Состав
Вопрос о подвидах шерстистого носорога до конца не прояснен, однако большинство источников сходится в том, что на огромном ареале существовало довольно много локальных и хронологических форм этого животного. Помимо мелких морфологических деталей, они отличались телосложением и размерами.
Различия в строении шерстистых носорогов, живших в конце среднего и в позднем плейстоцене Западной Европы, были выявлены давно. В основном они выражаются в массивности скелета, а также в степени отклоненности назад затылочной поверхности черепа, со временем только возраставших. Это дало К. Герену (1980 г.) основания раделить вид на 2 подвида: Coelodonta antiquitatis praecursor, представляющего собой более грацильную среднеплейстоценовую форму (впервые установлен из рисских отложений Македонии на севере Греции), и Coelodonta antiquitatis antiquitatis, классическую массивную форму из позднего плейстоцена. Более поздние исследования предполагают, что ранний подвид на самом представляет собой позднюю форму Coelodonta tologoijensis, а в качестве Coelodonta antiquitatis рассматривают лишь позднеплейстоценовых носорогов. Однако такая интерпретация спорна.
Смонтированный скелет шерстистого носорога (Coelodonta antiquitatis) из позднего плейстоцена Украины.
В Азии ситуация еще сложнее. Б.С. Русанов (1968 г.) выделил для территории Якутии 3 подвида шерстистого носорога: Coelodonta antiquitatis pristinus из среднего плейстоцена, а также Coelodonta antiquitatis jacuticus и Coelodonta antiquitatis humilis из позднего плейстоцена. При описании новых форм автор основывался на различиях размерных показателей, степени развития задней долинки верхних коренных и развитии дополнительных складок зубной эмали. Выводы Русанова недостаточно обоснованы, так как он не учитывал индивидуальные и возрастные особенности стирания зубов у шерстистого носорога. Для сравнения выделенных подвидов он берет единичные черепа их, которые неодинаковы по возрастным фазам. В результате последующих исследований подвид Coelodonta antiquitatis pristinus был признан невалидным, так как приводимый автором голотип без номера и описания местонахождения, а размеры и диагностические признаки вполне соответствуют среднеплейстоценовому носорогу.
В результате многолетних исследований П.А. Лазарев, С.Е. Григорьев и В.В. Плотников (2010 г.) пришли к выводу, что в Якутии существовало 2 подвида шерстистого носорога: Coelodonta antiquitatis jacuticus и Coelodonta antiquitatis antiquitatis. Костные остатки Coelodonta antiquitatis jacuticus встречаются в ограниченном количестве на всей территории Якутии. Черепа, зубы, нижние челюсти и другие кости наиболее часто находили в бассейнах рек Алдан, Вилюй, Яна, Колыма. Подвид существовал в среднем плейстоцене, ареал его был приурочен к открытым степным пространствам. Coelodonta antiquitatis antiquitatis в Якутии обитал в позднеплейстоценовое время, остатки этого животного встречаются повсеместно, но значительно реже, чем мамонта, бизона и лошади.
Смонтированный скелет шерстистого носорога (Coelodonta antiquitatis) из позднего плейстоцена Сибири.
Для Китая выделяются 3 последовательно сменяющих друг друга подвида — Coelodonta antiquitatis shansius из первой половины среднего плейстоцена, Coelodonta antiquitatis yenshanensis из второй его половины и Coelodonta antiquitati chilinensis из позднего плейстоцена. Отмечаются отличия в строении черепа азиатских и европейских шерстистых носорогов.
По мнению Н.В. Гарутт (1998 г.), в пределах ареала шерстистого носорога начиная с конца среднего плейстоцена и затем в позднем плейстоцене одновременно существовали два отдельных, хотя и близкородственных вида, эволюционно приспособленных к различным условиям обитания в нестабильном климате — степной Coelodonta lenensis и лесостепной Coelodonta antiquitatis. Впервые они были выделены в ранге подвидов Coelodonta antiquitatis pristinus и Coelodonta antiquitatis humilis, установленных по единичным образцам из плейстоценовых отложений Якутии. В то же время сама автор исследования подчеркивает, что вопрос о разделении шерстистого носорога на два вида требует дальнейшего изучения.
Ленский шерстистый носорог (Coelodonta lenensis (Pallas, 1772)) отличался крупными размерами, низко опущенной к земле крупной головой, приземистым туловищем и укороченными конечностями. Массивный череп имеет удлиненные пропорции, с длинным и низким лицевым отделом. Длинные предчелюстные кости образуют обширную поверхность перед предкоренными зубами. Носовые кости длинные и широкие, часто с сильно развитыми экзостозами в области крепления носового рога, который по длине превышал размеры головы животного.
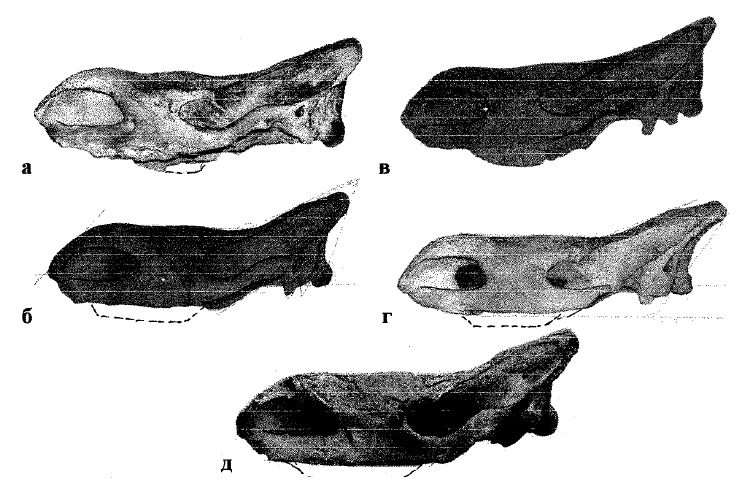
Морфологические вариации в черепе степной морфы шерстистого носорога (Coelodonta lenensis) (по Гаррутт, 1998 г.): а — село Веденяпино, Наровчатский уезд, Пензенская губерния; б, в, г — Казанская губерния, Средняя Волга; д — Восточная Сибирь.
Боковые, щечные области между носочелюстной вырезкой и передним краем глазницы, широкие и плоские. Это указывает на то, что верхней губой управляла сильная мускулатура, которая обеспечивала простые функции поднятия и опускания ее. Подвижность верхней губы была ограничена. Височная область характеризуется сильно удлиненными, узкими височными впадинами. Скуловые отростки височных костей слабо выступают в боковых направлениях и совместно со скуловыми костями образуют спрямленные и слабо выдающиеся в стороны скуловые дуги. Плоская теменная поверхность черепа полого приподнята над плоскостью лба и составляет с ней угол 158-160 °.
Затылочный гребень значительно развит и сильно выступал над затылочной плоскостью черепа. Мышцы головы, осуществлявшие боковые движения, имели достаточную площадь крепления на массивном затылочном гребне, что в свою очередь свидетельствует о их большом развитии и силе. Работа эти мышц была важна при поступательном движении головы в боковых направлениях, когда животное разгребало снег.
Плоскость затылочной чешуи ровная. К базальной плоскости черепа угол ее наклона составляет 90-110 °. На очень крупных черепах (более 85 см) выйная поверхность чешуи под затылочным гребнем имеет углубление для лучшего закрепления выйной связки. Затылочные мыщелки крупные, умеренно выступающие на затылочной кости и и отстоящие на большом расстоянии друг от друга. Они имеют больший угол наклона (развал) от вертикальной прямой, опущенной отвесно через затылочное отверстие к базальной оси черепа. Особенности строения затылочной поверхности указывают на ограниченное движение головы в верхне-нижних направлениях.
Крылья крыловидной кости длинные и низкие, слабо выступающие над плоскостью основания черепа. Лицевая и мозговая части черепа находятся приблизительно в одной плоскости. Такое строение черепных костей способствовало уменьшению общей высоты черепа, и соответственно, нижней челюсти, что уменьшало усиление сжимающего действия челюстей за счет ослабления действия мышц-аддукторов (крыловидной жевательной мышцы). При этом увеличивалась подвижность нижней челюсти в горизонтальном направлении, что важно было для перетирания жесткой травянистой пищи. Нижняя челюсть длинная с низким телом и короткими суставными и мышечным отростками. Симфизный отдел нижней челюсти удлинен и расширен у внешнего края.
Коронки щечных зубов крупные и высокие, по форме приближенные к призме. Эмаль утолщенная, грубо-морщинистая за счет развития в микроструктуре крупных эмалевых призм. Парастильная складка эктолофа слабо развита.
Особенности строения черепа, которые проявляются в общих удлиненных пропорциях его лицевого и мозгового отделов, удлинении зубного ряда и усилении гипсодонтности зубов, а также общие пропорции определяют адаптацию ленского шерстистого носорога к открытым степных ландшафтам, где основной корм составляла травянистая растительность. Зимой их приходилось добывать, проламывая мордой и носовым рогом корку наста и рагребая снежный покров. По аналогии с населяющим степи африканским белым носорогом (Ceratotherium simum), данный вид мог образовывать относительно многочисленные группы, в несколько десятков особей, которые можно было назвать стадами.
Собственно шерстистый носорог (Coelodonta antiquitatis (Blumenbach, 1799)) менее крупный, с более легким и стройным по строению телом. Голова была более короткой и высокой, а также более горизонтально поставленной. Очевидно, сокращение длины лицевого отдела черепа произошло в результате смены жесткого травянистого корма на корм более мягкий, в котором трава сочеталась с молодыми побегами и листьями. Отверстие костного носового входа короче и шире. Носовые кости короче, сужаются к концу и загибаются книзу.
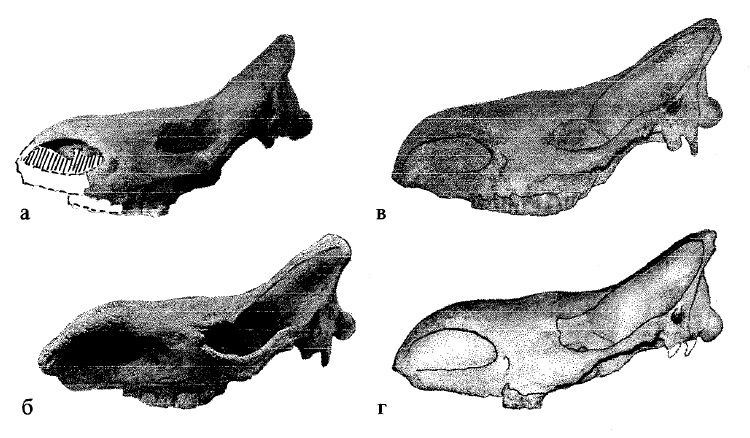
Морфологические вариации в черепе лесостепной морфы шерстистого носорога (Coelodonta antiquitatis) (по Гаррутт, 1998 г.): а — бассейн реки Улахан-Гаиллях (система реки Яны), Якутия; б — бассейн реки Малый Анюй, Якутия; в — деревня Новый Ильмовый куст, Буинский уезд, Симбирская губерния, Средняя Волга; г — Казанская губерния, Средняя Волга.
Уменьшение площади носовых костей не связано прямой зависимостью с уменьшением развития экзостозов, но, как правило, чаще встречаются черепа данного вида со слабо развитыми экзостозами. По меньшему развитию экзостозов на носовых костях можно предполагать, что передний рог не был таким массивным и длинным. Поскольку носовые кости имели изгиб книзу, передний рог был направлен вперед. Даже при небольшом опускании головы животное могло обеспечить горизонтальную постановку рабочей грани рога к земле. Судя по всему, этот вид обитал в более мягком климате, чем ленский шерстистый носорог, когда снежный покров не был таким глубоким и плотным.
Щечные области более рельефные, края у носочелюстной вырезки значительно углублены, по сравнению с внешними, глазничными краями. Такое строение щечной области указывает на дифференцировку в развитии и строении лицевых мускулов, поднимающих верхнюю губу, что в свою очередь указывает на большую подвижность верхней губы у лесостепного вида шерстистого носорога по сравнению с степным видом. Височные впадины черепа у лесостепного вида шерстистого носорога более широкие и короткие, чем у степного. Скуловые отростки височных костей значительно выступают в боковой плоскости и формируют со скуловыми костями резко изогнутые и сильно выступающие скуловые дуги.
Теменная плоскость черепа круто приподнята над плоскостью лба и составляет с ней угол 140-150 °. Затылочный гребень развит не так сильно, как у степного вида. Затылочная плоскость выпуклая, образует к базальной плоскости черепа угол наклона 75-90 °. Затылочные мыщелки крупные, выпуклые, сильно выдающиеся назад и отстоящие на небольшом расстояние друг от друга. Они имеют меньший угол наклона (развал) от вертикальной прямой, опущенной отвесно через затылочное отверстие к базальной оси черепа. Затылочный отдел черепа высоко приподнят над лицевым за счет увеличения высоты верхнечелюстной кости, а также крыловидной кости в основании черепа. Это создает характерную линию излома в основании черепа.
Перечисленные особенности в строении мозгового отдела черепа лесостепного вида шерстистого носорога обеспечивали большую подвижность его головы в верхне-нижнем направлении. Животное могло достаточно легко поднять голову, что важно было для срывания молодых побегов с кустарников и веток с деревьев, для скашивания верхушек тростника в низинных участках русловой поймы. Строение затылочного отдела черепа у лесостепного вида сходно со строением такового у современных черного (Diceros bicornis) и индийского (Rhinoceros unicornis), которые также относятся к лесному морфотипу, что служит прямым доказательством того, что данный вид шерстистого носорога существовал в лесостепях и редколесьях.
Верхнечелюстная кость высокая, что в целом увеличивало высоту черепа. Нижняя челюсть укорачивается. Становится высоким тело нижней челюсти, удлиняются суставные отростки, увеличивается площадь мышечного отростка. Изменяется угол прикрепления жевательной мускулатуры в височных ямках и на крыловидных костях черепа, и на внутренней поверхности тела и восходящей ветви нижней челюсти. Такое строение способствовало усилению сжимающего действия челюстей через усиление челюстных мышц адукторов. Два ее пучка, наружный и внутренний, имели достаточно большую площадь крепления на широких небных и крыловидных костях. Это способствовало увеличению сдавливающих движений челюстей и лучшему измельчению смешанного корма с увеличением содержания в нем веточно-лиственного компонента.
Соразмерно с черепом уменьшились в размерах коронки зубов, заметно сократив общую длину зубного ряда. Эмаль зубов тонкая, средне- или слабоморщинистая. Парастильная складка эктолофа хорошо развита.
Итак, указанные морфологические особенности Coelodonta antiquitatis указывают на его приспособленность к питанию не только травянистым, но и веточно-листовым кормом. Животное держалось в низинных участках русел рек, где были особенно распространены заросли ивняка, березы, осины. Участки, занятые осоками и пушицами также могли служить для него хорошим пастбищем. По мнению Н.В. Гарутт, данный вид существовал в небольших группах, но вероятно не в таких малочисленных, как у африканского черного носорога.
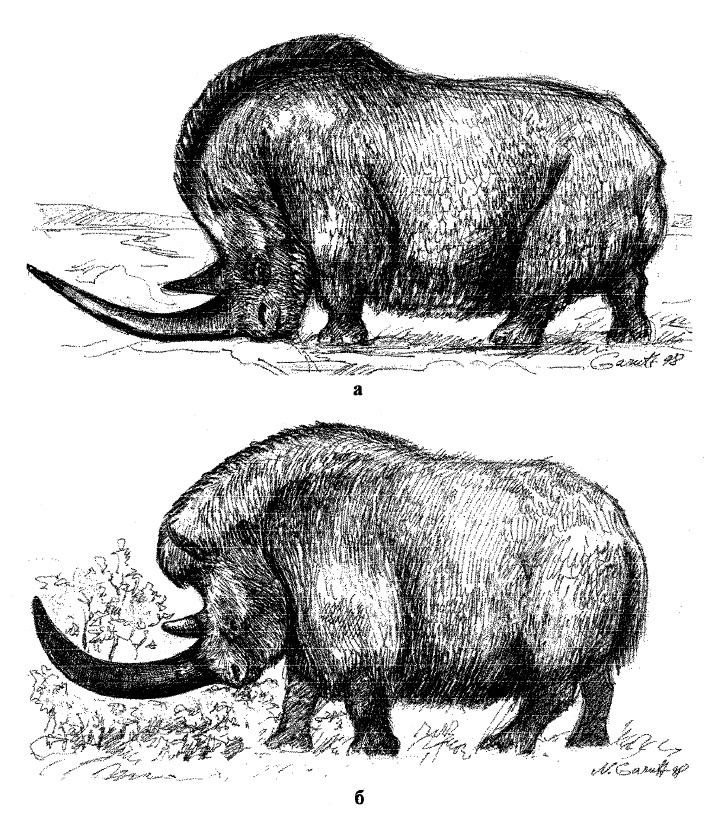
Реконструкции внешнего облика ленского шерстистого носорога (Coelodonta lenensis) (а) и обыкновенного шерстистого носорога (Coelodonta antiquitatis) (б). Автор — Н.В. Гарутт.
Coelodonta lenensis, более близкий к эволюционно исходной форме, сформировался в условиях холодного климата открытых степей Центральной Азии, быстро проник в северные широты и распространился в Забайкалье и Якутии. Во время максимального наступления ледников он распространился по остальной Азии и заселил Восточную Европу. В ходе расселения в другие регионы, где климат и ландшафтная обстановка отличались от первоначальных, в процессе дальнейшей эволюции оформился новый вид, приспособленный к существованию в условиях лесостепей и редколесий. Согласно этой точке зрения, в позднем плейстоцена Западной Европы водился Coelodonta antiquitatis.
В холодные ледниковые фазы позднего плейстоцена, во время максимального развития открытых ландшафтов в Евразии, Coelodonta lenensis расселился на территории Восточно-Европейской равнины. Вероятно, он в небольшом количестве также проник в Западную Европу, но не получил там широкого распространения. Местный лесостепной вид Coelodonta antiquitatis сохранял свою нишу и препятствовал расселении мигранта из Северной Азии. Во время межледниковых фаз позднего плейстоцена, когда происходило потепление и увлажнение климата и увеличение произрастания древесной и кустарниковой растительности, Coelodonta antiquitatis расширял свой ареал, в свою очередь проникая из Западной Европы в Восточную и дальше на восток в Сибирь до Якутии включительно.
Вероятно, в это время произошло объединение двух видов и сосуществование их на общей площади обширного ареала. При совместном обитании между 50 и 60 ° с. ш. на территории Восточной Европы, юга Западной, Восточной Сибири и Забайкалья по численности преобладал степной Coelodonta lenensis, а в Западной Европе и Северо-Восточной Сибири — лесостепной Coelodonta antiquitatis.
В конце вюрмского оледенения численность носорогов рода Coelodonta резко уменьшается в Западной Европе, что в дальнейшем привело к полному их вымиранию. В это время в Восточной Европе область распространения обоих видов значительно сужается. Очевидно это было связано с развитием в конце плейстоцена в субарктической части Восточной Европы и Западной Сибири скудных по кормовым ресурсам тундр. Возможно, что при сокращении общей численности двух видов шерстистого носорога их обобщенный ареал распадается на небольшие локальные ареалы, разделенные большими пространствами. Постепенно сужая ареалы, на рубеже позднего плейстоцена и голоцена шерстистые носороги ограничились своей исходной прародиной и обитали в Забайкалье, Монголии и Северном Китае до полного исчезновения.
П.А. Лазарев, С.Е. Григорьев и В.В. Плотников (2010 г.) полагают, что выделенные Н.В. Гарутт виды обоснованы недостаточно, не выделены номенклатурные типы, описания неполные, не обоснован их геологический возраст. Rhinoceros lenensis, описанный П. Палласом по черепу и нижней челюсти с реки Чикой в Забайкалье, на которого ссылается автор, скорей всего относится к раннесреднеплейстоценовому Coelodonta tologoijensis. Авторы допускают, что этот вид позже мог проникнуть из Забайкалья в Якутию, но этот вопрос остается открытым.
Палеогенетика
Группе ученых во главе с Э. Лорд (2020 г.) из генетического материала шерстистых носорогов из северо-восточной Сибири, живших в период от 50 до 14 тыс. лет назад, удалось секвенировать 1 полный ядерный и 14 митохондриальных геномов. Анализ мтДНК позволил выявить у шерстистых носорогов 13 митохондриальных гаплогрупп. Все они принадлежали к двум эволюционным линиям, которые разошлись около 205 тыс. лет назад. Приблизительно 154 тыс. лет назад от первой линии отделилась еще одна клада, уникальная для острова Врангеля. Высокое разнообразие митохондриальных геномов сохранялось вплоть до самого вымирания вида — более того, 86-22 тыс. лет назад в пределах обеих клад наблюдалась быстрая диверсификация.
На основе генетических данных исследователи построили демографическую модель популяции шерстистого носорога, согласно которой в период 110-14 тыс. лет назад численность вида не снижалась. Напротив, приблизительно 29,5 тыс. назад она какое-то время росла, а затем оставалась стабильной вплоть до самого исчезновения вида. Увеличение популяции могло быть связано с климатическими изменениями, благодаря которым на территории Восточной Сибири сформировались более подходящие условия для холодолюбивого шерстистого носорога.
Судя по ядерному геному особи, жившей 18,5 тыс. лет назад, шерстистые носороги в это время все еще был широко распространены в Восточной Сибири. Появление людей, которые заселили регион около 31,6 тыс. лет назад, не сказалось на их численности. По крайней мере, геномы носорогов не сохранили следов сокращения популяций и близкородственного скрещивания, в отличие от последних мамонтов с острова Врангеля. Это значит, что человек и шерстистый носорог сосуществовали на протяжении по меньшей мере 13 тыс. лет, а основным фактором, который привел к окончательному его вымиранию, являлась не охота.
Кусок кожи шерстистого носорога (Coelodonta antiquitatis), извлеченный из желудка сохранившегося в мерзлой почве щенка, обнаруженного у села Тумат в Якутии. Из этого образца возрастом около 14,4 тыс. лет был получен митохондриальный геном шерстистого носорога.
Авторы предполагают, что в исчезновении этого вида следует винить резкие изменения климата. Судя по полученным ими генетическим данным, шерстистый носорог вымер очень быстро, причем произошло это в период от 18,5 до 14 тыс. лет назад. Это совпадет по времени с резким беллинг-аллередским потеплением, которое началось около 14,6 тыс. лет назад и привело к замещению лесами открытых пастбищ как основной среды обитания шерстистого носорога.
Также был проведен поиск генов, которые позволили шерстистому носорогу приспособиться к холодным климатическим условиям. Для этого его ядерный геном сравнили с геномом ближайшего живого родственника — суматранского носорога (Dicerorhinus sumatrensis). После сопоставления 19556 генов этих двух видов, в геноме шерстистого носорога были выявлены 1524 несинонимичные замены. Среди них есть генетические варианты, связанные с внутриклеточными процессами, метаболизмом и размножением.
В отличие от шерстистого мамонта (Mammuthus primigenius), шерстистый носорог не приобрел особых генов, связанных с накоплением жира и изменениями циркадных ритмов. Тем не менее, некоторые генетические приспособления к холодному климату у двух видов оказались общими. Например, таков специализированный ген TRPA1, который активирует реагирующие на холод кожные рецепторы. У многих млекопитающих на белки воздействует ген KCNK17, что делает их чувствительными к холоду. Однако у шерстистого носорога KCNK17 утратил функциональность вследствие мутации, поэтому его неактивность, вероятно, способствовала адаптации животного к холодному климату. Сходные генные механизмы выявлены и у шерстистого мамонта.
Команда исследователей под руководством П. Зибера (2023 г.) проанализировала найденные в Германии экскременты пещерной гиены (Crocuta crocuta spelaea), поедавшей мясо шерстистого носорога. При этом впервые был реконструирован митохондриальный геном европейского шерстистого носорога. Хотя восстановленные последовательности ДНК демонстрируют признаки значительной деградации, что, возможно, ограничивает полученные выводы, авторы полагают, что они нашли возможные доказательства разделения популяции около 450 тыс. лет назад. Митогеномы европейских шерстистых носорогов отличаются от сибирских, что позволяет предположить их расхождение примерно в то время, когда шерстистый носорог впервые появляется на территории Европы. Появление изменчивости в митохондриальных линиях шерстистого носорога также указывает на то, что динамика популяций на его обширном ареале могла быть очень сложной.