Образ жизни
Сохранившиеся остатки пищи и споро-пыльцевой анализ древних почв указывают на преимущественную безлесность территории, луговой или степной характер растительных сообществ, в которых существовал шерстистый носорог в восточной части своего обширного ареала. На территории Европы он обитал главным образом в лесостепях, редколесьях, островных, пойменных и горных лесах. Нет оснований полагать, что он вел образ жизни, сильно отличный от образа жизни современных носорогов. Судя по всему, шерстистый носорог значительную часть времени пасся на открытых равнинах, предпочитая наиболее богатые кормом места — широкие поймы рек и берега озерных котловин. В благоприятные климатические фазы он также расселялся на более возвышенных участках и плато. Однако высокогорий шерстистый носорог избегал ввиду обилия выпадаемого снега и сильной пересеченности местности, которые сложно было преодолеть.
Шерстистые носороги (Coelodonta antiquitatis) в естественном окружении. Автор — К. Флеров.
На характер пищи шерстистого носорога прямо указывают ее частицы, застрявшие и спрессованные в колодцеобразных лунках эмали щечных зубов, а также сохранившиеся в ротовой полости и желудочно-кишечном тракте мумифицированных трупов. Из зубов черепов носорогов из Казанского геологического музея и Зоологического музея РАН извлечены размолотые частицы древесины мягких лиственных пород (ивы, березы, ольхи) и землистые частицы. В образцах из Иркутского музея из остатков пищи были определены ель, лиственница, пихта, ива, береза, листья, близкие к бруснике, остатки злаков и некоторых других растений.
В связи с развитием в ботанике спорово-пыльцевых методов исследований были повторно изучены растительные остатки из ротовой полости мумифицированной головы Вилюйского носорога. Помимо наличия плодов хвойных, эфедры и побегов ивы, проведенный анализ выявил остатки листьев и пыльцу злаков, осок и сложноцветных, а из хвойных была определена пыльца зерна ели и лиственницы. Состав растений, определенный в остатках пищи Вилюйского носорога, значительно обеднен из-за малого количества пробы. Вероятно, в момент гибели животного в его ротовой полости пищи уже не было. В образцах пищи, добытой из полостей жевательной поверхности зубов мумифицированной головы Верхоянского носорога, выявлено малое количество пыльцы древесных (не более 5 %) и доминирование пыльцы травянистых видов растений (90 %), из которых следует отметить явное преобладание пыльцы злаков (148 зерен) и полыней (37 зерен).
В комках растительной массы из распавшегося желудка и кишок Чурапчинского носорога содержались побеги злаков и осок. Споро-пыльцевой анализ показал наличие большого количества пыльцы травянистых растений степных ассоциаций — злаков (89 %), сложноцветных (4,5 %), полыней (2,5 %), осок, гречишных, маревых, лютиковых, гвоздичных, крестоцветных, мауновых. Пыльца древесных видов (лиственница, береза, ель, сосна) единична, а среди спор преобладает зеленый мох (80,7 %), кочедыжниковые (11,2 %) и сибирский плаунок (3,6 %). Сходные данные были получены и из вмещавших суглинок. Судя по результатам исследований, при жизни этого носорога в Центральной Якутии преобладали луговостепные пространства, перемежавшиеся с небольшими лесами из лиственницы, березы и некоторых видов хвойных. Такое широкое распространение остепненных растительных сообществ характерно для сухого и холодного сартанского оледенения.
Пасущиеся шерстистые носороги (Coelodonta antiquitatis). Автор — З. Буриан.
Находка хорошо сохранившейся туши взрослой самки шерстистого носорога на речке Малая Филипповка в низовьях Колымы позволила провести чрезвычайно подробное исследование остатков пищи из ее желудка. В них доминирует пыльца травянистых растений (98,5 %), в основном степных и лугово-степных видов. Преобладает пыльца злаков (45,9 %) и сложноцветных (40,6 %), главным образом полыней (до 40,1 %). Пыльца деревьев и кустарников (елей, сосен и ив) составляет лишь 0,9 % и очень плохо сохранилась. Вероятно, это свидетельствует о ее случайном попадании в желудок животного. На долю спор печеночных мхов, хвощей, плаунка сибирского и плауна колючего пришлось только 0,6 %. Таким образом, ведущими в рационе этого носорога, как и некоторых других, найденных ранее в более континентальных районах Якутии особей, были различные виды злаков и богатый набор разнотравья.
Рыхлые отложения на месте захоронения туши и прилипший к ее шерсти грунт более полно отражают растительность и климатические условия территории. В отложениях и грунте также преобладает пыльца травянистых растений, среди которых доминируют злаки и полыни. Однако доля разнотравья оказалась ниже, его состав беднее, а количество пыльцы деревьев и кустарников (лиственницы, берез, ольховника, ив), наоборот, больше, чем в пищевом комке в желудке.
В совокупности спорово-пыльцевые анализы содержимого желудка носорога и грунтов на месте обнаружения животного показывают, что 40 тыс. лет назад в низовьях Колымы преобладала степная и лугово-степная растительность, тундровые же элементы в ней были выражены не явно. Ядро степных группировок составляли злаково-разнотравные, полынно-гвоздично-разнотравные и осочково-разнотравные ассоциации. Открытые пространства, вероятно, сочетались с развитыми на склонах северной экспозиции разреженными лиственничными лесами, зарослями ольховника, березовыми и ивовыми ерниками. Такой состав растительности в целом соответствует климатическим условиям каргинского интерстадиала: климат тогда был континентальным, а летние периоды — засушливыми и жаркими.
Реконструкция внешнего облика шерстистого носорога (Coelodonta antiquitatis). Авторы — А. и А. Кеннисы.
Произведенные изотопные исследования серии рогов указали на сезонные измерения рациона их владельцев. Различные участки роста рогов предполагают, что они в основном питались травой летом и объедали ветки кустарников зимой.
Изотопный анализ остатков 71 особей шерстистого носорога практически со всего ареала показывает, что рацион данного вида в общих чертах не менялся на протяжении последних 15 тыс. лет его истории и основывался главным образом на потреблении C3-травянистых растений. В то же время, существуют региональные различия в индивидуальных соотношениях изотопов между западно-центральной Европой, восточной Европой, западно-центральной Сибирью и северо-восточной Сибирью. Данный факт отражает изменчивость ландшафтных условий, свидетельствуя о поведенческой гибкости носорога и его способности приспосабливаться к различным местообитаниям. При этом общим для всех регионов является изменение количественных значений изотопов во время последнего ледникового максимума примерно 20-16 тыс. лет назад. Возможно, данное явление было обусловлено более сухими климатическими условиями.
Соотношение изотопов в остатках шерстистого носорога совпадает с таковым у современных сайгака и овцебыка, но явственно отличается от шерстистого мамонта и лошади той эпохи. Как и шерстистый носорог, двое последних специализировались к открытым пространствам тундростепи и обладали сходным устройством пищеварительного тракта. Вероятно, это стало следствием последовательной экологической дифференциации и межвидовой конкуренции. Сходные отношения наблюдаются, например, между африканским саванным слоном (Loxodonta africana) и черным носорогом (Diceros bicornis), а также между азиатским слоном (Elephas maximus) и индийским носорогом (Rhinoceros unicornis).
Таким образом, рацион шерстистого носорога состоял главным образом из степных трав (злаковые, осоковые, полыни и др.) и побегов кустарников (брусника), включая также, особенно в осенне-зимний период, определенное количество веточек низкорослой древесной растительности — лиственной (ива, береза, ольха) и хвойной (ель, пихта, лиственница). Помимо этого, в рацион входили мхи и плауны. Животное практически не питалось корнями и клубнями, поскольку был лишено резцов, которые позволили бы ему эффективно разрывать грунт в поисках подземных частей растений. В его объемистом брюхе находился вместительный желудок и длинный кишечник, способные справиться с подобным рационом — бедным белками, но богатым трудно перевариваемой клетчаткой. Ввиду свойственного всем непарнопалым относительного несовершенства пищеварительной системы и низкой питательной ценности корма носорог был вынужден был поглощать значительное его количество.

Пасущиеся шерстистые носороги (Coelodonta antiquitatis). Автор — Р. Евсеев.
Анатомическое строение шерстистого носорога дополнительно подтверждает, что он являлся преимущественно пастбищным животным, собирающим корм на уровне своих копыт.
Изучение биологии современных носорогов установило четкую зависимость общих пропорций тела животного от образа жизни и, прежде всего, от характера питания. Так, носороги, пищей которых служат главным образом побеги кустарников и деревьев, обычно имеют относительно высокие конечности и короткое туловище. Подобный тип строения характерен для лесных форм современных носорогов — индийского (Rhinoceros unicornis), суматранского (Dicerorhinus sumatrensis), а также обитателя лесостепи черного носорога (Diceros bicornis). Среди ископаемых носорогов наиболее характерное строение обитателя кустарниковых зарослей и пойменных лесов имеет носорог Мерка ('Dihoplus' kirchbergensis). Наоборот, у степного белого носорога (Ceratotherium simum), питающегося в основном травой, конечности относительно короткие, поэтому туловище кажется сильно вытянутым в длину. Шерстистый носорог также обладал короткими конечностями и длинным туловищем, т. е. по своим пропорциям практически не отличался от травоядного носорога Африки. Кроме того, сходные пропорции характерны для яванского носорога (Rhinoceros sondaicus), питающегося в основном низкорослой древесной и кустарниковой растительностью, а также упавшими на землю плодами.
У носорогов плоскость затылка образует с линией основания черепа угол, величина которого характеризует положение головы по отношению к шее и туловищу. Этот показатель находится в прямой зависимости от образа жизни и характера питания животных. У лесного индийского и лесостепного черного носорогов, объедающих ветки кустарников и невысоких деревьев, величина этого угла меньше, чем у питающегося травой белого носорога. Плоскость затылка шерстистого носорога образует с линией основания черепа тупой угол. Это свидетельствует о наклонном положении его головы по отношению к туловищу, что, очевидно, связано с добыванием травянистой растительности с земли. Среди вымерших носорогов сходным положением головы также обладали населяющие лесостепи узкомордый носорог (Stephanorhinus hemitoechus) и сибирский эласмотерий (Elasmotherium sibiricum). Усиление височной мышцы и шейной мускулатуры шерстистого носорога также указывает на противодействие высокой силе натяжения, возникающей при захвате больших пучков низкорослой растительности.
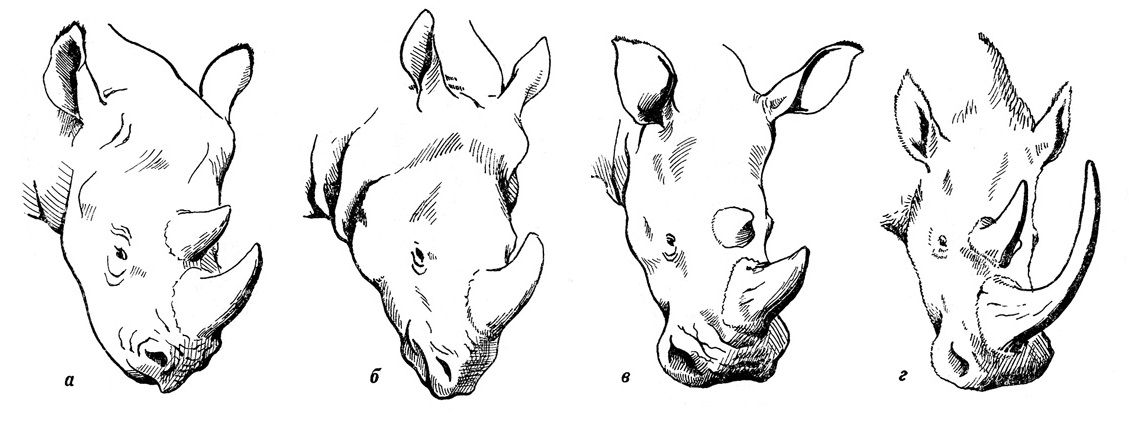
У лесных и лесостепных видов современных носорогов, пищей которых являются преимущественно ветки и листья, сильно развитая верхняя губа довольно длинная, острая на конце, свисающая книзу и очень подвижная. С помощью такой губы, действующей наподобие хоботка, эти животные прихватывают ветки, обрывая с них листья и снимая кору. У степного белого носорога, наоборот, короткая и широкая верхняя губа. У переднего края рта она оканчивается твердой кромкой ороговевшего эпидермиса, служащей для скусывания травы. Исследования мумифицированных голов шерстистого носорога показало, что это животное, так же как и белый носорог Африки, обладает широкой и короткой верхней губой с ороговевшим краем, что подтверждает предположение о питании его травой. Поскольку передние зубы у зверя отсутствовали, захват корма осуществлялся ороговевшими краями губ. Затем он перетирался на высоких и богатых цементом щечных зубах. Часто наблюдается горизонтальное стирание зубов, вызванное содержащимся в травах кремнеземом.
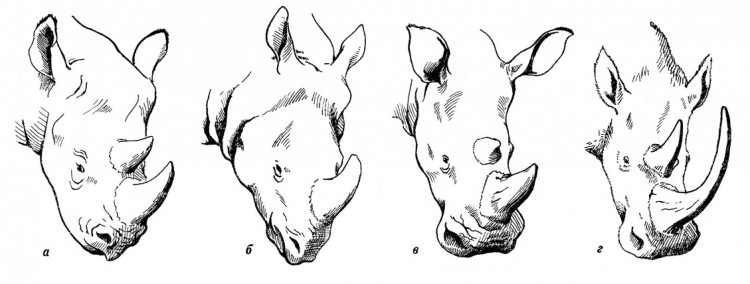
Строение верхней губы носорогов: а — черный носорог (Diceros bicornis); б — индийский носорог (Rhinoceros unicornis); в — белый носорог (Ceratotherium simum); г — шерстистый носорог (Coelodonta antiquitatis).
Климат основного биотопа шерстистого носорога — тундростепи — был чрезвычайно холодным, но весьма сухим и малоснежным. Высота снежного покрова обычно не превышала 20-25 см, что позволяло легко добираться до подснежной растительности. На переднем выпуклом крае практически всех изученных носовых рогов имеется т. н. площадка стирания: стертая и заполированная поверхность, для каждого образца — индивидуальная. Обычно она составляет от 1/2 до 2/3 всей длины рога. У многих найденных рогов более рыхлый периферийный слой разрушен настолько, что обнажена их центральная уплотненная часть. По всей вероятности, площадка стирания возникала за счет трения рога о плотный грунт и снежный наст. Судя по всему, ее наличие свидетельствует о том, что носовой рог наряду с копытами использовался для добывания корма из-под снега в зимнее время. Предполагается, что разгребание снега проводилось боковыми движениями головы. Однако представляется более вероятным, что в условиях малоснежных зим в тундростепи стачивание рога происходило при соприкосновении его переднего выпуклого края с поверхностью земли при пастьбе и вертикальных движениях головой (на себя и слегка под себя), когда животное отрывало пучок травы, как это делают современные полорогие. Стиранию способствовало и сильное смещение носового рога к переднему краю морды животного. Однако вполне возможны оба варианта. Помимо переднего края, при разгребании снега стирались и боковые поверхности носового рога, особенно в его верхних 2/3.

Стертая передняя грань на носовых рогах шерстистого носорога (Coelodonta antiquitatis): А — Чурапчинский носорог (Якутск); Б — образец UHMP 43 (Хельсинки); В — Музей ледникового периода (Москва).
В связи с подобной функцией рога становятся понятными и причины развития у этого животного окостеневшей носовой перегородки — дополнительной опоры для носовых костей, несущих передний рог, и, соответственно, дополнительного приспособления к усиленным нагрузкам на всю морду во время кормежки. Несмотря на это, зверь по возможности старался избегать местностей со значительным снежным покровом, скрывающим пищу и затрудняющим передвижение. Показатель трудности перемещения по снегу у массивного и коротконогого шерстистого носорога среди представителей мамонтовой фауны самый высокий.
Примечательно, что потертости выявлены не только на носовых, но и на лобных рогах шерстистых носорогов, которые никак не могли достать ими до поверхности снега, разгребая его во время жировок. Возможно, эти потертости вызваны ударами о рога конкурентов во время схваток в период размножения. Помимо демонстрации своего статуса и выяснения отношений с сородичами, с помощью своих рогов шерстистые носороги отражали атаки хищников ледникового периода. Также носовой рог, словно клин, раздвигал заросли перед мордой идущего животного.
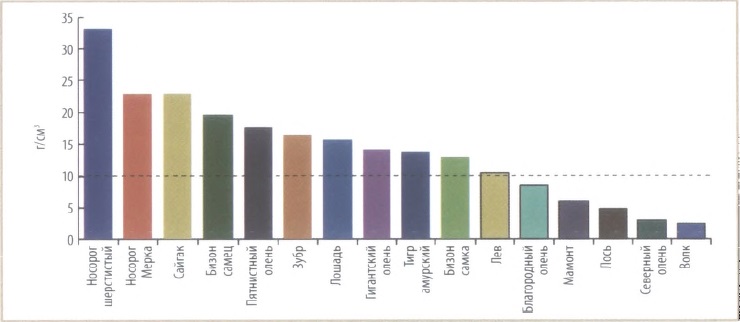
Показатель трудности перемещения по снегу для некоторых современных крупных млекопитающих и представителей мамонтового фаунистического комплекса (по Шпанскому, 2021 г.).
У обладающих значительными размерами и массивными рогами взрослых носорогов практически не было естественных врагов, однако детеныши и подростки наверняка регулярно становились добычей волков (Canis lupus), пещерных гиен (Crocuta crocuta spelaea) и, особенно, пещерных львов (Panthera spelaea). Найден череп со следами зубов, указывающими на нападение крупного хищника из семейства кошачьих, каким мог быть пещерный лев, однако пострадавшее животное дожило до взрослого возраста. В затылочно-теменной области другого черепа отчетливо видны глубокие борозды от когтей крупной кошки (лишь кошачьи при удержании добычи фиксируют одну лапу на загривке жертвы); данная травма была получена уже в достаточно взрослом возрасте. Известны также шейные позвонки молодых особей с характерными следами укусов крупных кошек. Помимо этого, существовали и другие опасности, которым подвергались шерстистые носороги: преодоление пересеченных участков, крутых или скользких склонов и т. п. могли приводить к серьезным травмам или даже смерти отдельных особей.
Поведение и жизненный цикл
Остатки обыкновенного шерстистого носорога встречаются довольно часто, однако их доля в многовидовых местонахождениях не превышает 7-10 %. Одно из редких исключений представляет местонахождение у поселка Сергеево на реке Чулым, где остатки носорога составляют 15 % от общего количества. Здесь около 32 тыс. лет назад существовало пойменное болото, которое привлекало животных сочной растительностью, но становилось естественной ловушкой для тяжелых и громоздких носорогов. Еще реже остатки носорогов встречаются на палеолитических стоянках. Например, на Мезинской стоянке найдено всего 17 костей, что составляет 0,2 % от общей численности остатков всех найденных здесь животных.
Изображения шерстистых носорогов (Coelodonta antiquitatis) из пещеры Шовэ, Франция. Высказано предположение, что они отображают стадность этих животных.
По аналогии с большинством современных носорогов можно предположить, что шерстистые носороги жили семейными группами по 2-3 особи (самка с детенышами) и предпочитали держаться подальше от массовых скоплений других животных. Однако примечательно, что единственный ныне живущий представитель семейства, специализирующийся на травяной пище, белый носорог, собирается в матриархальные группы численностью до 10-15 и даже 20 особей. Такое групповое объединение для степного вида вполне объяснимо, если учесть, что на открытых пространствах, хорошо просматриваемых хищниками, даже очень крупное одиночное животное максимально незащищено. Последнее особенно справедливо в отношении молодых особей. Не исключено, что стадное поведение было характерно и для шерстистого носорога. Самцы современных носорогов держатся одиночно, защищая свой участок от пищевых конкурентов и присоединялись к самкам только в период гона. В это время самцы белого и черного носорога вступают друг с другом в схватки за обладание самками, в которых редко ранят друг друга. Имеющие детенышей самки указанных видов также демонстрируют агрессивное поведение, пытаясь согнать со своих кормовых участков взрослых самцов.
На некоторых черепах шерстистого носорога имеются прижизненные повреждения, половина которых локализованы в области теменных костей. Они представляют собой довольно глубокие вмятины (от 5 до 18 мм) во внешнем слое компакты теменной кости, которая у носорогов очень прочная и толстая. По внешнему краю вмятин нависают валики смещенной в сторону направления удара костной ткани. Благодаря прочности и массивности теменных костей и минимальной толщине мышечного и кожного слоя в этой области, нанесенные раны не были смертельны для животных и быстро заживали, не осложняясь другими болезнями. Другая группа прижизненных повреждений локализована в области носовых костей, глазницы, челюстного сустава. Кроме того, известны сломанные и повторно сросшиеся нижние челюсти и задние ребра. Первичная механическая травма осложнялась посттравматическими заболеваниями — остеомиелитом или артрозом.
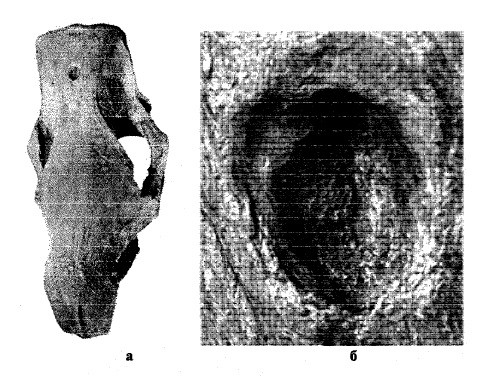
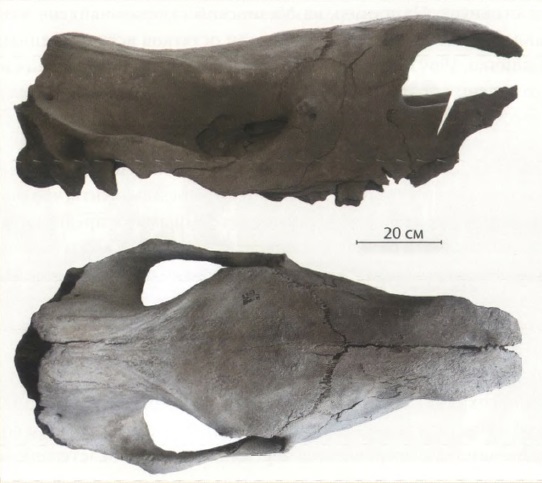
Прижизненно поврежденный череп шерстистого носорога (Coelodonta antiquitatis) со Средней Волги: а — вид сверху; б — фрагмент теменных костей с повреждением.
Вышеуказанные области черепа относятся к жизненно значимым. Они имеют важные группы мышц, связок, нервов и кровеносных сосудов. Ранения в такие области приводили к размозжению мягких тканей и нарушению трофики органа. Заживление ран требовало большего времени, поэтому они инфицировались и осложнялись другими заболеваниями. Как правило, это приводило к скорой гибели животного, о чем свидетельствует отсутствие на большинстве черепов компенсаторных изменений.
Локализация травматических повреждений находится в зависимости от позы и приемов, применяемых животными в бою. Травматические повреждения распределены в тех участках, которые бывают наиболее открыты и доступны для ударов со стороны противника. Носороги во время поединка сходятся голова к голове и бьются передними большими рогами. Именно такая сцена была изображен верхнепалеолитическим человеком на стене пещеры Шовэ во Франции. Естественно, что при подобной позе удары чаще всего достигают теменной области, а при боковом положении головы удары попадают в область носовых костей, глазницы и челюстного сустава.
Рисунок сражающихся шерстистых носорогов (Coelodonta antiquitatis) из пещеры Шовэ, Франция.
Относительно высокая частота встречаемости прижизненных травм у шерстистого носорога по сравнению с ныне живущими представителями семейства может объясняться быстро меняющимися климатическими условиями позднего плейстоцена, когда взрослые шерстистые носороги сталкивались с повышенным стрессом, конкурируя с другими особями популяции за ограниченные пищевые ресурсы. Это было особенно важно для выживания самок, обеспечивающих стабильность существования всего вида в целом. Вероятно, жестокие бои происходили в неблагоприятные периоды засухи, когда при нехватке воды и кормов усиливаются конкурентные отношения у животных.
О размножении шерстистого носорога практически ничего не известно. Оценки и выводы на этот счет делаются на основе сравнения с размножением современных носорогов. Беременность продолжалась приблизительно от 15 до 18 месяцев. Наличие у самки только 2 сосков говорит о том, что она рожала обычно 1, значительно реже 2 детенышей. На протяжении примерно 2 лет самка кормила детеныша молоком, а затем еще 1-2 года он пасся рядом с матерью. Затем шерстистый носорог становился самостоятельным и обзаводился собственным индивидуальным участком. Росли молодые носороги довольно медленно, и половозрелость наступала только в 10-12 лет. Детеныши носорогов в возрасте 0,5-3,5 лет довольно часто погибали. Низкий темп размножения означал, что естественное воспроизводство шерстистых носорогов было очень медленным — за 20-25 лет фертильности самка могла произвести на свет всего 6-8 детенышей.
Самка шерстистого носорога (Coelodonta antiquitatis) с детенышами разного возраста. Автор — З. Буриан.
Остатки детенышей носорогов встречаются крайне редко, гораздо реже, чем остатки мамонтят, поэтому особенно ценной является находка челюстей и костей скелетов в местонахождении Красный Яр, Кривошеинский район Томской области. Остатки принадлежат минимум 10 детенышам разного индивидуального возраста — от только родившихся до подростков 7-14 лет. Пересчет пропорциональных размеров полных трубчатых костей конечностей по сохранившимся диафизам, и в конечном итоге, размеров тела для детенышей 1-го месяца жизни дал следующие примерные размеры — высота в холке 72 см, высота в крестце около 65 см, длина тела около 120 см. Динамика увеличения длины трубчатых костей показала, что наиболее интенсивный рост происходил до 3-4 лет с достижением высоты в холке 120-135 см. После этого рост замедлялся, и к 7 годам размеры тела носорогов-подростков приближались к размерам взрослых животных. Смена молочных зубов на постоянные происходила в возрасте 3-7 лет по мере стирания молочных зубов и роста головы и нижней челюсти. Процесс развития и смены молочных зубов у шерстистого носорога совпадает с такими же данными относительно детенышей современных носорогов.
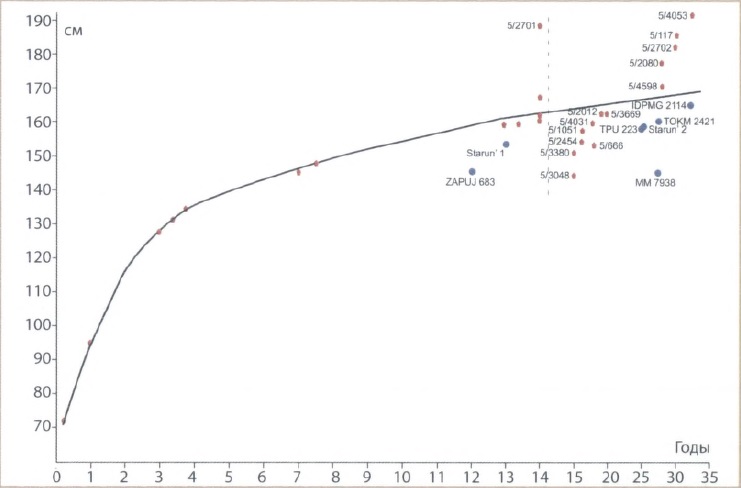
Изменение размеров тела шерстистого носорога (Coelodonta antiquitatis) в зависимости от индивидуального возраста (по материалам из местонахождения Красный Яр, Томская область, и находкам скелетов и трупов). Красным отмечена расчетная высота животных по лучевым и большим берцовым костям, синим — высота скелетов и трупов животных, найденных в Сибири и Европе. (По Шпанскому, 2021 г.)
Череп детеныша шерстистого носорога в основных пропорциях имеет сходство с таковыми ныне живущих носорогов. Отличия заключаются в выпрямленности профильной линии черепа от вершины затылочной кости до конца носовых костей. Угол наклона теменной плоскости к лобной поверхности равен нулю. Плоскость затылка у детенышей Coelodonta antiquitatis составляла к базальной линии основания черепа угол 90 °. Скуловые дуги выпрямлены. В черепах ранней ювенильной стадии прослеживаются признаки рода в удлиненных пропорциях всего черепа, которые формировались позднее за счет роста костей лицевого отдела черепа. Закрепленные на ранней стадии онтогенеза морфологические особенности черепа шерстистого носорога могут свидетельствовать о том, что эти признаки очень рано были сформированы в филогенезе.
Череп молодого шерстистого носорога (Coelodonta antiquitatis) в возрасте 3-5 лет из Башкирии. Заметны несросшиеся черепные швы и неокостеневшая носовая перегородка.
Двухслойная носовая перегородка, которая приблизительно до 5-летнего возраста оставалась еще хрящевой, после 10 лет прочно срасталась с носовыми, лобными, предчелюстными, верхнечелюстными костями и сошником, образовывая единый монолит лицевого черепа. Примерно от 6 лет образуются первые центры окостенения в передней (внешней) части перегородки, на конце носовых костей. Естественно, что окостенение носовой перегородки начинается там, где возникают наибольшие нагрузки на носовые кости и возрастает раздражающее воздействие на эпидермальные ткани в области морды животного, когда оно при добывании корма терлось мордой и передним рогом о землю. В поздней ювенильной стадии, совпадающей с периодом активного созревания (в момент прорезывания М3 и его функционирования), как правило, происходило быстрое окостенение носовой перегородки и срастание ее с костями лицевого черепа.
В коллекциях встречаются образцы черепов шерстистого носорога, у которых на одинаковой стадии развития Р4 костная носовая перегородка была в разной степени окостеневшая. Как правило, в черепах с территории Восточно-Европейской равнины носовая перегородка менее сформирована, чем в черепах, которые происходят с территории Северо-Восточной Сибири. Следовательно можно предположить, что популяция шерстистого носорога с территории Восточной Европы существовала в более мягких климатических условиях, в отличие от популяции носорогов обитавших на территории Северной Азии.
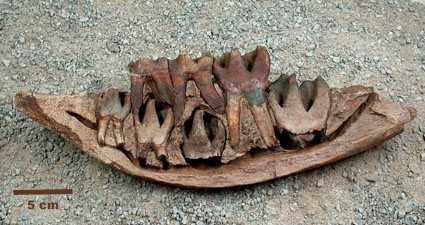
Фрагмент нижней челюсти молодой особи шерстистого носорога (Coelodonta antiquitatis) с молочными и постоянными зубами.
На взрослой стадии онтогенетического развития, соответствующей времени стирания М3 примерно наполовину, носовая перегородка полностью окостеневает и срастается с костями лицевого черепа. В дальнейшем идет только упрочнение и разрастание внешних частей и краевых ее листков, которые полностью срастаются и подпирают носовые кости черепа. На поздней ювенильной стадии онтогенеза (на стадии прорезывания Р4) заканчивается в основных чертах формирование мозгового отдела черепа. С прорезыванием и функционированием последнего коренного М3 устанавливаются размеры черепа. После этого, вероятно, череп уже медленно растет в длину. Идет лишь увеличение общей массивности костей и рост их в широтном направлении.
Черепа молодых животных имеют суженные носовые кости в концевой их части. Это связано с тем, что передний рог тоже не достиг своих окончательных размеров и продолжал рост еще долго в течение жизни животного. У взрослой особи (когда М3 стерт примерно наполовину) носовые кости по форме приближаются к удлиненному прямоугольнику. С возрастом усиливается общая скульптурированность костей лицевого и мозгового отделов черепа. Особенно хорошо она становится развитой в области прикрепления рогов (экзостозы на носовых и лобных костях), и в области теменных и височных костей, в местах прикрепления жевательных мышц. Также с возрастом уплощаются и становятся более скульптурированными скуловые дуги. Кости лицевого черепа увеличиваются в ширину. Это связано с тем, что снашивание Р2-4 происходит быстрее. Постепенно нагрузка с передней части челюстей перемещается назад, к М1-М3, что способствует росту черепа в широтном направлении. Носовой рог к зрелому возрасту (25-35 лет) часто был больше метра в длину.
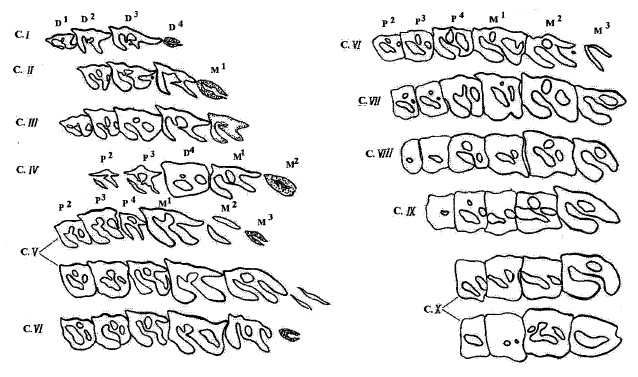
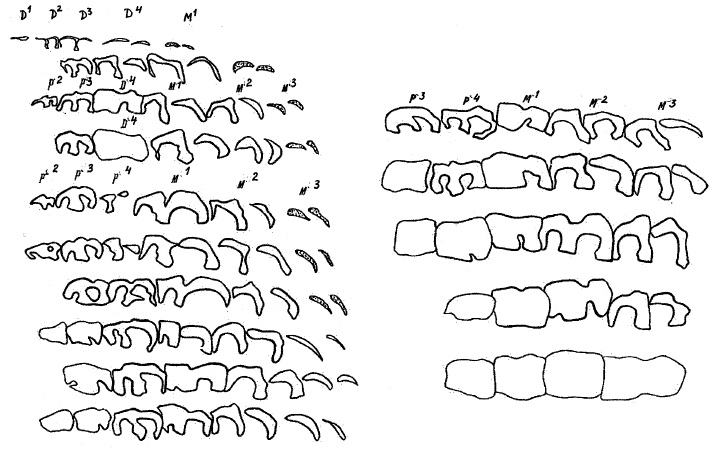
Возрастные стадии стирания верхних щечных зубов шерстистого носорога (Coelodonta antiquitatis).
Возрастные стадии стирания нижних щечных зубов шерстистого носорога (Coelodonta antiquitatis).
Изучение большого количества носорожьих черепов и отдельных челюстей свидетельствует, что темп износа зубов шерстистого носорога почти в точности совпадал с износом зубов современных африканских видов. Исследователи на этом основании сделали вывод об идентичности возрастных стадий шерстистого и современных носорогов и, следовательно, максимальной продолжительности жизни — 40-45 лет. Эти данные подтверждаются и при подсчете максимального числа микроскопических сезонных слоев нарастания дентина на срезах верхних коренных зубов у старых особей белого и индийского носорогов.