Родственные связи
А.А. Борисяк (1939 г.) отнес роды Indricotherium, Baluchitherium, Paraceratherium и Aralotherium к семейству Indricotheriidae, указывая на их отличие от Hyracodontidae в строении М3. На Западе гигантских носорогов относили к Rhinocerotidae вплоть до 1967 г., когда они были переведены в состав Hyracodontidae на основании особенностей строения передних зубов. Rhinocerotidae и Hyracodontidae тогда рассматривались в качестве сестринских групп в рамках Rhinocerotoidea. У Rhinocerotidae имеется пара пластинчатых верхних резцов, тогда как их нижние резцы долотовидные и острые. С другой стороны, у Hyracodontidae передний зубной ряд относительно изменчив. Гигантские носороги обладали долотообразными верхними резцами. Л. Радинский отнес их именно к этому семейству в качестве подсемейства Indricotheriinae. Решетов (1991 г.) полагал, что Indricotherium относится к отдельному семейству Indricotheriidae, связанному с Hyracodontidae лишь общим происхождением. В 2010-х гг., из-за высокой изменчивости Hyracodontidae, независимость семейства Indricotheriidae была признана западными и китайскими специалистами.
По мнению В.И. Громовой (1959 г.), Indricotherium transouralicum еще сохраняет некоторые архаические признаки в строении зубов — изолированный энтоконид нижних предкоренных, немоляризованные верхние предкоренные с не вполне выработанным металофом, ясно моделированные паракон и метакон верхних коренных и некоторые другие. И в костях конечностей этого вида, несмотря на высокую общую специализацию, имеются отголоски общего происхождения с настоящими носорогами (Rhinocerotidae), очевидно, унаследованные последними, как и гигантскими носорогами, от раннеэоценовых общих предков.
Paraceratherium bugtiense представляет собой более поздний и прогрессивный таксон. Есть, однако, одна черта строения верхних предкоренных, которая у Paraceratherium bugtiense более архаична, чем у Indricotherium transouralicum. Это меньшее выступание внутрь гипокона чем протокона на верхних предкоренных и в связи с этим более асимметричное, треугольное очертание их коронок. У Indricotherium transouralicum гипокон предкоренных выступает внутрь так же далеко, как протокон, и очертание коронки почти трапециевидное. В этом отношении предкоренные Indricotherium transouralicum более моляризованы и, следовательно, более прогрессивны, чем у Paraceratherium bugtiense, хотя в других отношениях те же зубы первого архаичнее, чем второго (более слабое разделение внутренних бугров, меньшая выработанность металофа, более ясная моделировка наружных бугров и т. д.). Эту прогрессивную черту Indricotherium transouralicum отметил и А. Борисяк (1939 г.), что заставило его исключить этот род из прямых предков Paraceratherium bugtiense. Тем не менее, В. Громова (1959 г.) и Л. Радинский (1967 г.) считают его предковой формой Paraceratherium bugtiense.
М.Д. Бирюков (1953 г.) полагает, что Pristinotherium brevicervicale представляет собой начальную своеобразную ветвь в развитии гигантских носорогов, при этом с Indricotherium transouralicum он существовал некоторое время одновременно. Несколько позже, с изменением климата и условий среды, от родоначальной ветви короткошеих гиганских носорогов выделились новые. Очевидно, наибольшего своего расцвета Pristinotherium brevicervicale мог достигнуть в эпоху сильного сокращения Чеганского морокого бассейна, которое имело место в начале олигоцена. После же отступления к югу морских вод и осушения Тургайского пролива, на бывшей территории Чеганского бассейна, как считается, образовались большие пойменные, озерные и болотные лесистые пространства, среди которых и жил короткошеий гигантский носорог.
В конце раннего олигоцена отмечается резкое сокращение лесных пространств и распространение более сухих ландшафтов саванного характера. В этих экологических условиях ряд жизненно важных особенностей организма Pristinotherium brevicervicale оказался несовместимым с возникшими новыми условиями обитания. Надо полагать, это и стало главной причиной его вымирания. В этих условиях шло развитие новых представителей семейства гигантских носорогов. Indricotherium transouralicum, вероятно, начал свое формирование лишь в возникшей новой обстановке саванн (например, на территории Монголии, где леса стали редеть раньше) и существовал дольше Pristinotherium brevicervicale.
Происхождение самого таксона Indricotherium transouralicum также представляется туманно. А.А. Борисяк (1918 г.), а вслед за ним В. Гренджер и В. Грегори (1936 г.) сравнивают остатки Indricotherium transouralicum с родом азиатских гиракодонтов — Allacerops, выдвигая его на место вероятного предка. Л. Радинский (1967 г.) в качестве предка называет другой род гигантских носорогов из раннего олигоцена Китая — Urtinotherium, который в свою очередь происходит от позднеэоценового рода Forstercooperia, известного из Азии и Северной Америки, также вероятного предка Allacerops (в настоящее время Allacerops признается младшим синонимом рода Eggysodon, выделяемого в семейство Eggysodontidae).
Проблемы реконструкции
Реконструкция скелета гигантских носорогов встречает значительные затруднения. Ни одного достаточно полного скелета или хотя бы целых конечностей неизвестно, точнее говоря, не описано (звестны только целые кисть и стопа от запястья и заплюсны включительно из Монголии и стопа оттуда же, из Татал-Гола). Известны не все шейные позвонки, что затрудняет определение длины шеи. Величина индивидуальной изменчивости огромна, так что при подборе скелета может быть сильно искажено соотношение в длине разных его частей.

Первая реконструкция скелета и внешнего облика "Indricotherium asiaticum", выполненная под руководством А. Борисяка в 1914 г.
Первая реконструкция скелета и внешнего облика игантского носорога была выполнена под руководством А.А. Борисяка (1914 г.), в распоряжении которого был тургайский скелет Indricotherium asiaticum без черепа. Последний был восстановлен исходя из общих пропорций носорогов, в результате чего получился на удивление громадным. В дальнейшем Борисяк (1924 г.) поправил свою реконструкцию, присовокупив к ней череп Baluchitheium grangeri, найденный в Монголии. На рисунках Борисяка шея и конечности животного значительно длиннее, чем на более поздней реконструкции Грейнджера и Грегори, а конечности также длиннее, чем на реконструкциях Осборна. Разница в длине ног зависит частично от различной длины их костей по отношению к длине туловища, а отчасти от разной согнутости в суставах.
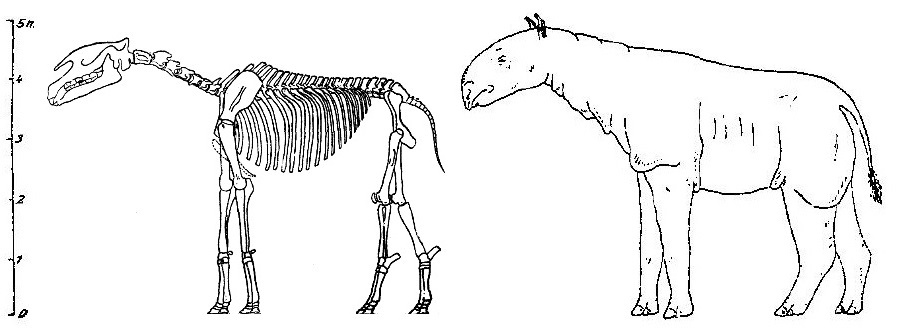
Вторая реконструкция скелета и внешнего облика "Indricotherium asiaticum", выполненная под руководством А. Борисяка в 1924 г.
Первая реконструкция Г.Ф. Осборна (1923 г.) была основана лишь на нескольких находках из Монголии и поэтому напоминала по своим пропорциям настоящих носорогов, со слишком грузным туловищем, короткой шеей и конечностями. В том же году под руководством Осборна была выполнена более достоверная реконструкция, базирующаяся на достаточно обширном материале из Монголии и Белуджистана. Парацератерий здесь представлен более легко сложенным, с длинной шеей и конечностями. Однако и на новой реконструкции пропорции ног не до конца отражают исключительную для млекопитающих длину их костей, и не только проксимальных, но и в метаподиальном отделе. Ноги гигантских носорогов, во всяком случае, должны были быть выше, чем у слонов, у которых удлинены только проксимальные отделы конечностей.
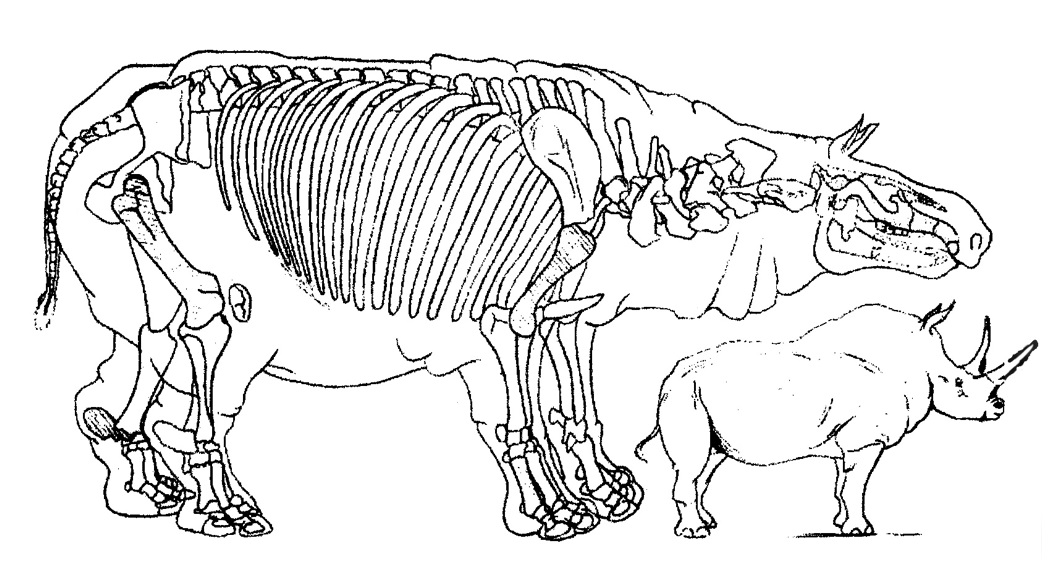

Две реконструкции скелета и телесных очертаний "Baluchitheium grangeri" в сравнении с белым (Ceratotherium simum) и индийским (Rhinoceros unicornis) носорогами, выполненные под руководством Г. Осборна в 1923 г.
Весьма исчерпывающая реконструкция парацератерия, до сих пор в значительной мере определяющая его образ, была выполнена под руководством У. Грейнджера и У.К. Грегори (1935 г.). Авторы приложили много усилий для достоверного изображений пропорций. Они разделили все наличные кости Baluchitheium grangeri по величине на 4 категории, поскольку они принадлежат целому ряду особей различной величины, затем вычислили коэффициенты перевода длины костей из одной категории в другую и привели размеры всех костей скелета к самой крупной категории; на этих размерах и был восстановлен скелет на рисунке. Естественно, что и на этом пути возможны значительные искажения пропорций, особенно, если учесть большие пробелы в позвонках и ребрах. Так, чрезмерно укороченной выглядит шея животного.

Реконструкция скелета и телесных очертаний "Baluchitheium grangeri" в сравнении с человеком, выполненная под руководством У. Грейнджера и У. Грегори в 1935 г.
Очень близко к реконструкции А.А. Борисяка смонтирован сборный скелет Paraceratherium transouralicum в Палеонтологическом музее в Москве. Н.А. Яншинов под руководством палеозоолога и художника К.К. Флерова сделал прорисовку этого скелета (с небольшими изменениями), которую затем опубликовала В.И. Громова (1959 г.). Являясь заведующим музея, Флеров требовал, чтобы скелет сделали как можно выше и он являлся самым заметным экспонатом. Очертания нескольких отсутствовавших шейных позвонков были реконструированы неверно. Кроме того, Флеров сделал несколько художественных реконструкций животного. На них парацератерий выглядит как гигантская лошадь с жирафьей шеей и сравнительно небольшой головой. Однако на самом деле зверь выглядел несколько иначе — было приземистее, обладал более короткой шеей.
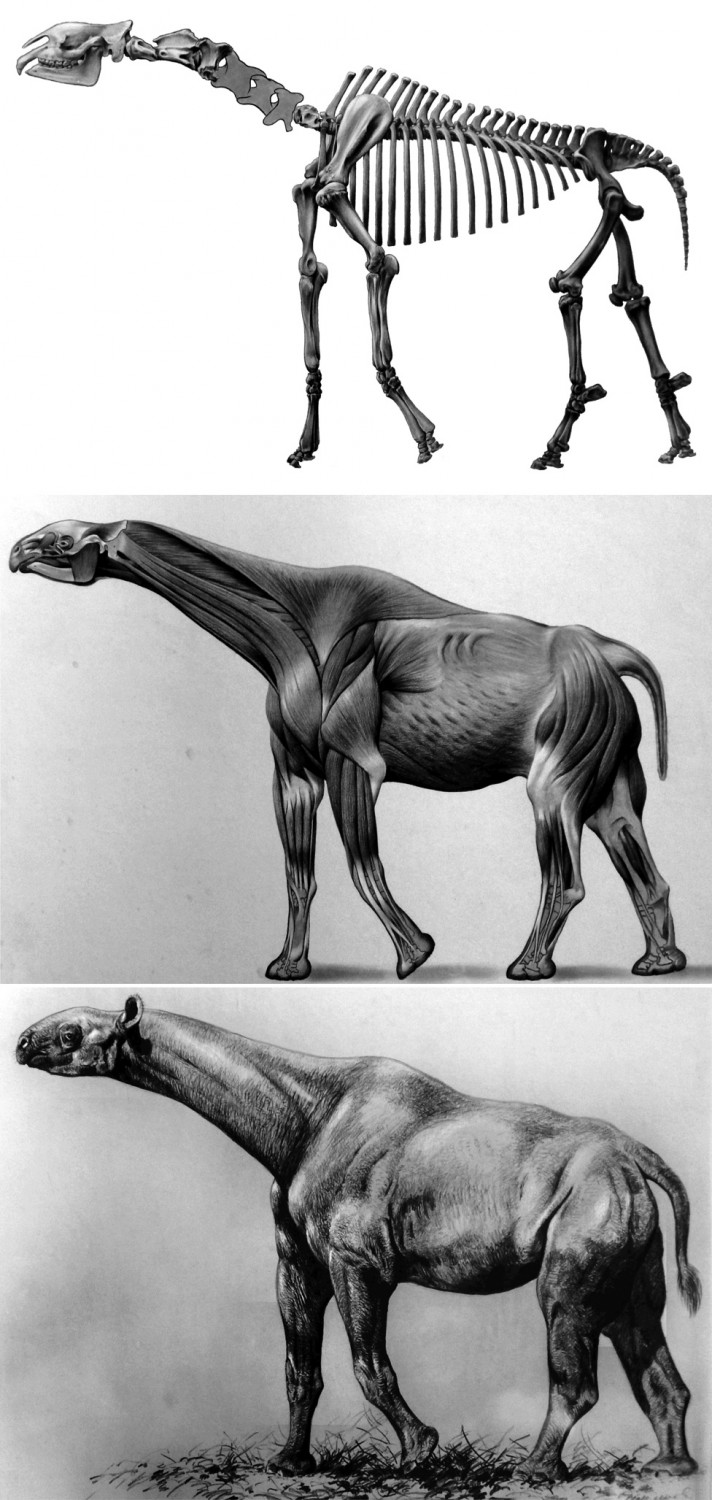
Поэтапная реконструкция внешнего облика "Indricotherium transouralicum". Рисунки скелета и мускулатуры выполнены Н. Яншиновым под руководством К. Флерова, восстановившего облик зверя в 1950 г.
Сложный вопрос представляет собой верхняя линия профиля черепа Paraceratherium transouralicum. На многих реконструкциях изображена сильно выпуклая линия черепного профиля. Это представление коренится в реконструкции сильно раскрошенного черепа монгольского "Baluchitheium grangeri", выполненной Американским музеем естественной истории, где череп представлен горболобым и горбоносым. Однако нет полной уверенности в правильности этой реконструкции. Единственный кусок кости на лобной поверхности указанного черепа очень маленький. Неизвестно, насколько правильный придан ему наклон на месте его контакта с вертикальной пластинкой лобной кости; если наклон сделать другим, то и выпуклость лба сильно уменьшается. Американские авторы ничего не пишут о способе реконструкции лба и вопрос о его профиле не достаточно ясен (в своей реконструкции всего скелета Грейнджер и Грегори дают значителыю менее выпуклый профиль). Более-менее полных черепов Paraceratherium transouralicum больше не известно. У других видов парацератериев лоб, во всяком случае, выпукл слабо или даже слегка вогнут.

Парацератерии (Paraceratherium transouralicum) в естественном окружении. Автор — К. Флеров, 1959 г.
Строение скелета парацератериев указывает на их питание растительностью верхнего древесного яруса. Именно этим объясняются их гигантский рост, высокие ноги и длинная шея, сходные с таковыми представителей семейства жирафовых (Giraffidae). Эти особенности, в свою очередь, определили ряд других морфологических приспособлений.
Расположение сильно наклонных передних поверхностей тел шейных позвонков заметно выше задних указывает на наклонное в норме положение шеи с подъемом вперед. Такое положение дано в реконструкциях Борисяка и Осборна; напротив, по Грейнджеру и Грегори, шея реконструированного скелета направлена почти горизонтально. Относительно небольшая голова, большие выемки и сквозные отверстия в телах шейных позвонков облегчали напряжение мышц-разгибателей шеи, которое, при длинной шее животных, должно было быть значительным (большое плечо сопротивления рычага). Сиьно напряжены были и связки, поддерживавшие шею и голову, о чем свидетельствует глубокая срединная ямка на затылке — место прикрепления выйной связки, которая автоматически удерживает череп. Значительно расширенные затылочные мыщелки подтверждают наличие хорошо развитых шейных мышц, позволяющих нагибать голову вниз во время кормления.

Возможное поведение парацератерия (Paraceratherium transouralicum) на водопое и рисунок его скелета с максимально опущенной вниз головой.
Длинная, подвижная, особенно в вертикальном направлении, шея парацератериев, большей частью приподнятая кверху, хорошо увязывается с предполагаемым способом их питания и с необходимостью иногда (при питье) сильно опускать голову вниз. Вопрос о том, могли ли они пить, не раздвигая в стороны ноги, подобно современному жирафу, пока не может быть решен, так как до сих пор не найдено или, по крайней мере, не описано не только целого скелета, но даже полного шейного отдела позвоночника.
Согласно другой точке зрения, расположение суставных поверхностей шейных позвонков свидетельствует, что голова обычно занимала горизонтальное положение, но могла поворачиваться из сторогны в сторону. Кроме того, первые два шейных позвонка благодаря своему строению ограничивали движения в вертикальной плоскости. Вследствие этого постоянное положение головы с изогнутой вверх шеей, подобное таковому у лошади или жирафа, была невозможным.

Реконструкция внешнего облика "Baluchitheium grangeri". Автор — Ч. Найт.
Огромное, тяжелое туловище при длинных сегментах конечностей делало невозможным сколько-нибудь значительное сгибание их в суставах: как поддержка при стоянии, так и разгибание при передвижении вперед требовало бы очень сильного мышечного напряжения. Поэтому выработались прямые, колоннообразные конечности, при которых давление веса тела принимают на себя почти исключительно сами кости, а не мышцы, разгибающие суставы. Сходное строение конечностей, и по той же причине, наблюдается в далеко отстоящей ветви млекопитающих — у слонов.
Слабое сгибание ног в суставах, т. е. сильно открытые углы между их отделами, имеет следствием также незначительное увеличение этих углов при разгибании, которое происходит при передвижении тела впред, а, следовательно, и незначительное перемещение тела вперед с каждым шагом животного. Поэтому парацератерии следует считать малоподвижными животными. Отчасти это малое изменение взаимного положения отделов ног при каждом шаге компенсировалось значительной длиною конечностей. При колоннообразном строении конечностей необходимо было ограничение движений в их суставах, и притом в направлении не только сгибания и разгибания, но и приведения и отведения, так как при огромном весе животного каждое отклонение любого сегмента конечностей от прямого, отвесного положения требует значительных усилий для его восстановления.

Пластическая реконструкция внешнего облика "Baluchitheium grangeri".
Выпрямленность конечностей и ограничение возможных движений в их суставах запечатлелись на всех без исключения костях гигантских носорогов. Это хорошо видно при сравнении элементов их конечностей с таковыми носорогов настоящих — значительно более коротконогих животных, с более согнутыми в суставах ногами, большей частью довольно хороших бегунов. Для парацератериев характерны слабо выпуклые и обращенные почти прямо вверх головки плечевой и бедренной костей и почти прямо вниз — нижние мыщелки бедренной кости; короткий локтевой отросток локтевой кости (короткое плечо силы рычага), небольшой клювовидный отросток этой кости и мелкая ямка для него на плечевой; мелкий блок таранной кости и мелкая впадина для него на большой берцовой кости; слабый рельеф суставных поверхностей костей кисти и стопы, особенно верхних поверхностей метаподий; имеются тормозные приспособления для предупреждения верхне-нижнего сгибания в разных этажах сложного запястного сустава; слабое развитие сгибательной и разгибательной мускулатуры, что особенно ясно на плече, бедре и голени; слабый связочный аппарат на волярной и плаптарной стороне кисти и стопы (эти связки при выпрямленных конечностях, естественно, менее обременяются) и многие другие признаки.

Пластическая реконструкция внешнего облика "Baluchitheium grangeri"; справа — реконструкция внешнего облика Hyracodon nebrascensis.
В связи с необходимостью значительного увеличения роста, у гигантских носорогов сильно удлинялись кости конечностей, особенно их проксимальных отделов — плеча и бедра. При этом во избежание чрезмерного увеличения общего веса тела эти кости сравнительно мало увеличились в толщину, т. е. стали относительно тоньше, насколько это допускала необходимая для поддержания огромного тела крепость костей конечностей. Характерно, что, в отличие от слонов, у которых удлинены только проксимальные отделы ног, у гигантских носорогов сильно удлинены также метаподии. Как правило, длинные метаподии — показатель способности к быстрому бегу. В данном случае, у медлительных животных с колоннообразными конечностями, значение удлинения, очевидно, другое — увеличение роста с целью добычи листьев и ветвей с высоких деревьев. Вероятно, при необходимости максимального (большего, чем у слонов) удлинения конечностей, дальнейшее удлинение плеча и бедра было бы механически нецелесообразно. Кроме того, индрикотериевые, в отличие от хоботных, произошли от уже довольно быстрых бегунов с удлиненными пястью и плюсной — Forstercooperiidae или Hyracodontidae.

Реконструкция внешнего облика "Indricotherium transouralicum". Автор — З. Буриан, 1955 г.
У предков Indricotheriidae уже началось сокращение в ширину боковых пальцев и увеличение среднего. У гигантских носорогов, при увеличении роста и при связанном с этим ослаблении свободы движений конечностей, процесс сокращения боковых пальцев продолжался. I-й и V-й пальцы исчезли совсем (сохранился только рудимент V-й пястной кости), а ІІ-й и IV-й пальцы стали значительно тоньше. Удлинение метаподиальных отделов вместе с сокращением боковых пальцев привело к своеобразной "монодактилизации" конечностей парацератериев. Однако направление приспособления конечностей у них резко отличалось от того, которое имело место в семействе лошадиных (Equidae) — группе, где развитие однопалости достигло высшего предела. У лошадиных в процессе эволюции удлинялись не только метаподии, но и фаланги пальцев, которые все больше приподнимались над землей, так что вся конечность уже очень рано начинает опираться на подошвенную поверхность последних фаланг (у представителей рода Equus — одного среднего пальца).
Огромное, тяжелое тело парацератериев требовало относительного расширения площади опоры. Поэтому они не утеряли боковые ІІ-й и IV-й пальцы. Их метаподии, сдвинувшись несколько назад относительно средней и расширившись в сагиттальной плоскости, образовали вместе со средней метаподией, расширенной фронтально, нечто вроде прочного костного опорного полуцилиндра. Фаланги пальцев у них сократились в длину, а на среднем пальце еще и расширились; при этом они направились почти горизонтально, образовав увеличенную опорную площадь. Копытные фаланги всех пальцев расширились, их удлиненные, близко сходящиеся позади "ветви" охватили почти непрерывным прочным костным "лимбом" существовавшую, без сомнения, эластическую подушку. Все вместе представляло собою прочную, широкую и вместе с тем не слишком тяжелую подпорку для огромного тела животного.

Реконструкция внешнего облика "Indricotherium transouralicum". Автор — З. Буриан, 1980 г.
Своеобразное строение передних зубов гигантских носорогов можно объяснить следующим образом. Длинная и загнутая вниз передняя часть морды Paraceratherium transouralicum с огромными, также загнутыми вниз бивнеобразными верхними резцами на конце, вероятно, служила для пригибания и обламывания самых высоких веток. Значительное мышечное усилие при таком пригибании вызвало сильное развитие мускулов-сгибателей головы, что сказалось на черепе разрастанием глоточных бугров — мест их прикрепления. Функция нижних резцов должна была быть иной, чем верхних. Они не противостояли верхним резцов и не стирались о них. Направленные прямо вперед, с площадкой стирания, также обращенной вперед, массивные в передне-заднем поперечнике, глубоко укорененные в челюсти (с длинными корнями), они могли служить для сдирания древесной коры. Противодавление ствола или ветви, испытывавшееся резцами при этом сдирании, придавливало их друг к другу; в результате образовывались характерные площадки стирания на внутренних поверхностях нижних резцов-бивней. Помимо прочего, столь необычный аппарат — по два верхних и нижних несоприкасающихся коротких тупых бивня — мог использоваться как толстый и грубый пинцет для захвата масс веточного корма.

Реконструкция внешнего облика парацератерия (Paraceratherium transouralicum). Автор — Г. Угуэто, 2023 г.
Высказано предположение, что Paraceratherium transouralicum обитал в ландшафте с двумя четко выраженными сезонами — влажным и сухим. Во время сухого сезона листвы не было, и животное переходило на более жесткую диету — ело древесную кору.
Между разными родами и видами гигантских носорогов имеются в отношении рассмотренных приспособительных признаков, а также некоторых особенностей строения зубов, значительные отличия, также допускающие функциональное толкование. Paraceratherium bugtiense, таксон более поздний, чем Paraceratherium transouralicum, в некоторых отношениях ушел по пути специализации дальше, чем последний. Способность к движениям в суставах конечностей у него была еще более ограничена, что проявляется в форме поверхностей суставов локтевого, тазобедренного, коленного, частей сложного запястного сочленения, а отчасти и заплюсневого — все суставные поверхности у Paraceratherium bugtiense менее рельефны. О том же говорят более развитые у него тормозные приспособления на костях запястья, препятствующие верхнему переразгибанию. Заметно слабее у Paraceratherium bugtiense также суставный рельеф пальцевых фаланг.
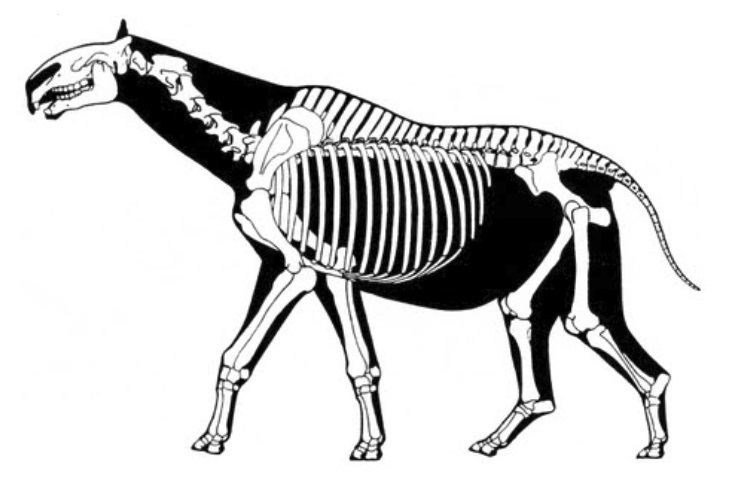
Реконструкция скелета парацератерия (Paraceratherium transouralicum) (по Г. Полу, 1997 г.).
Далее, у Paraceratherium bugtiense еще сильнее, чем у Paraceratherium transouralicum, выражена "монодактилизация": элементы, связанные с I-м и V-м пальцами — трапецийная кость, V-я метакарпальная, I-я клиновидная — более рудиментарны, а II-е и IV-е метаподии и фаланги сильнее сужены по сравнению с III-и, что сказывается и на относительно узких у Paraceratherium bugtiense костях запястья и заплюсны, связанных с этими пальцами. По-видимому, у этого вида еще более выпрямлены в суставах конечности, что проявляется в ослаблении плантарных и волярных связок на запястных и заплюсневых элементах (большая кость, крючковатая, кубовидная) и в меньшем рельефе мест прикрепления мышц — сгибателей и разгибателей. Еще более усовершенствовался и опорный аппарат Paraceratherium bugtiense: удлинились "ветви" последних фаланг, особенно боковых пальцев, на которых задние "ветви" сильно вытянулись назад; это сделало более прочными и компактными подставки для тяжелого тела животных.
Еще более пустотелыми стали шейные позвонки, воздушные камеры в которых увеличиваются даже в атланте, где у Paraceratherium transouralicum они были едва намечены.

Реконструкция внешнего облика парацератерия (Paraceratherium transouralicum) в сравнении с современным саванным слоном (Loxodonta africana). Автор — Ф. Шпиндлер.
Все описанные особенности у Paraceratherium bugtiense как бы продолжают их дальнейшее развитие, начатое у Paraceratherium transouralicum, делая его строение еще более присиособленным к огромному росту и питанию листьями высоких деревьев. Наряду с этими чертами специализации, у Paraceratherium bugtiense наблюдаются и такие, которые как бы возвращают его к несколько более архаической стадии развития. Такой характер имеет изменение у Paraceratherium bugtiense переднего отдела черепа. Огромные, загнутые вниз, верхние резцы-бивни Paraceratherium transouralicum, сидящие в длинных, также слегка загнутых вниз, предчелюстных костях, у Paraceratherium bugtiense сокращаются до незначительных рудиментов. Вместе с тем у него значительно укоротился весь предщечнозубый отдел черепа, что должно было сильно изменить облик головы животного. С укорочением морды связано, очевидно, и исчезновение у Paraceratherium bugtiense предкоренного Р1, имеющегося у Paraceratherium transouralicum. Напротив, нижние резцы-бивни у более позднего таксона стали еще массивнее; при этом на них чаще встречается и сильнее выражена площадка стирания, находящаяся на верхушке коронки, обращенная прямо вперед и совершенно плоская.
Описанное различие в строении передних зубов заставляет думать, что пищей Paraceratherium bugtiense служила не столько листва, сколько кора высоких деревьев: исчезла надобность пригибать (верхними бивнями) недоступные иначе, очень высокие, ветви; усиленное развитие нижних бивней показывает, что кора деревьев стала играть в его пищевом режиме большую роль, чем у Paraceratherium transouralicum.

Парацератерий (Paraceratherium transouralicum), обдирающий древесную кору. Автор — А. Атучин.
С последним предположением согласуются и некоторые различия между обоими таксонами в строении щечных зубов. У Paraceratherium bugtiense больше развита противошпора (антекроше) на верхних коренных и уплощен или даже слабо раздвоен внутри протокон, что несколько усиливает перетирающую способность зубов и указывает на изменение пищи в сторону большей твердости. То же значение имеет у Paraceratherium bugtiense окончательное завершение процесса слияния гребней коренных зубов из составляющих их конусов: исчезает моделировка пара- и метакона на верхних коренных и предкоренных и изолированность энтоконида на нижних предкоренных. Значение этого процесса, повторяющегося в эволюции многих групп копытных, заключается в приобретении большей компактности зубов и большей сопротивляемости их стиранию жестким растительным кормом (во многих случаях этот процесс сопровождается повышением коронки; у rигантских носорогов этого подметить не удается).

Реконструкция внешнего облика парацератерия (Paraceratherium transouralicum), основанная на воззрениях Д.Р. Протеро. Автор — К. Бьюэлл, 2013 г.
По аналогии с современными носорогами можно с высокой долей уверенности утверждать, что кожа парацератериев была толстой, серой, лишенной волос и, возможно, складчатой. Строение лицевого отдела черепа с костными возвышениями для прикрепления мышц допускает наличие у них подвижной верхней губы с хватательным придатком, приспособленной для ощипывания веток. По мнению некоторых исследователей, глубокая носовая вырезка даже подразумевает присутствие небольшого хоботка наподобие тапирьего. Тапиры используют свой хоботок для того, чтобы охватывать ветки и сдирать с них кору резцами; такая способность являлась полезной и для парацератериев.
Также высказано предположение о наличие у парацератериев больших ушей наподобие слоновьих, используемых для терморегуляции. Уши слонов увеличивают площадь поверхности тела и пронизаны сетью кровеносных сосудов, что облегчает отведение из организма избыточного тепла. Строение ушной области черепа парацератериев, особенно форма и расположение засуставного и заслухового отростков, соответствуют таковым у слонов не противоречит данной точке зрения. Впрочем, другие исследователи относятся к этой идее скептически и считают ее лишенной каких-либо оснований.

Современная пластическая реконструкция внешнего облика парацератерия (Paraceratherium transouralicum). Похожая на доспехи складчатая кожа по типу таковой современных носорогов из рода Rhinoceros является чисто умозрительным допущением.
Первоначально масса парацератериев была оценена 30-34 т. Эта оценка проистекала из сравнения с современными носорогами и отчасти основывалась на первой, неудачной реконструкции Г.Ф. Осборна. Однако вскоре выяснилось, что гигнтские носороги обладали гораздо более стройными пропорциями. Также было установлено, что скелет современных носорогов значительно мощнее, чем это необходимо для поддержания массы их тела, а многие древние представители группы были сложены куда более грацильно. Новые методы исследований снизили вес крупных видов парацератериев до 10-20 т, тогда как вес более мелких представителей рода был оценен в 7-9 т. Эти методы в значительной степени основаны на сравнении друг с другом отдельных элементов скелета, особенно черепа и костей конечностей, а также включают большое число других видов животных. Тем не менее, данные о весе гигантских носорогов следует рассматривать лишь как приблизительную оценку, поскольку по телесным пропорциям они значительно отличаются от других непарнопалых. Также высказано предположение, что 15-20 т являются максимально возможным весом для наземных млекопитающих из-за специфики обработки пищи в желудочно-кишечном тракте и гигантские носороги приблизились к этому пределу вплотную. Однако причина верхнего ограничения веса может быть экологической, а не биомеханической и, возможно, связана со стратегией воспроизводства.