Строение и функции
Обыкновенный шерстистый носорог известен не только по многочисленным находкам отдельных костей и целого ряда более или менее полных скелетов, но и по сохранившимся в результате естественной мумификации ископаемым трупам с мускулатурой, кожей, шерстью и рогами, а также по оставленным верхнепалеолитическим человеком изображениям. Все это позволяет реконструировать его внешний облик и образ жизни с гораздо большей точностью, чем любого другого ископаемого носорога.
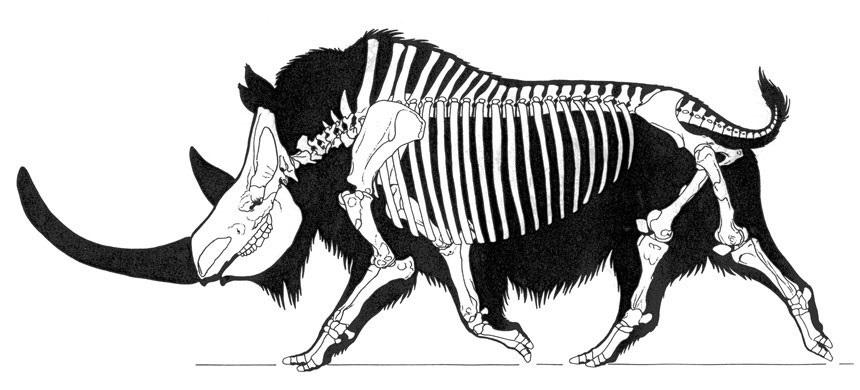
Смонтированный скелет шерстистого носорога (Coelodonta antiquitatis).
Этот носорог представлял собой крупное и грузное животное, достигавшее от 3,2 до 4,5 м в длину и 1,4-2 м в холке (в среднем — 3,5 м и 1,55 м соответственно), а его вес варьировал в пределах 1,8-4 т. Такой разброс по размерам объясняется достаточно выраженной вариабельностью различных популяций. Например, носороги из Украины и Якутии в целом были относительно небольшими, а забайкальские и из Кузнецкой котловины — крупными; значительной величиной отличались и северокитайские экземпляры.
Прорисовка скелета шерстистого носорога (Coelodonta antiquitatis), вписанного в телесные очертания.
Длина большинства черепов обыкновенного шерстистого носорога составляет приблизительно 70-86 см (в среднем — 77 см), а ширина в скулах — около 29-36 см (в среднем — 32 см). Массивная нижняя челюсть может достигать 60 см в длину и 10 см в высоту. В Иркутской области и Забайкалье были обнаружены черепа, достигающие 90 см в длину, а с юга Сибири известна бедренная кость длиной 56 см, для шерстистого носорога очень большая. Примечательно, что для некоторых регионов (например, Якутии) предполагается измельчание позднеплейстоценовых популяций носорогов, в то время как для других областей (Южная Сибирь и др.) таких данных не выявлено.
Таким образом, по своим габаритам крупный шерстистый носорог сопоставим с ныне живущим белым носорогом (Ceratotherium simum) и даже несколько превосходил его, но на большей части ареала он достигал величины черного носорога (Diceros bicornis), при этом шерстистый носорог был массивнее и приземистее упомянутых представителей семейства. В своем биоценозе по величине и силе этот зверь уступал только шерстистому мамонту (Mammuthus primigenius).
Череп с нижней челюстью и рогами шерстистого носорога (Coelodonta antiquitatis) из Сибири.
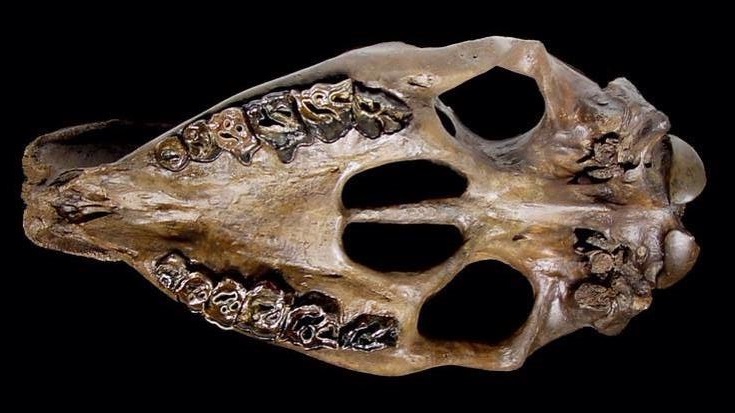
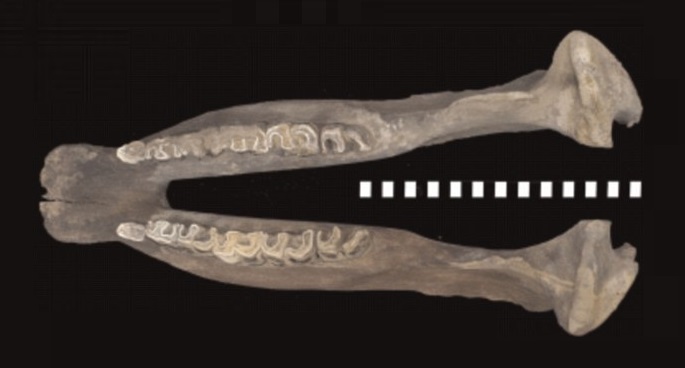
Череп шерстистого носорога удлиненный, с мало прогнутым носолобным профилем. Носовые кости длинные и широкие. Спереди они загибаются вниз, характерно закругляясь, и срастаются с предчелюстными костями; у более ранних представителей рода концы этих костей не срастаются. Имеется сплошная перегородка, находящаяся между носовыми проходами и сросшаяся с сильно развитыми носовыми и предчелюстными костями, у взрослых особей полностью окостеневающая. Необходимо отметить, что у ископаемых родов Stephanorhinus и Dihoplus, а также ныне существующих Ceratotherium, Diceros, Rhinoceros и Dicerorhinus эта перегородка сохраняется хрящевой на протяжении всей жизни или окостеневает не полностью. Носовой вход черепа узкий и длинный. Затылочный отдел сильно отклонен назад, так что плоскость затылка образует с линией основания черепа тупой угол. Симфизный отдел нижней челюсти расширен.
Череп шерстистого носорога (Coelodonta antiquitatis); снизу.
Существование современных носорогов в тропическом климате способствует ослабеванию отбора в сторону усиления признаков полового диморфизма. Шерстистый носорог обитал в условиях холодного климата. Неблагоприятные факторы внешней среды способствовали некоторому усилению признаков полового диморфизма. У шерстистого носорога черпа самцов отличаются от черепов самок следующими признаками: внешний контур черепа самцов отчетливо профилирован, с более резкими изгибами в области перехода от носовых костей к лобным, и далее, к теменным; усилена скульптурированность поверхности костей черепа; большее развитие лицевого отдела черепа за счет укрупнения верхнечелюстных и носовых костей; носовые кости длиннее и шире; теменные кости массивные; теменная площадка более выпуклая; расстояние между теменными линиями, оконтуривающие височные ямки, больше; на теменной площадке развит сагиттальный бугорок; глазничная область черепа отчетливо выступает за счет большего развития слезного, надглазничного и заглазничного отростков; скуловые дуги более массивные и широкие; затылочные гребень крупнее; внешняя поверхность затылочной кости отчетливо скульптурирована; затылочная кость высокая и широкая; яремный и сосцевидный отростки развиты сильнее.
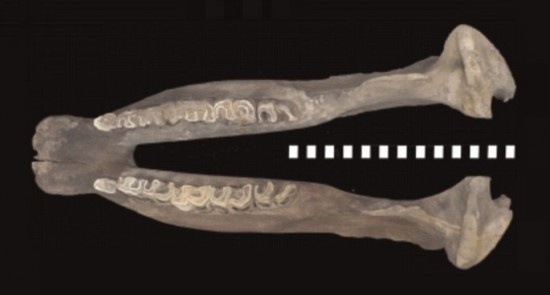
Нижняя челюсть шерстистого носорога (Coelodonta antiquitatis); сверху.
Голова шерстистого носорога ориентирована наклонно по отношению к туловищу, с сильно опущеной вниз мордой. Морда имела прямоугольные очертания, напоминая таковую белого носорога. В объемистую ротовую полость вело отверстие шириной около 35 см, ограниченное губами с веревкообразным краем. Широкая и толстая верхняя губа с ороговевшим краем несколько выдавалась над нижней. Нижняя губа была снабжена в углах рта бугорками, входящими в соответственные углубления верхней губы. Конец морды, включая верхнюю и нижнюю губу, был покрыт короткой и мягкой шерстью. Ноздри заметно меньше, чем у белого носорога (Сeratotherium simum). Глаза не крупнее, чем у белого носорога (4-5 см в диаметре) и глубоко погруженные в глазницы, окруженные густой сетью морщинок. Наружное ухо расположено на большом кожном бугре и в начальной своей части имеет вид короткой трубки, которая затем переходит в ушную раковину до 24 см длиной и 8 см шириной, сильно заостренную на конце. Шерсть густым слоем покрывала ушную раковину и образовывала на ее конце кисточку из более грубого остевого волоса.
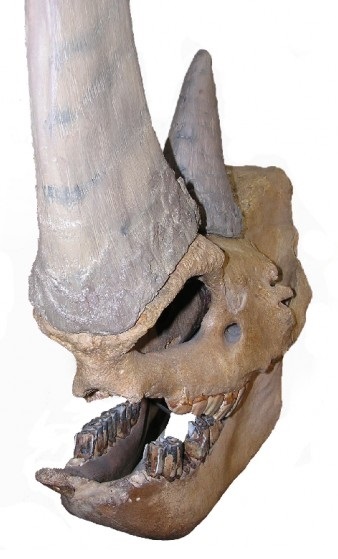
Расположение на носовых костях рога шерстистого носорога (Coelodonta antiquitatis).
На утолщениях носовых и лобных костей, представляющих собой шершавые и прочные мозоли размером примерно 10 × 25 см, сидело два крупных рога. Шероховатости мозолей образованы экзостозами — мелкими шипообразными костными выростами высотой 0,4-0,8 см. Они служили дополнительной защитой кровеносным сосудам, питающих рог, от сильного сжатия, когда животное активно работало носовым рогом при разбивании плотного снежного наста и разгребании снега. Степень выраженности экзостозов указывает на степень развития рогов у различных особей. Очевидно, усиление рабочих функций рога приводило к увеличению его размеров.
Срединное узкое поле на носовых костях, свободное от экзостозов, вмещало главную питающую носовой рог артерию. От конца носовых костей, к центру срединного поля, проходит костный валик, который входил в соответствующее углубление на подошве рога. Он служил лучшему его укреплению. Наличие костного валика на носовых костях указывает на воздействие сил, направленных на боковые поверхности рога, когда животное поворачивало голову вправо и влево при разгребании снега. На черепах, где экзостозы развиты слабее, заметно увеличена площадь срединного поля.
Носовой рог шерстистого носорога (Coelodonta antiquitatis) возрастом более 30 тыс. лет. из урочища Юннюген, Якутия.
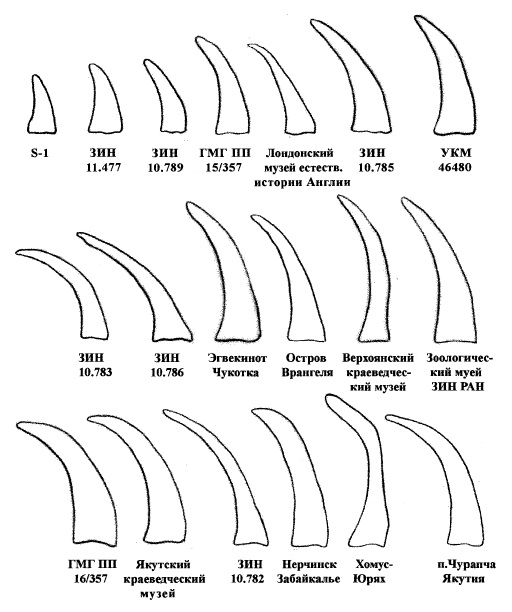
Передний (носовой) рог очень длинный, широкий у основания и утончающийся кверху, уплощенный с боков и саблевидно изогнутый назад, с острым концом. При виде сбоку рог имеет плоскую переднюю грань и покрыт многочисленными поперечными бороздами. Длина переднего рога по большой кривизне составляет 70-140 см (в среднем — 100 см), длина по хорде — 62-102 см (в среднем — 78 см), наибольший диаметр у основания — 10-24 см (в среднем — 17 см), вес — 10-15 кг. Передний рог достигает кончика носовых костей и направлен вперед в существенно большей степени, чем у современных носорогов. В отличие от современных африканских носорогов, для которых характерна практически круглая или четырехугольная форма основания рогов, основание переднего рога шерстистого носорога имело форму эллипса (слегка трапециевидное) и заметно вытянуто в передне-заднем направлении (длина основания примерно в 1,5 раза превышает ширину). Характерно уплощены уже носовые рога молодых особей. Второй, лобный рог значительно короче — 30-48 см в длину и весом до 5 кг, иногда слегка изогнутый, с поперечным сечением от треугольного до овального, но имел в целом типичную для носорогов конусовидную форму. В отличие от шерстистого носорога, у современных видов оба рога примерно округлые в сечении.
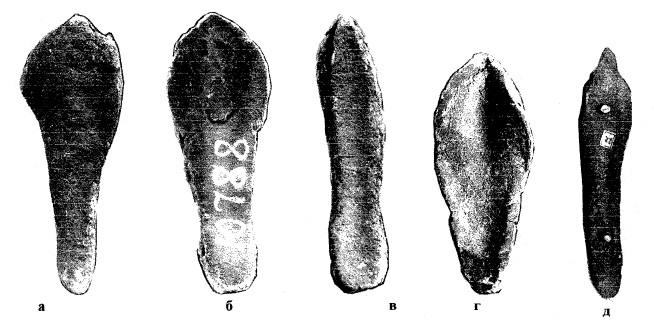
Изменчивость формы основания носовых рогов шерстистого носорога (Coelodonta antiquitatis).
Надо отметить, что саблевидная форма носового рога у шерстистого носорога не имеет аналогов среди ныне живущих носорогов. Заметно вытянутое в передне-заднем направлении основание рога характерно также для суматранского носорога (Dicerorhinus sumatrensis), который филогенетически наиболее близок к шерстистому носорогу среди сохранившихся видов. Однако после износа кератина на периферийной части рога суматранского носорога он приобретает коническую форму, характерную для других современных носорогов, а не латерально уплощенную и саблевидную форму рога шерстистого носорога. Увеличение передне-задней длины плотной сердцевины носового рога у шерстистого носорога, вероятно, имеет функциональное объяснение. При виде сбоку передняя грань носового рога шерстистого носорога имеет отчетливую фасетку износа, которая визуально "срезает" передний изгиб рога. Причем в некоторых редких случаях рог может быть изношен более чем на половину своей передне-задней длины.
Эта плоская грань разделена на левую и правую грани, что указывает на то, что она образовалась в результате активных боковых движений головой в разные стороны. Наиболее вероятной причиной стирания передней грани рога является его использование при разрушение снежного наста и/или ледяной корки (джуда). Это единственный известный пример использования носорогами рога для питания. Удлиненная в передне-заднем направлении плотная сердцевина носового рога давала шерстистому носорогу резерв устойчивости к такому виду износа. Вероятно, более тонкая конусовидная сердцевина была бы недостаточно устойчивой для ее продолжительного использования, что могло бы привести к перелому рога и утрате его функциональности.

Поперечный (А) и продольный (B) спилы дистальной части носового рога шерстистого носорога (Coelodonta antiquitatis). Длина масштабной линейки — 1 см.
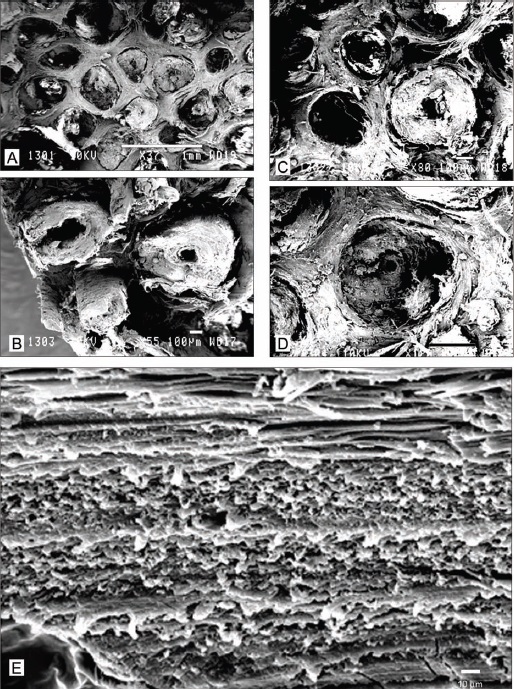
Составляющее рог кератиновое вещество состоит из темно-бурых или сизоватых стерженьков-филаментов, погруженных в ячеи полупрозрачной основы (матрикса). Стерженьки ориентированы вдоль длинной оси рога, но не строго параллельны друг другу, поскольку незначительно извиты и переплетаются. Их форма варьирует от правильной цилиндрической до немного сглаженной трех-пятигранной. Последняя сформировалась, вероятно, из-за большей механической нагрузки на периферийные отделы рога. Толщина филаментов различная: самые толстые (до 600 мкм) расположены в центральной области рога, наиболее тонкие (до 64 мкм) — на периферии. Толщина межфиламентных перегородок составляет 30-160 мкм. Расстояние между филаментами в срединной части рога стабильно (100-200 мкм) и здесь они разделены более толстыми перегородками матрикса. На периферии же слои основы тоньше и стерженьки сомкнуты теснее.
Отдельный филамент состоит из вложенных один в другой 4-5 цилиндров разной толщины. Боковые поверхности цилиндров соединены между собой каркасом из клеточных выростов, а ребристая поверхность наружного цилиндра закреплена в ячеях матрикса кутикулярными чешуями. Сердцевинная часть филамента представлена полостью разного диаметра и овальной или треугольно-многоугольной формы. Внутренние стенки полости слоистые, губчатые, образованные сплющенными, вытянутыми по окружности филамента клетками. Полость заполнена пронизанной капиллярами соединительной тканью, которая питает растущий рог.
Электронограммы срезов дистального отдела носового рога шерстистого носорога (Coelodonta antiquitatis): А, B — поперечные срезы центральной части; C, D — поперечные срезы на границе между срединной и периферийной частями; E — продольный срез отдельного филамента. Электронограммы. Масштабные линейки: А — 1 мм; B — 100 мкм; E — 10 мкм.
Эллипсоидная форма носового рога определяла дальнейшее уплощение его рабочей части, которое было связано со спрессовыванием филаментов срединно-верхушечной части рога под воздействием нагрузок, когда животное употребляло рог для добывания пищи. Кроме того, боковые филаменты имеют большие диаметры центральных полостей и неплотно скреплены между собой, вследствие чего довольно легко отслаивались с боковых плоскостей у основания рога. Сердцевина рога значительно более твердая, чем его внешние области из-за более длинных и плотнее расположенных филаментов. Слоистость матрикса и люфт между его слоями делали рога при жизни животного прочными и упругими, а после его гибели они легко отделялись от черепа. При разрушении в осадках боковые части носовых рогов оказываются менее прочными и поэтому многие сохранившиеся образцы имеют необычную форму саблевидно изогнутых плоских досок толщиной 3-3,5 см. Филаменты лобного рога расли равномернее и одинаково прочно соединялись между собой, благодаря чему они сохранили свою объемную форму. Как и у современных носорогов, в 5-10 см над основанием рогов шерстистого носорога происходило стремительное разрушение наружных слоев кератина, в результате чего рог приобретал форму, которая соответствует более плотной меланизированной и кальцифицированной сердцевине. Эта часть благодаря ее большей устойчивости к разрушению и сохраняется в ископаемом виде.

Рога шерстистого носорога (Coelodonta antiquitatis). Слева — носовой и лобный рога одной особи с реки Тирехтях, Якутия; на передней поверхности носового рога хорошо заметно прижизненное стирание. Справа — лобный рог той же особи с прижизненно заполированной вершиной.
Оба рога несут одинаковое количество поперечных полос разной ширины, каждая из которых состоит из двух частей — темной и светлой. По направлению от основания к середине рога полосы располагаются заметно чаще. К вершине рога они все более разрежаются, расстояние между ними больше. Имеется прямая зависимость количества поперечных полос от размера рога шерстистого носорога. Чем рога больше, тем большее количество полос на нем отмечается. В вершинной (апикальной) части рога расстояние между полосами колеблется от 4 до 6-7 см, у основания оно равно 1,5-3,5 см. Эти полосы образовались в результате неравномерности сезонного роста молодых пучков филаментов: в зимний период рост рогов замедлялся, а в летний ускорялся. При этом более темная часть полосы представляет летние месяцы, а более светлая — зимние. Таким образом, индивидуальный возраст животного можно определять, подсчитывая количество полос на роге. Наибольшее зафиксированное их число — 35.
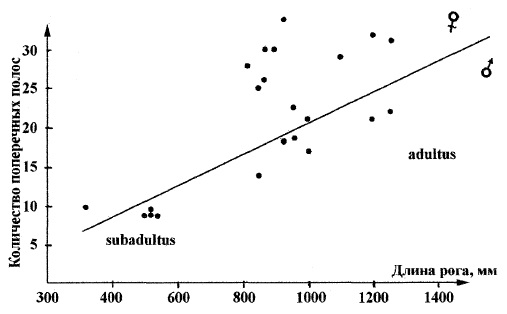
Зависимость количества поперечных полос и длины рога у шерстистого носорога (Coelodonta antiquitatis).
Поперечные полосы, соответствующие зонам годового прироста, слабо просматриваются на рогах ныне живущих африканских носорогов. Однако на них невозможно выделить участки сезонных (зимне-летних) приростов. Поскольку современные носороги живут в областях с постоянно теплым климатом, сезонные колебания в приросте филаментов рога, по-видимому, не выражены столь сильно, как у шерстистого носорога, который обитал в суровых климатических условиях. Нарастание филаментов у них идет одинаково равномерно. Согласно замерам, скорость роста рога шерстистого носорога была сопоставима с таковой современных носорогов, являясь максимальной у молодых особей.
Крупные носовые рога шерстистого носорога заметно различаются по форме. Одни из них массивные и широкие, плавно сужающиеся к вершине. Другие рога, широкие в базальной части, затем резко сужаются, становясь пикообразными. Вершина их заострена. Причину подобных отличий усматривают в половом диморфизме. Широкие и массивные рога, по-видимому, принадлежали старым самцам, а тонкие и длинные — самкам шерстистого носорога. У ныне живущих самцов черного (Diceros bicornis) и белого (Ceratotherium simum) носорогов носовые рога мало различаются по размерам от рогов самок. С другой стороны, у самок этих видов зарегистрированы наибольшие размеры рогов: для черного носорога 138 см, для белого — 158 см. Зато рога самцов заметно отличаются большим весом и массивностью. Отличия в форме рогов самок и самцов у шерстистого носорога, возможно, были такими же, как и у современных африканских носорогов.
Вариации в размерах и форме носового рога шерстистого носорога (Coelodonta antiquitatis).
Формула постоянных зубов: I0/i0, C0/c0, P3/p3, M3/m3. Формула молочных зубов: dI1/di2, dC0/dc0, dP4/Dp4. Постоянные рудиментарные резцы сохраняются лишь в примерно 4 % случаев. Крупные щечные зубы, по 6 с каждой стороны верхней и нижней челюсти, имеют высокие коронки с утолщенной эмалью и большим количеством цемента. Форма зубов угловатая, призматическая; сужение зуба, от основания к вершине, выражено слабо. Внешняя и внутренняя стенки зуба направлены под углом 40 ° одна к другой, внутренняя стенка почти отвесная. Жевательная поверхность плоская. Поверхность эмали варьирует от грубо- до слабо-морщинистой, на боковых сторонах коронки грубо шероховатая. Внешняя поверхность края эктолофа заполирована у морф, обитающих в залесенных ландшафтах. Внутренний конец протолофа направлен параллельно эктолофу. Металоф отходит от протолофа в передней части зуба. Средняя долинка имеет почти параллельные стенки, сходящиеся только на дне долинки. Средняя долинка по направлению к внутренней части зуба суживается, а при стирании замыкается.
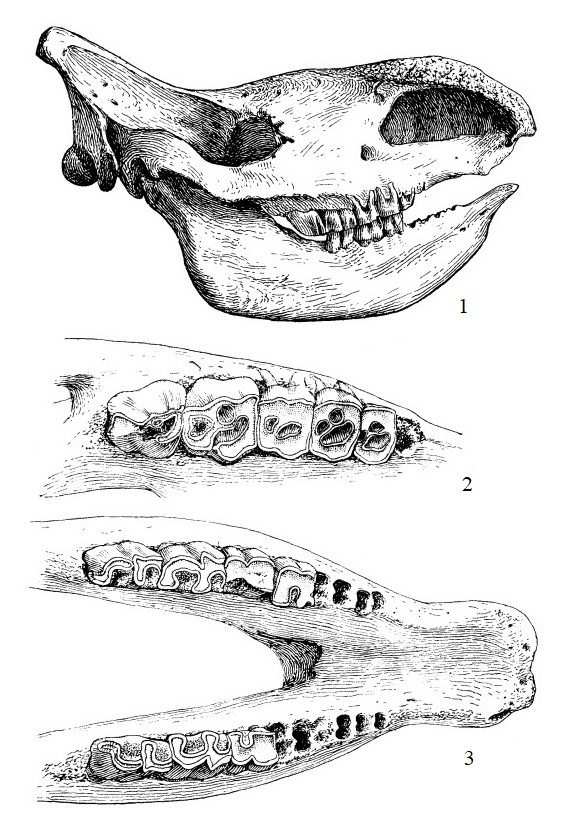
Прорисовки черепного материала шерстистого носорога (Coelodonta antiquitatis): 1 — череп; 2 — правые Р3-М3; 3 — передний отдел нижней челюсти с коренными зубами.
Криста и кроше хорошо развиты, не разветвлены. При стирании зуба криста и кроше сливаются друг с другом, образуя дополнительную замкнутую фасетку. Замыкание задней долинки и образование задней фасетки происходит уже при несильном стирании зуба. Поперечные гребни на верхних коренных направлены очень косо к продольной оси зуба. Парастиль резко обособлен позади глубокой бороздой на внешней стенке зуба. Метакон на внешней стенке хорошо выражен. Цингулюм на передней стороне зуба образует вертикальную складку, на границе, с внутренней стороны зуба, снижающуюся до основания коронки. Подобно другим представителям рода, зубы имеют открытую внутреннюю полость, явллясь гипсодонтными (т. е., обладали постоянным ростом).
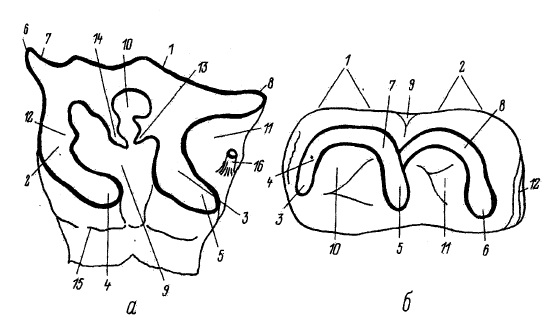
Схема строения М2 и m2 шерстистого носорога (Coelodonta antiquitatis) и обозначение основных морфологических деталей. Для М2 (а): 1 — эктолоф; 2 — протолоф; 3 — металоф; 4 — протокон; 5 — гипокон; 6 — парастиль; 7 — парастильная складка; 8 — метастиль; 9 — главная средняя долинка; 10 — добавочная долинка; 11 — задняя долинка; 12 — антикроше; 13 — кроше; 14 — криста; 15 — цингулюм; 16 — стебелек. Для m2 (б): 1 — металофид; 2 — гиполофид; 3 — параконид; 4 — парастилид; 5 — метаконид; 6 — энтоконид; 7 — протоконид; 8 — гипоконид; 9 — наружная долинка; 10 — передняя внутренняя долинка; 11 — задняя внутренняя долинка; 12 — цингулюм.
В позвоночнике насчитывается 7 шейных, 18 грудных, 4 поясничных, 4 крестцовых и около 20 хвостовых позвонков. Подобно многим другим млекопитающим, имеющим тяжелую и большую голову, шейные позвонки короткие и очень высокие. Остистые отростки первых четырех грудных позвонков, особенно III-го и IV-го, массивные и длинные (до 36 см). Остистые отростки последней трети грудного и поясничного отдела позвоночника короткие.
В задней части шеи поднимался горб, переходивший над лопатками в мощный бугор, образованный сильно развитыми остистыми отростками передних 4 грудных позвонков. К их концевым утолщениям крепилась выйная связка, удерживающая и фиксирующая тяжелую голову животного, а также увеличивающая жесткость позвоночника в грудном отделе.Также горб содержал значительное количество жира, служившего в качестве запаса питательных веществ на случай бескормицы. За бугром спинной профиль опускался, образуя седловину, ограниченную сзади двумя возвышениями, расположенными на высоте поясничных позвонков и крестца. Линия живота гармонировала с линией спины. Туловище бочкообразное, вытянутое. В паховой области у самок находилась пара сосков. Хвост широкий, сильно сплющенный у основания. Снизу и с боков его покрывали грубые волосы, росшие по направлению к концу хвоста и здесь расходившиея в стороны в виде широкой, веерообразной кисти.
Тело поддерживали относительно короткие, массивные конечности. Задняя их пара немного длиннее передней. Длинные кости конечностей широкие и массивные, с хорошо выраженными буграми и гребнями. Плечевая кость слабо изогнутая, с мало расширенным нижним эпифизом. Бедренная кость с прямым диафизом, обладающим дополнительным вертелом посередине. Большая и малая берцовые кости частично сращены. Однако, поскольку обе кости довольно часто находят по отдельности, срастаться они были должны в довольно позднем возрасте. Кисть массивнее стопы, ее подошва значительно больше. Кисть и стопа трехпалые. Пальцы разделены и оканчивались ногтеобразными копытцами. Средний палец более крупный, чем два боковых. Центральный (III-й) метаподий особенно массивный, прямой. Внутренний (II-й) и наружный (IV-й) метаподии короче и слегка изогнуты.
Передние конечности шерстистого носорога (Coelodonta antiquitatis).
Кожный покров в целом почти лишен морщин и совсем без сегментов, но с вкраплениями волосяных фолликулов. Кожа не настолько грубая, как у современных носорогов. Ее толщина варьировала на различных участках тела от 0,5 до 2 см, причем наиболее тонкой она была на голове, а самой толстой на груди и плечах. Благодаря тому, что кожа имела тонкий слой внешнего рогового эпидермиса, многочисленные сальные и потовые железы функционировали нормально и стимулировали рост густого волосяного покрова. Его длина достигала 20-30 см, что позволяло выдерживать экстремально низкие температуры последнего оледенения. Распределение волос сходно с таковым у шерстистого мамонта (
Mammuthus primigenius) и яка (
Bos grunniens): на макушке головы, шее и холке более длинные, жесткие и густые волосы создавали подобие гривы, на животе, груди и основании ног длинные остевые волосы образовывали подвес, конечности были покрыты намного более короткой шерстью. Под грубым кроющим волосом находился тонкий густой подшерсток. Несомненно, что длинная шерсть отрастала только в зимнее время, а в летний период она была короткая и не такая густая. Все это наглядно можно наблюдать у современных животных, живущих в условиях умеренного и северного климата. Сохранившиеся образцы волос из мерзлой почвы — светло-бурого цвета, иногда с желтоватым оттенком. Мех детенышей был гораздо короче, относительно мягче и имел более светлый окрас. Некоторые образчики пещерного искусства предполагают наличие более темного "пояса" или полосы вокруг центральной части туловища животного.

Бегущий шерстистый носорог (
Coelodonta antiquitatis). Автор — М. Антон.
Ряд внешних особенностей строения шерстистого носорога свидетельствует об отличной приспособленности к длительным сильным морозам. Так, его уши были относительно короче и намного уже, чем у носорогов тропических широт, хвост тоже относительно намного более короткий. Такие черты свойственны вообще всем животным, обитающим в холодном климате, поскольку более короткие уши и хвост уменьшают общую площадь тела, сводя к минимуму тепловые потери. Толстая и волосатая шкура также снижала потерю тепла организмом. Холодный зимний воздух вдыхался через небольшие ноздри и хорошо прогревался, контактируя с капиллярной сетью слизистой в длинных и узких носовых дыхательных ходах от входа носового отверстия до хоан. Сильное развитие экзостозов на носовых костях указывает на активное использование переднего рога в зимнее время для добывания корма из под уплотненной корки снежного наста, которая часто образуется на открытых, обдуваемых ветром пространствах. По сравнению с ныне живущими сородичами, шерстистый носорог более коротконогий, а его голова и туловище относительно более вытянуты.
Портретная реконструкция шерстистого носорога (Coelodonta antiquitatis).