-позднеплейстоценовых форм возникла, несомненно, вследствие того, что в большинстве случаев остатки встречаются редко и зачастую являются фрагментарными. Также нельзя исключать возможный половой диморфизм либо случаи индивидуальной или географической изменчивости в пределах одного вида.
Череп гомотерия (Homotherium ischyrus) из Цитата-Каньон в штате Техас, США.
В Северной Америке род Homotherium просуществовал от позднего плиоцена или раннего плейстоцена (около 3-2,6 млн. лет назад) до самого его конца (примерно 12 тыс. лет назад). В более древний и примитивный вид Homotherium ischyrus были сведены бланкские таксоны Dinobastis ischyrus, Homotherium idahoensis, Homotherium johnstoni и Homotherium crusafonti. Стоит отметить, что эти виды известны лишь по нескольким неполным образцам, большинство из которых являются типовыми. Сегодня едва уловимые различия между ними объясняют внутривидовой изменчивостью. Например, Dinobastis ischyrus отличается от Homotherium johnstoni тем, что скорее имеет на р4 один задний бугорок, а не два. Недавно вид Homotherium ischyrus был переписан на основе большей части скелета из Берч-Крик в Айдахо, США. По общему строению это животное сходно с Homotherium crenatidens и Homotherium latidens из Евразии, но некоторые признаки — например, меньшее по размерам носовое отверстие, сохранение двукорневого р3 на нижней челюсти и удлиненного поясничного отдела позвоночника — предполагают более раннее его отделение от европейской линии. Кроме того, валидность Homotherium ischyrus может быть обоснована чисто биогеографическими соображениями.
Нижняя челюсть гомотерия (Homotherium ischyrus).
В ирвингтонское и ранчолабрейское время Североамериканский континент населял более продвинутый Homotherium serum (= Dinobastis serus). Здесь его ископаемые остатки встречаются от Аляски до Техаса. В южной части ареала этот вид сосуществовал (не обязательно в тех же местах или в аналогичных экологических условиях) со Smilodon fatalis, а на севере был единственным махайродонтом. К югу от границ висконсинского оледенения Homotherium serum не обнаружен в северо-центральных и северо-восточных Соединенных Штатах, что идентично ареалу Smilodon fatalis. Однако в сравнении с последним, остатки Homotherium serum находят довольно редко. Хотя морфологически и функционально эти две саблезубые кошки различны, не вполне ясно, почему они не представлены в палеонтологической летописи одинаково. На это обстоятельство вполне могли влиять местные особенности ландшафта (в частности, характер растительного покрова и наличие укрытий), тип и численность потенциальной добычи, а также наличие либо отсутствие социального образа жизни. Характерно, что остатки Homotherium serum на ранчо Ла-Бреа (всего в 2 местах в количестве 5 особей) столь же редки, как и окаменелости Panthera onca, тогда как остатки Smilodon fatalis найдены здесь по меньшей мере в 15 точках.

Череп с нижней челюстью гомотерия (Homotherium serum).
Отметим, что филогенетические связи между американскими видами
Homotherium, а также их связи с евроазиатскими видами окончательно не выяснены. Ряд ученых исключают ранне- и среднеплейстоценовых представителей рода
Dinobastis из состава
Homotherium (де Бонис, 1976 г.; Берта, 1987 г.; Баллезио, 1996 г.), хотя большинство синонимизируют оба рода (Куртен, 1962, 1968 гг.; Черчер, 1966 г.; Фиккарелли, 1979 г.; Андерсон, 1984 г.; Рон-Шатцингер, 1992 г.; Маккенна и Белл, 1997 г.). Некоторые авторы (Мартин с соавт., 1988 г.; Рон-Шатцингер, 1992 г.) синонимизируют бланкские формы с
Homotherium crenatidens, в то же время не дискутируя о таксономическом статусе более молодого
Homotherium serum. Рон-Шатцингер определенно считает это наименование действительным для всех ранчолабрейских образцов. Отмечено, однако, что особи из ирвингтония (например, найденные в Орегоне) отличаются значительной массивностью, будучи сопоставимы с
Homotherium crenatidens, тогда как в ранчолабреании они грацилизируются (экземпляры с ранчо Ла-Бреа). При этом более массивные образцы, чем позднеплейстоценовый
Homotherium serum, присутствуют в слоях озера Мэнникс с примерным возрастом 200 тыс. лет. Возможно, это указывает на наличие во второй половине плейстоцена Северно Америки двух разных форм
Homotherium.

Череп гомотерия (
Homotherium venezuelensis) (образец OR-1352) из Эль-Бреал-де-Орокуэл, Венесуэла.
Присутствие рассматриваемого рода в Южной Америке на сегодняшний день ограничивается находками из раннего плейстоцена (около 1,8-1 млн. лет назад) северо-восточной Венесуэлы (Эль-Бреал-де-Орокуэл) на самом севере континента — эта форма описана как Homotherium venezuelensis; вместе с ней найдены остатки Smilodon gracilis. Кроме того, близкая к Homotherium форма обнаружена в Уругвае, но ее возраст неизвестен, а родовое положение вызывает сомнения из-за неполноты и большой фрагментарности (беззубая часть ветви нижней челюсти). Незначительность остатков представителей трибы Homotherini в Южной Америке может обусловливаться существующей на этом континенте биогеографической дифференциацией. С другой стороны, тот факт, что обнаружено только несколько образцов в Венесуэле и один образец в Уругвае, может быть связано с низкой плотностью популяции этих саблезубых кошек. Такое объяснение отчасти согласуется с редкой, по сравнению с другими кошачьими (например, Megantereon, Smilodon и Panthera), частотой находок Homotherium на других континентах. Как долго он просуществовал в Южной Америке, не ясно.
вперед