Строение и функции
Парацератерии несомненно являлись одними из крупнейших когда-либо существовавших наземных млекопитающих, однако их точный размер неясен из-за значительной фрагментарности материала. Расчеты базируются главным образом на окаменелостях Paraceratherium transouralicum, поскольку скелет этого вида известен наиболее полно. Оценки основаны на измерениях зубов, костей черепа и конечностей, но они принадлежат особям разной величины, поэтому все скелетные реконструкции парацератериев в значительной мере условны. Кроме того, у различных их видов (Paraceratherium transouralicum, Paraceratherium prohorovi, Paraceratherium bugtiense) отмечена исключительная индивидуальная изменчивость в размерах: на сериях их костей из одного местонахождения длина самых меньших образцов составляет 2/3, а иногда даже 1/2 длины самых больших.
Согласно расчетам, наиболее крупные представители рода достигали 8,5 м в длину и 5,5 м в холке (по более скромной оценке — 7,5 м и 4,8 м соответственно). Самый большой относительно полный череп Paraceratherium transouralicum (образец AMNH № 18650) имеет 130 см в длину, а его ширина в области скуловых дуг и мозгового отдела составляет соответственно 61 и 38 см; некоторые обнаруженные фрагменты предполагают наличие еще более крупных черепов. Расчетная длина шеи составляет 2-2,5 м. Вполне вероятно, что максимальной величины достигали только взрослые самцы. Их масса оценивается в 15-20 т, тогда как самки весили 10-15 т. Лишь некоторые из самых больших хобтных, такие как Deinotherium giganteum, Palaeoloxodon namadicus и Mammuthus trogontherii, могли достигать подобных размеров. Более мелкие виды парацератериев были порядка 6,2 м в длину, 3,5 м в холке и имели массу 7-9 т.

Парацератерий (Paraceratherium transouralicum) в сравнении с современным саванным слоном (Loxodonta africana) и человеком. Автор — Р. Учитель.
Несмотря на значительные размеры, череп по сравнению с телом кажется небольшим. Он длинный и узкий, безрогий. Особенно вытянута задняя часть черепа: длина от заднего края затылочного мыщелка до заднего края засуставного отростка составляет больше 1/4 длины от последней точки до переднего края Р2. Передний отдел верхней челюсти расширен в обоих направления. Предчелюстные кости загнуты вниз. Носовые кости длинные, без поддерживающей их носовой перегородки. Носовая вырезка глубокая. Лобные кости гладкие. Мозговой отдел короткий (у Paraceratherium transouralicum он составляет 8 % от общей длины черепа, тогда как у современного Rhinoceros unicornis — 17,7 %), узкий и низкий. Порой имеется слабо выраженный сагиттальный гребень. 3асуставный и заслуховой отростки не соприкасаются. Затылок не такой широкий, как у современных носорогов. Затылочные мыщелки очень массивные. 3атылочные мыщелки очень крупные, длина их большой оси составляет около половины высоты затылка от нижнего края мыщелков. Между затылочными мыщелками имеется глубокая ямка. Нижняя челюсть длинная и узкая, но довольно массивная. Ее передний отдел отогнут вниз. Нижнечелюстной симфиз массивный и длинный, достигает уровня р3.


Слепок сборного скелета парацератерия (Paraceratherium transouralicum). Палеонтологический музей им. Ю.А. Орлова РАН, Москва.
Зубная формула: I1/i2-1, С1-0/с0, P4-3/р3, М3/m3. III-й, а иногда и II-й нижние резцы, нижний клык, нижний I-й предкоренной, а порой и верхний I-й предкоренной утрачены. Два верхних резца (вероятнее всего, I2) очень крупные (длиной до 7 см), конические, направленные вниз и/или немного вперед, либо рудиментарные; два нижних (по-видимому, i1) — очень крупные, от округлых до овальных в сечении, направленные прямо вперед и вплотную сближенные; иногда, кроме них, имеется вторая пара (i2) рудиментарных. Среди известных носорогов такое расположение резцов уникально для рода Paraceratherium и близкого к нему Urtinotherium. Вероятно, у самцов резцы были крупнее, чем у самок. Верхние клыки рудиментарные или отсутствуют, нижних клыков нет. В обоих челюстях передние зубы отделены от щечных широкими диастемами.
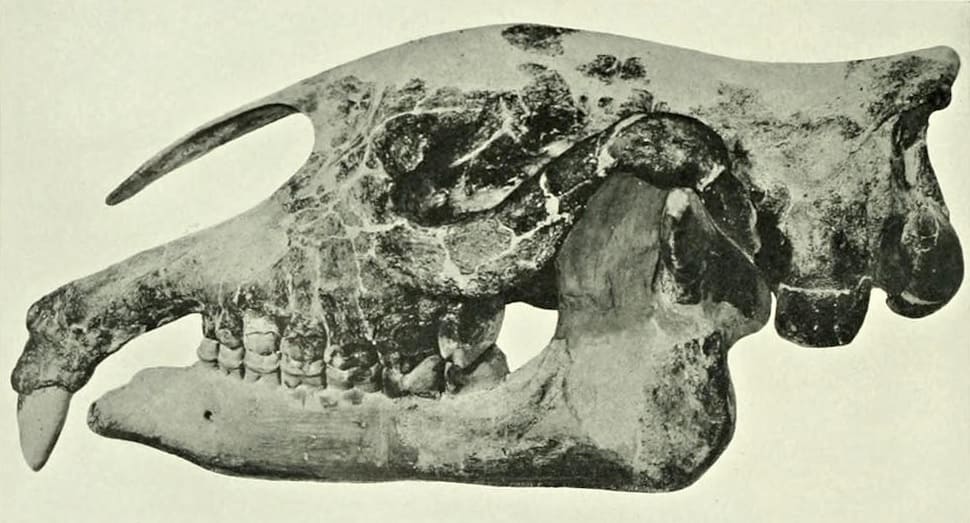
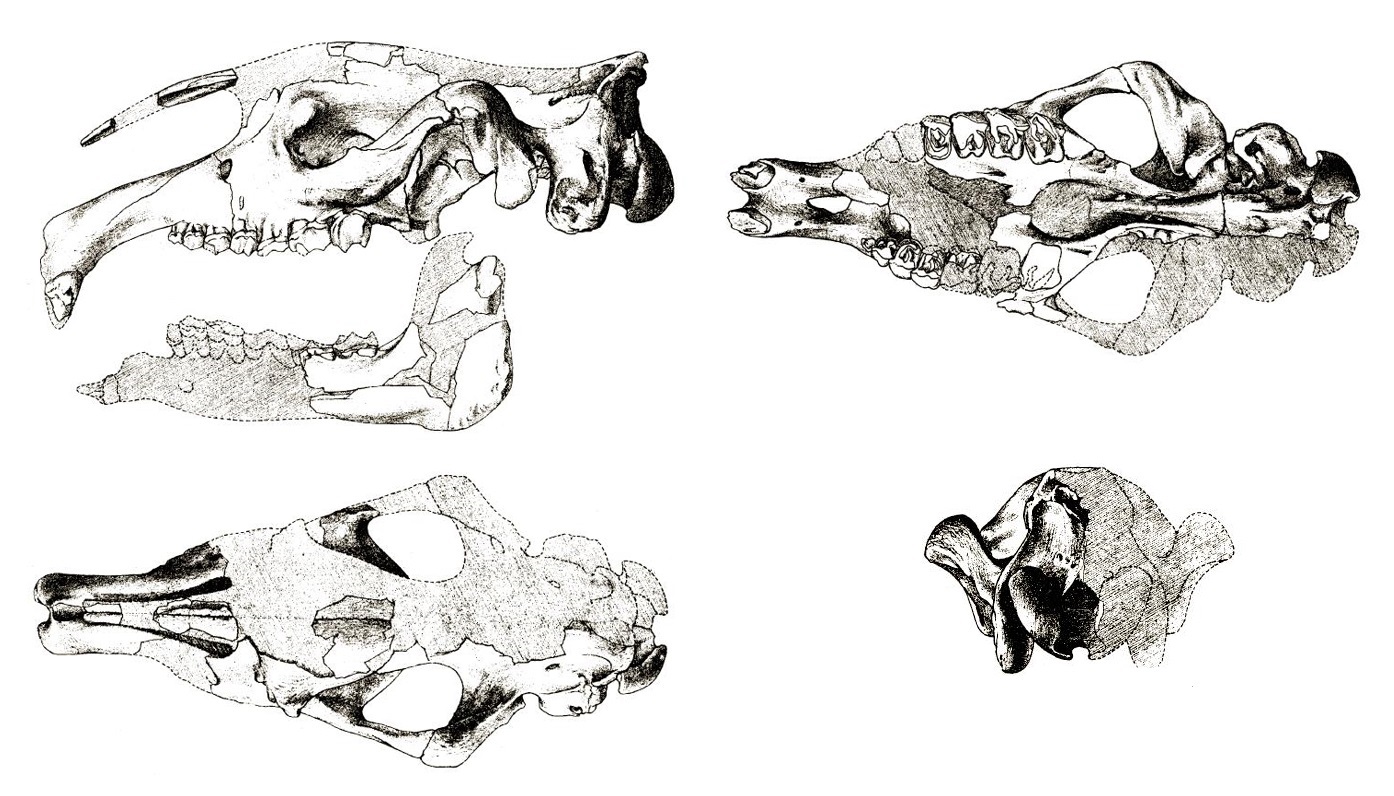
Частично реконструированный череп с нижней челюстью парацератерия (Paraceratherium transouralicum) (образец AMNH № 18650) и их прорисовки.
Передние предкоренные зубы сильно уменьшены, в то время как задние размером почти с последующие коренные. Длина Р2-Р4 значительно превосходит половину длины М1-М3. Зачастую последний коренной имеет самые большие размеры, достигая в длину более 10 см, а общая длина ряда щечных зубов составляет 40 см и более. Но в целом щечные зубы по сравнению с размерами животных очень невелики. Коронки щечных зубов умеренно высокие с простым для носорогообразных рисунком жевательной поверхности, слабо развитыми и мало изогнутыми дополнительными эмалевыми складками. При этом покрывающий зубы слой эмали очень мощный — до 4 мм. Верхние предкоренные слабомоляризованы, особенно Р3 и Р4: они не имеют цельного до вершины металофа и раздельных до основания прото- и гипокона; степень моляризации убывает от Р2 к Р4. Верхние и нижние коренные со слабо скошенными поперечными гребнями. M3 с очень слабым рудиментом заметалофной части эктолофа и без задней долинки. Нижние коренные с хорошо развитой передней (поперечной) ветвью металофида и с наружной (продольной) ветвью гиполофида, присоединяющейся к задней (поперечной) ветви металофида. Переднее и заднее полулуния нижних коренных разделены снаружи глубокой бороздой. Гребни верхних коренных образуют П-образный узор, за исключением V-образного M3. Предкоренные формируют π-образный узор лишь частично. Нижние коренные с L-образными гребнями.
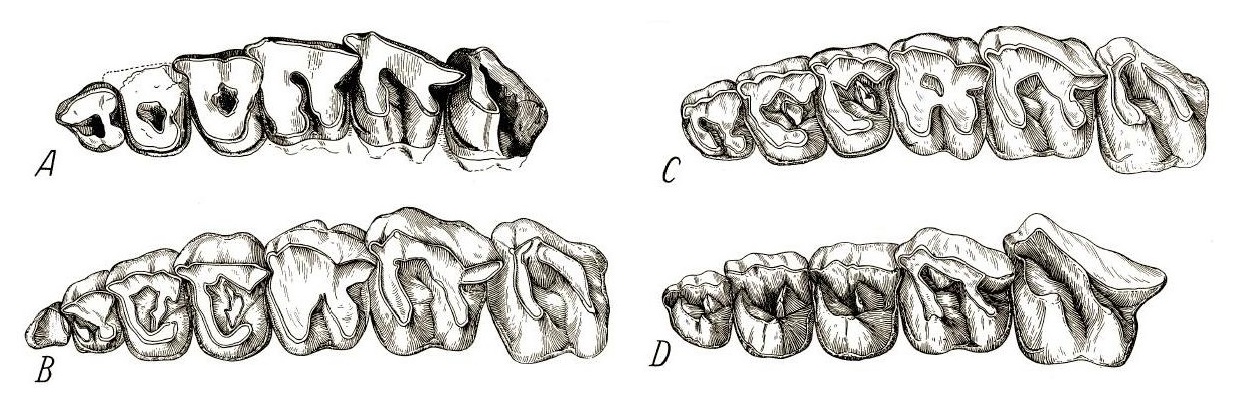
Прорисовки жевательной поверхности верхнего правого ряда щечных зубов парацератериев: А — Paraceratherium bugtiense; B — "Paraceratherium grangeri"; С — взрослая особь Paraceratherium transouralicum; D — молодая особь Paraceratherium transouralicum.
Полные серии позвонков и ребер не обнаружены. Длина шейного отдела позвоночника в 1,5-2 раза превышает длину черепа. Атлант длинный и узкий, его длина составляет от 1/2 до 3/4 ширины. Прочие шейные позвонки также удлиненные, с каналами для прохождения связок и мышц, необходимых для поддержания головы. Внутри тел позвонков каналы расширяются в большие полости, облегчающие вес шеи. Тела остальных позвонков очень широкие, с массивными сочленовными отростками и обширными полостями для мышц, сухожилий, связок и нервов. Остистые отростки туловищных позвонков высокие, особенно развитые в передней части грудного отдела, где к ним прикреплялись шейные мышцы и затылочные связки. Последний спинной позвонок сращен с крестцом, что характерно для продвинутых носорогов. Как и у динозавров-завропод, в телах предкрестцовых позвонков имеются округлые выемки (вплоть до сквозных отверстий), которые, вероятно, помогали облегчить скелет. Ребра сходны с таковыми современных носорогов, но образуемая ими грудная клетка на фоне большого тела и длинных конечностей кажется не столь массивной.

I-й шейный позвонок Paraceratherium.
Конечности очень длинные, колоннообразные, почти не согнутые в суставах, напоминающие таковые слонов и динозавров-завропод. Кости плеча, бедра, предплечья, голени, пясти и плюсны сильно удлинены (длина плечевой кости может достигать 99 см, лучевой — 120 см, бедренной — 150 см, большой берцовой — 90 см, центральных метаподиев — 63 см). Поскольку ноги занимали выпрямленное положение, это уменьшало потребность в их развитой мускулатуре.
Головка плечевой кости направлена почти прямо вверх, большой бугор, дельтовидный гребень и дельтовидная бугристость развиты слабо. Характерные черты строения плечевой кости легко объяснимы колоннообразностью строения передней конечности. Так, форма головки явно указывает на слабый размах движений в плечевом суставе; слабость нижних надмыщелков — на умеренное развитие сгибателей и разгибателей кисти, естественно умеренно обременяемых при выпрямленных конечностях; слабый рельеф суставных поверхностей локтевого сустава объясним слабым размахом сгибательно-разгибательных движений и т. д.
Локтевой отросток локтевой кости очень короткий, полулунная вырезка мелкая. Лучевая кость частично опирается на трехгранную кость запястья. Головка бедренной кости также обращена почти прямо вверх; большой вертлуг расположен ниже нее, малый вертлуг отсутствует, III-й вертлуг очень небольшой. Внутренняя лодыжка на большой берцовой кости не развита.

Пястные кости 3 особей парацератерия (Paraceratherium transouralicum) в сравнении с той же костью современного белого носорога (Ceratotherium simum) (крайняя слева).
В кисти и стопе по 3 функционирующих пальца. Их элементы, связанные с I-м и V-м пальцами, крайне рудиментарны, иногда отсутствуют. Средний палец развит сильнее боковых, сдвинутых относительно него назад. На головку локтевой кости опирается только полулунная I-го ряда, которая внизу спереди вклинивается между большой и крючковатой II-го ряда. Верхние и нижние суставные поверхности костей запястья и заплюсны уплощенные, со слабо выработанным рельефом, с приспособлениями, препятствующими верхнему сгибанию. На головку локтевой кости опирается только полулунная I-го ряда, которая внизу спереди вклинивается между большой и крючковатой II-го ряда. Блок таранной кости мелкий, гребни закругленные.
Пястные и плюсневые кости сильно удлиненные, боковые (II-я и IV-я) сужены по сравнению со средней (III-й) и несколько отодвинуты назад. Фаланги средних пальцев очень короткие и широкие, их верхние и нижние поверхности уплощенные. Фаланги боковых пальцев сплющенные в боковом направлении, их широкие поверхности обращены в стороны, а на их суставных поверхностях слабо развиты направляющие гребни и борозды. Концевые фаланги большие, с длинными "ветвями". По отношению к оси конечности боковые пальцы повернуты таким образом, что поверхность, обращенная у настоящих носорогов вперед, здесь направлена почти прямо наружу, а задняя — внутрь; "ветви", соответствующие наружной и внутренней, получают направление назад и вперед. Хотя конечности функционально трехпалые, они отчетливо демонстрируют тенденцию к образованию однопалости.
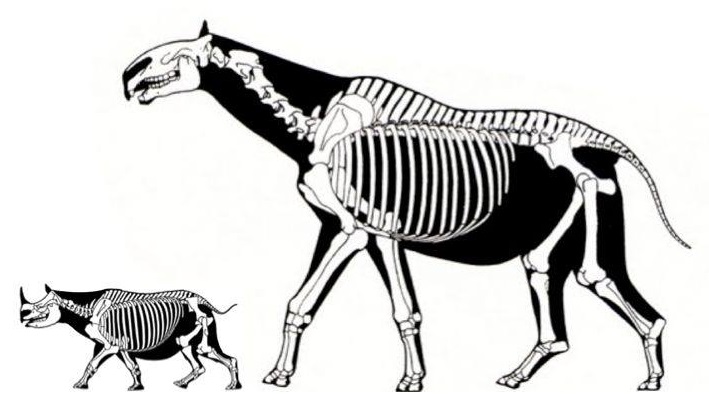
Прорисовка реконструированного скелета парацератерия (Paraceratherium transouralicum) в сравнении со скелетом современного черного носорога (Diceros bicornis).