Строение и функции
Размеры горгоноповых варьируют от мелких до очень больших, преимущественно средние и крупные: длина черепа колеблется от 8 до 60 см, общая длина — от 0,6 до 3,5 м и, возможно, более. Группа обнаруживает много примитивных признаков.
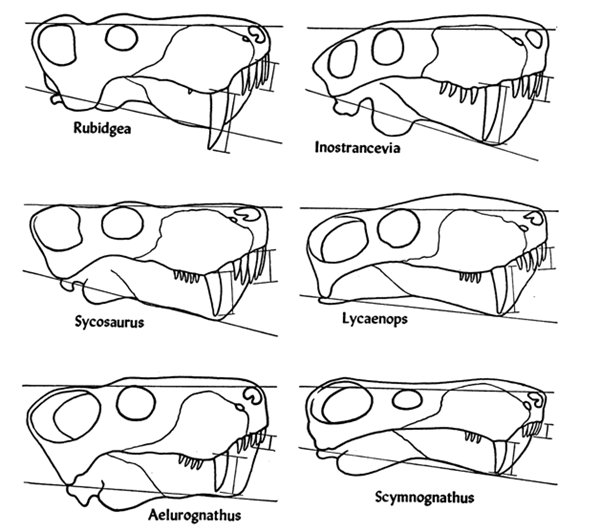
Черепные пропорции некоторых горгоноповых.
Череп крупный, напоминающий в профиль топор. Лицевая часть массивная и высокая, варьирует от относительно короткой до вытянутой. Ноздри разделены на конце морды. Спереди череп заканчивается загнутыми вниз предчелюстными костями. Верхнечелюстные кости слабо сочленены с остальным черепом и могли обладать некоторой подвижностью — эта особенность, возможно, обеспечивала более эффективный укус. Септомаксилла широко выходит на поверхность черепа. Предлобная кость сравнительно сильно выдается вперед. Заднелобная кость неизменно сохраняется. Глазница не превосходит по размерам височное окно. Последнее расширено вниз и назад, обращаясь главным образом наружу. Заглазничная дуга узкая или расширенная. Скуловая дуга опущена и резко расширена с латеральным отгибом в сквамозной части, так что плоскость височного окна ориентирована несколько косо вверх, либо узкая скуловая дуга полого изгибается вниз и плоскость височного окна практически латеральна.
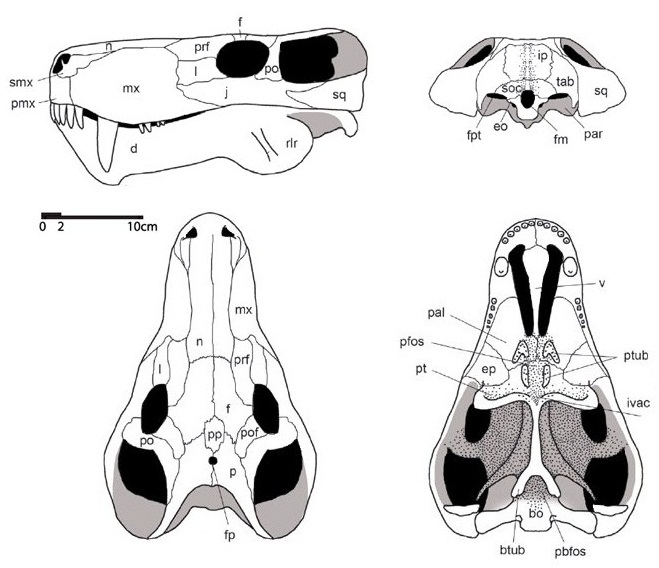
Прорисовки реконструированного черепа с нижней челюстью горгонопса (Gorgonops torvus).
Верхний край височного окна ограничен длинным задним отростком заглазничной кости, смыкающимся сзади с чешуйчатой костью. Чешуйчатая кость толстая, с мощным отростком, окаймляющим сзади скуловую дугу. Из-за расширения височного окна чешуйчатые кости выступают далеко за уровень затылка. Теменная область широкая, как у пеликозавров. Межтеменная кость отсутствует, или очень мала, неправильной формы, либо крупная, ромбическая. Теменные кости расширены. Обычно имеется предтеменная кость, расположенная впереди пинеального отверстия. Последнее почти всегда развито. У некоторых горгоноповых на крыше черепа между глазницами имеются мелкие ямки, в которых могли находиться какие-то рецепторы — возможно, электрические. Затылочный мыщелок остается одиночным.
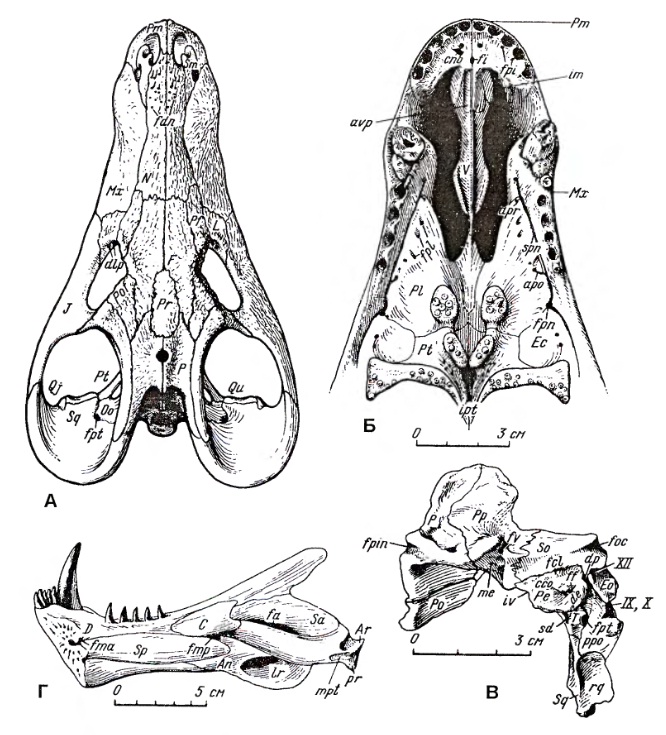
Прорисовки черепного материала завроктона (Sauroctonus progressus): A — череп сверху, Б — снизу (передний отдел), В — фрагмент области темени и затылка, вид вполоборота сбоку и сверху, Г — нижняя челюсть изнутри.
Небо сводчатое. Вторичного неба нет хотя предсошники срослись в одно образование. В носовой полости имеются турбиналии, служившие основой для хрящевых обонятельных раковин. Хоаны открываются между верхними клыками. В области хоан обычны ямки для нижнечелюстных клыков. Сошник непарный. Подглазничные окна отсутствуют. Крыловидные кости обычно соприкасаются с сошником. Верхняя крыловидная кость узкая. Базикраниальный отдел крыловидных костей удлиненный. Межптеригоидные ямы исчезают. Квадратные ветви крыловидных костей широко прилегают к основанию черепа, и вырезка для челюстной мускулатуры кнаружи от них очень велика. Базиптеригоидные отростки хорошо развиты. Базиптеригоидное сочленение подвижное. Имеются парные сближенные небно-птеригоидные бугры, на которых обычно расположены по 2 группы небных зубов; зубы развиты и на поперечных флангах птеригоидов.
Нижняя челюсть массивная. Зубная кость не намного больше других элементов нижней челюсти, хотя велика и имеет восходящую ветвь. Ее венечный отросток развит умеренно либо высокий. Наружный отросток угловой кости (ангулярный лепесток) укороченный и не доходит до заднего конца нижней челюсти. Для ряда горгоноповых характерен массивный, выступающий далеко вниз и зачастую на конце загнутый вперед ретроартикулярный отросток сочленовной кости, внешне имеющий сходство с концевым ушным отростком среднего уха. Нет достаточных свидетельств, что к этому отростку действительно прикреплялось конечное звено среднего уха. Но о связи ретроартикулярного отростка с механикой челюстного сочленения также нет достаточных данных.
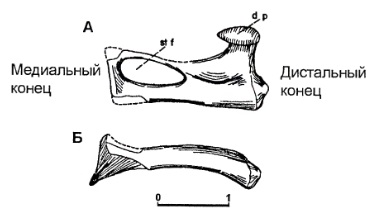
Прорисовки левой слуховой косточки завроктона (Sauroctonus progressus): A — сзади; Б — сверху.
У Suchogorgon, а возможно, еще у ряда горгоноповых в области среднего уха и заднего конца нижней челюсти развит своеобразный синдром признаков: грацильное уплощенное стремя с большим отверстием и без выраженного верхного отростка; длинный и загнутый на конце вперед уплощенный засуставный отросток без явных признаков мышечного прикрепления на нем; желобок на нижнем крае угловой кости впереди от наружного углового лепестка, переходящий позади в желобок на передне-внутреннем крае засуставного отростка. Л.П. Татаринов (2000, 2009 гг.) отмечает также чуть выступающий вниз и назад нижний край углового лепестка.
Возможно, у упомянутых горгоноповых вырезка на нижней челюсти позади углового лепестка поддерживала большую, но несовершенную барабанную перепонку, в задний край которой входит засуставный отросток, но слуховая косточка не связана с этой вырезкой. Во всяком случае, топография костей по краям этой вырезки поверхностно напоминает картину у поздних эмбрионов однопроходных (утконоса). Но на других горгоноповых данное сходство не распространяется.
При этом нужно учитывать, что засуставный лепесток этих горгоноповых обычно плотно прижат к наружной поверхности угловой кости, а не выступает вниз и назад, как его гомолог не только у цинодонтов, но и у однопроходных, и у сумчатых. По Т. Кемпу (1982 г.), у горгоноповых между наружным лепестком и телом угловой кости входит пучок нижнечелюстного аддуктора. Это предположение, хотя оно и не доказано, не следует игнорировать. Варьирует не только морфология слуховой косточки и засуставного отростка, но и в редких случаях венечный отросток нижней челюсти остается сравнительно небольшим. Значение этих вариаций пока непонятно.
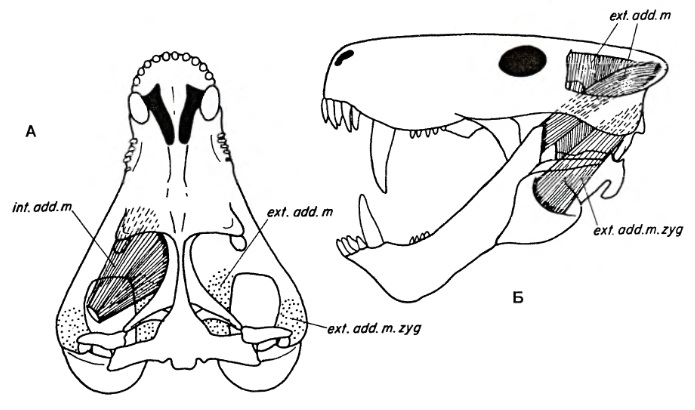
Реконструкция челюстной мускулатуры горгоноповых: A — вид сверху (слева изображен наружный челюстной аддуктор, справа пунктиром отмечены средний и глубокий (скуловой) аддукторы); Б — вид сбоку (средний и скуловой аддукторы).
Пасть могла широко открываться за счет целого ряда уникальных конструкционных особенностей челюстного аппарата, позволяющих нижней челюсти сильно откидываться вниз. Наиболее полно данная способность реализовалась у поздних, самых специализированных горгоноповых. Наблюдается стрептостилия: квадратная кость обладает подвижностью (признак, достоверно установленный лишь у форм с высоким венечным отростком).
По крайней мере у некоторых горгоноповых (в частности, у Viatkogorgon), квадратная кость, хотя и входит в глубокую впадину, развитую на задненижнем крае чешуйчатой кости, не образует с ней настоящего сочленения, а, по-видимому, остается туго соединенным связками. Квадратная кость по наружному краю латерального мыщелка соединяется швом с уменьшенным в размерах квадратноскуловой костью и, несомненно, двигалась вместе с ней. Над своим медиальным мыщелком квадратеая кость соединяется связками с эпиптеригоидом, птеригоидом и стременем, так что самостоятельная подвижность комплекса квадратной и квадратноскуловой костей невелика. Но даже при легком скольжении квадратной кости по чешуйчатой при открывании пасти происходит смещение латерального мыщелка квадратной кости слегка вперед, а медиального мыщелка — слегка назад и наружу. Заметим, что при закрытой пасти, по крайней мере у прогрессивных горгоноповых, латеральный мыщелок расположен относительно позади медиального.
Т. Кемп (1969 г.) полагает, что у горгоноповых имелась настоящая стрептостилия, при которой только верхний конец квадратной кости остается связанным с чешуйчатой костью, а нижний конец отходит вперед. Это мнение разделяет и А.Ю. Соколов (1994 г.). Описанную у вяткогоргона конструкцию с очень незначительным диапазоном подвижности квадратной кости Л.П. Татаринов (2009 г.) назвал псевдострептостилией. Совершенно особую форму кинетизма описал М.Ф. Ивахненко (2003 г.) у российского Suchogorgon. По мнению автора, сочленение между квадратной и скуловой костями функционально неподвижно, обе названные кости участвуют в обеспечении подвижности нижней челюсти.
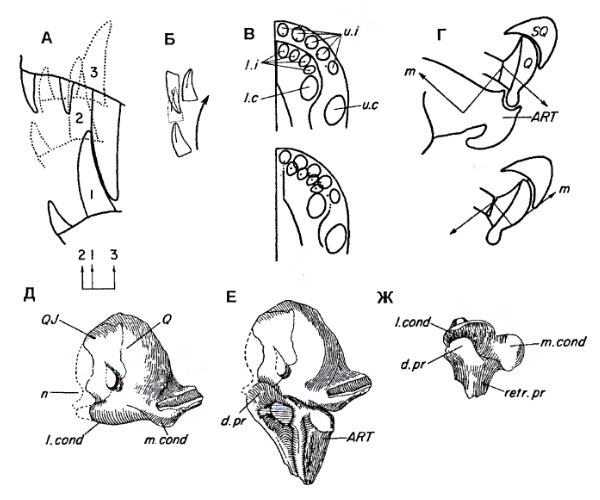
Прорисовки передних верхнечелюстных зубов (A-В) и челюстного сустава (Г-Ж) горгоноповых.
Оригинальный механизм работы челюстного аппарата предложил А.Ю. Соколов (1994 г.), исходивший из принимаемой им внутричелюстной подвижности нижней челюсти. Он считал, что зубная кость подвижно по отношению к задним постдентальным костям. Движения осуществляются под действием специальной части наружночелюстного аддуктора, прикреплявшейся к венечному отростку, при этом зубная кость скользит вдоль верхнечелюстных резцов и клыка. В пользу того свидетельствует обычно наблюдаемая некоторая обособленность дентальной части нижней челюсти от постдентальной. Но на материале хорошей сохранности в ряде случаев отчетливо видно, что обе части челюсти с внутренней стороны широко перекрываются короноидом и предсочленовной костью, причем никаких следов их подвижности относительно друг друга нет (например, у Gorgonops). Так что проблема подвижности нижней челюсти у горгоноповых нуждается в дополнительном исследовании.
Л.П. Татаринов (2009 г.) полагает, что в горизонтальном смещении нижней челюсти при прикусе основную роль играет стрептостилия или псевдострептостилия, а не смещение зубной кости относительно постдентальных костей челюсти. Особенно большое значение имеет горизонтальное смещение челюсти у Inostrancevia и Pravoslavlevia, утративших все заклыковые зубы на нижней челюсти и имевших необычное уплощенное небо. В привлечении внимания к горизонтальной работе нижней челюсти у горгоноповых основная заслуга принадлежит А.Ю. Соколову (1994 г.), хотя на незначительное смещение назад при прикусе нижней челюсти с ее резцами указывал также Т. Кемп (1969 г.). В механике движения нижней челюсти у горгоноповых остается много неясного, но с мнением М.Ф. Ивахненко (2003 г.) о неподвижности челюстного сустава хотя бы у некоторых горгоноповых типа Viatkogorgon и Suchogorgon согласиться трудно.

Череп с нижней челюстью иностранцевии (Inostrancevia alexandri). Палеонтологический музей им. Ю.А. Орлова РАН, Москва.
В верхней челюсти обычно имеется 5 резцов, 1 клык и не более 5 заклыковых зубов. Все зубы конические, заостренные. Резцы сравнительно крупные. Клыки большие, саблевидные, в сечении округлые или сплюснутые с боков, режущего типа. Корни верхних клыков очень длинные, чем и объясняется большая высота черепа в лицевом отделе. Снаружи клык вкладывается в выемку на латеральной поверхности нижней челюсти, опускаясь нередко до ее нижнего края. В связи с этим следует отметить, что на старых реконструкциях длина верхних клыков часто преувеличена, так как на ископаемых черепах клык "вываливается" из альвеолы. В сумке верхнечелюстного клыка сзади функционирующего обычно помещается и практически сформированный замещающий клык, а иногда еще один или несколько меньших. Заклыковые зубы мелкие, остроконические, иногда утрачены. Клыки демонстрируют тенденцию к увеличению, тогда как заклыковые зубы — к полному исчезновению. Таким образом, вся мощь укуса явно сосредотачивалась в передней части челюстей.

Портретная реконструкция иностранцевии (Inostrancevia alexandri). Автор — А. Атучин.
Горгоноповые были способны охотиться на крупную добычу, сопоставимую по размерам с ними самими. Хищник вцеплялся в жертву клыками, а затем как бы вырезал из нее большой кусок резким продольным рывком. Слабое закрепление клыка в верхней челюсти вместе с некоторой подвижностью составляющих ее костей могло обеспечивать функцию амортизации в челюстном аппарате и «гасило» рывки от сопротивления схваченной добычи. Охота на крупных растительноядных могла обернуться потерей клыка, но таким образом предотвращался более серьезный травматизм. Именно на случай потери клыков — основного орудия нападения — в клыковой сумке имелись сменные зубы, находящиеся в зачаточном состоянии и способные за короткое время развиться в новый функциональный клык. Более того, замена клыков у горгоноповых происходила регулярно вне зависимости от наличия или отсутствия повреждений.

Слева:верхнечелюстной клык иностранцевии (Inostrancevia alexandri). Палеонтологический музей им. Ю.А. Орлова РАН, Москва.
Справа: передняя часть нижней челюсти неназванного гогонопида из позднепермских отложений формации Усили, Танзания.
Вырезаемые из добычи куски плоти быстро заглатывались целиком, без дополнительной обработки в ротовой полости. Дело в том, что во время расчленения добычи передними зубами задняя часть расположенных между клыками хоан оставалась открытой и дышать еще было можно. Но когда хищники начинали проглатывать кусок мяса, то пища закрывала хоаны и дыхание становилось невозможным. Кроме того, небольшие и малочисленные (в случае их наличия) заклыковые зубы годились лишь для фиксации, но никак не пережевывания кусков добычи. Подобным образом питались и другие примитивные терапсиды, в связи с чем их передние зубы развиты сильнее.

Схема действия зубочелюстного аппарата рубиджеи (Rubidgea atrox). Автор — М. Антон.
Посткраниальный скелет внешне напоминает характерный для бегающих млекопитающих. Лопатка узкая, со слабо выраженным акромиальным отростком. Клейтрум иногда сохраняется. Грудина окостеневает. Подвздошная кость расширенная. Тазовый пояс без тироидного отверстия, но запирательное отверстие лобковой кости иногда увеличено.

Смонтированные скелеты горгоноповых: A — иностранцевия (Inostrancevia alexandri), длина масштабной линейки 20 см; B — лиценопс (Lycaenops ornatus), длина масштабной линейки 15 см; C — завроктон (Sauroctonus parringtoni), длина масштабной линейки 25 см.
Конечности длинные и стройные. Передняя их пара обычно занимала растопыренное положение, когда плечевая кость ориентирована почти горизонтально. Однако они могли выпрямляться при более быстром передвижении, а также для того, чтобы животное смогло широко открыть пасть — иначе нижняя челюсть заденет землю. Строение бедренной кости сравнимо с таковым у крокодилов и демонстрирует сходную способность к 2 типам локомоции: с растопыренными задними конечностями, как у примитивных амниот, и с бедром, направленным под углом около 45 °, что позволяло нижней конечности двигаться в парасагиттальной плоскости. По-видимому, горгоноповые могли бегать рысцой, хотя и не очень быстро и недалеко. До этого все терапсиды (да и прочие наземные позвоночные) передвигались только медленным шагом. Изменения в строении голеностопного сустава и стопы, отличающие терозавров, слабо заметны у горгоноповых. Сохраняется примитивная фаланговая формула 2 - 3 - 4 - 5 - 3 или 4 с незначительным уменьшением длины "избыточных" фаланг III-го и IV-го пальцев. Имелись крупные когти. Хвост относительно короткий.
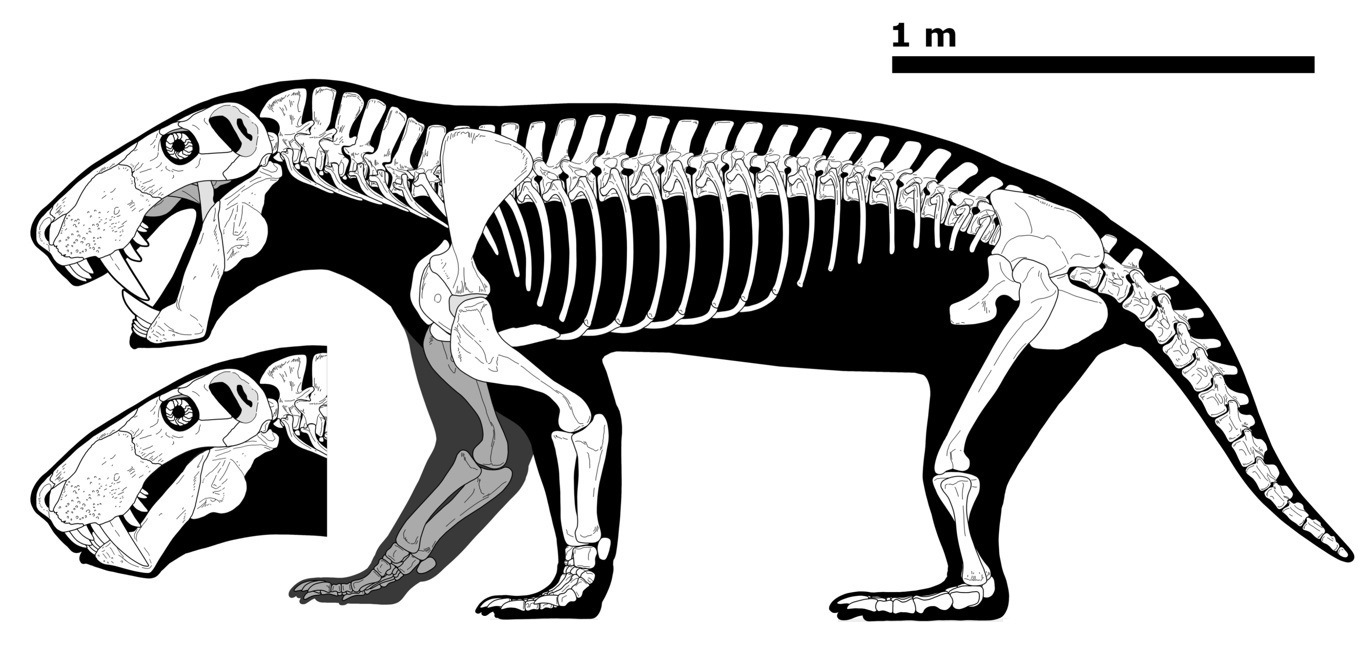
Прорисовка реконструированного скелета иностранцевии (Inostrancevia latifrons). Автор — И. Белоусов.
Головной мозг мал и крайне примитивно устроен. Самой развитой его частью являлся мозжечок — это обеспечивало хорошую координацию движений и свидетельствует о большой подвижности. По крайней мере у некоторых горгоноповых имелись 2 органа пинеального комплекса, причем с пинеальным отверстием в крыше черепа связан задний орган.
Из анализаторов важнейшим мог являться зрительный. Строение склероткальных колец свидетельствует о развитии сумеречного зрения. Звукопроводящий аппарат включал структуры, расположенные в задней части нижней челюсти. Он очень своеобразен и механизм его действия до конца не установлен. Слуховой проход шел от нижней челюсти вверх-назад к выемке в задней части черепа, где могло находиться наружное ухо. Высказано предположение, что горгоноповые слышали звуки в воздухе только тогда, когда открывали рот — тогда кости задней части нижней челюсти касались черепа. Также существует мнение, что с возрастом они утрачивали слух — кости становились слишком толстыми для проведения звуков. По мнению других исследователей, возможность восприятия воздушных звуковых колебаний горгоноповыми полностью исключена. Наличие обонятельных раковин может свидетельствовать о хорошем обонянии. Отпечатки кожных структур, имеющиеся на черепных костях ряда видов, порой трактуются в качестве электрорецепторов (межглазничная область) или даже оснований вибрисс (конец морды).
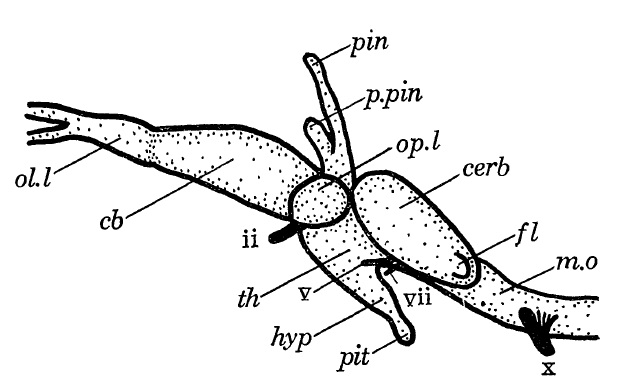
Реконструкция головного мозга горгонопса (Gorgonops torvus); сбоку.
Представители рассматриваемой группы более продвинуты по сравнению с бурнетиидами в связи с расширением аддукторной камеры и уменьшением относительного размера глазниц. Клыки еще более выражены, а заклыковые зубы редуцированы.
Согласно результатам остеогистологического анализа, большинство найденных образцов горгоноповых принадлежит субадультным особям. Особенности роста костей позволяют предположить, что они достигали близких взрослым размеров, прежде чем становились половозрелыми. Вполне вероятно, что за счет интенсивного роста в детстве это происходило довольно быстро.