в оглавление
назад
Другие роговые образования
Помимо волос, коже млекопитающих свойственны и другие роговые образования — мозоли, чешуи, когти, копыта, небные ороговения, шпоры, чехлы челюстей и рогов. Все они состоят из твердых, заполненных кератином клеток эпидермиса.
На безволосых участках тела, где кожа испытывает частое трение и давление, нередко формируются более или менее значительные утолщения ее рогового слоя — мозоли. Особенно мощные мозоли расположены на седалищных поверхностях некоторых обезьян, коленях и груди верблюдов (Camelus), запястных суставах бородавочников (Phacochoerus) и т. п. Кроме того, для многих млекопитающих характерны расположенные определенным образом на нижней поверхности лап подошвенные подушечки. Как правило, у пятипалых представителей класса в проксимальной части ладони или подошвы обычно имеется по паре подушечек, по одной у основания I-го пальца, ряд подушечек между основаниями остальных пальцев и по одной на кончике каждого пальца. У верблюдов распластанные по субстрату широкие пальцы покрыты сверху мягкой кожей, а снизу толстыми мозолистыми подушками. Маленькие копытца одевают концевые фаланги и едва участвуют в опоре. У коала (Phascolarctos cinereus) и приматов, которые при передвижении обхватывают кистями и стопами ветки деревьев, ладони и подошвы покрыты кожными гребешками, помогающими крепче удерживаться на ветках. Эти гребешки, расположенные на тех местах, где у представителей других групп находятся подушечки, образуют сложные папиллярные узоры из петель и завитков.
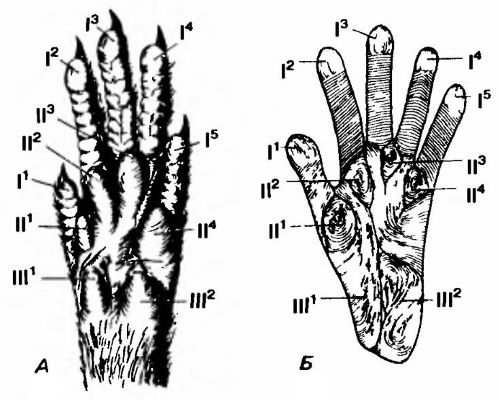
Ладонная поверхность кисти насекомоядного (А) и обезьяны (Б). Строение ладони насекомоядного можно, вероятно, рассматривать как примитивное для млекопитающих. Толстые подушечки (III1, III2) имеются на обеих сторонах проксимального конца ладони, подушечки между основаниями соседних пальцев (II1-II4) и на кончике каждого пальца (I1-I5) У высших приматов эти подушечки заменяются узором из кожных гребешков.
Для многих млекопитающих характерно наличие чешуевидных образований, приуроченных к определенным частям тела и играющих роль полезных приспособлений. Каждая чешуйка образована плоским кератиновым колпачком, надетым на короткий выступ дермы. Со временем поверхностный кератин стирается, но постепенно возобновляется из более глубоких слоев; общее число чешуй при этом остается постоянным. Как правило, чешуи у млекопитающих сочетаются с волосами. Чешуйчатый покров может иметься у них на лапах (различные землерои, например кроты (Talpinae) и большинство мышиных (Muridae)), на хвосте (мышевидные грызуны, бобры (Castoridae), выхухоли (Desmaninae) и многие другие насекомоядные, некоторые сумчатые) и на других участках тела, подвергающихся механическим раздражениям. Кольцевое расположение чешуй характерно для хвостов многих сумчатых, грызунов, насекомоядных. У шипохвостых (Anomaluridae) имеется двойной ряд чешуек на нижней поверхности хвоста у его основания. Наиболее сильно чешуйчатый покров развит у панголинов (Pholidota): их крупные, черепицеобразно налегающие друг на друга чешуи покрывают почти все тело за исключением морды, брюха и внутренней стороны конечностей. Сворачиваясь в клубок, панголины становятся неуязвимыми для большинства хищников. Рудименты чешуи присутствуют в области спинного плавника некоторых китообразных (морских свиней (Phocoena), гринд (Globicephala)). Имеются все основания полагать, что чешуи млекопитающих представляют собой не остатки чешуйчатого покрова их дальних предков, а являются вторичными образованиями, развившимися из собственных эмбриональных зачатков.

Чешуйчатый покров степного панголина (Smutsia temminckii).
Весьма совершенное защитное приспособление представляет собой панцирь броненосцев (Dasypodidae). Его образуют костные пластинки (производные дермы), покрываемые сверху роговыми щитками (производными эпидермиса). Панцирь покрывает спину и бока, образует «шапку» на голове и охватывает кольцами хвост. На конечностях он выражен меньше или совсем отсутствует, нет его и на брюхе. В зависимости от того, как, обороняясь от врага, животное пользуется панцирем, он может иметь разное устройство. Гигантский (Priodontes maximus) и щетинистый (Chaetophractus villosus) броненосцы при опасности стремительно закапываются брюшной стороной в почву. В связи с этим их плоский, состоящий из крупных пластин панцирь защищает только спину и бока. Мелкий плащеносный броненосец (Chlamyphorus truncatus) вырывает нору, прикрывая ее своим плоским бронированным тылом. Трехпоясный шаровой (Tolypeutes tricinctus) и девятипоясный (Dasypus novemcinctus) броненосцы сворачиваются в плотный, непроницаемый шар. Тело этих видов покрыто выпуклым, гибким панцирем, состоящим из узких кольцеобразных полос и дающим возможность скрыть все тело.
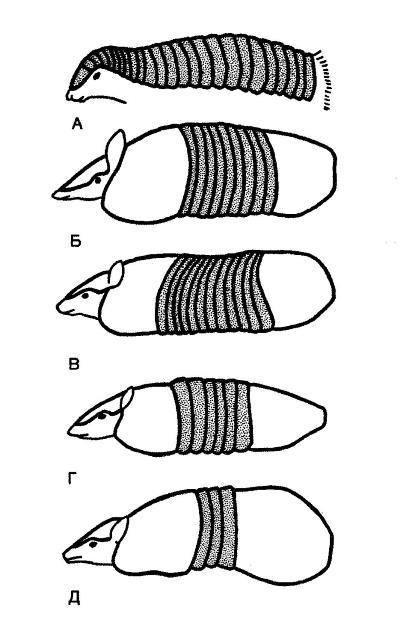
Схема панциря и объем его подвижной части (заштриховано) у разных броненосцев: А — плащеносный броненосец (Chlamyphorus truncatus); Б — девятипоясный броненосец (Dasypus novemcinctus); В — большой голохвостый броненосец (Cabassous unicinctus); Г — белощетинистый броненосец (Euphractus sexcinctus); Д — трехпоясный броненосец (Tolypeutes tricinctus).
Когти, ногти и копыта — это кератинизированные эпидермальные структуры на кончиках пальцев. Исходным типом является коготь, а ноготь и копыто — его модификации, возникшие у млекопитающих. Наружная часть когтя представлена выпуклой и твердой когтевой пластинкой, прикрывающей когтевой шип концевой фаланги сверху и с боков подобно чехлику. В поперечном разрезе когтевая пластинка имеет форму перевернутой буквы V, все сильнее сужающейся к концу, а дистальнее кончика пальца загнутой вниз. У своего основания она защищена складкой кожи; из этого зачаткового ложа кератинизированный эпителий непрерывно растет наружу, поверх дермы. Снизу когтевого шипа расположена более рыхлая подошвенная пластинка (subunguis), нарастающая из глубины подобно типичному эпидермису и граничащая с мягкой подушечкой концевой фаланги. В процессе роста когтевая пластинка непрерывно сползает к вершине пальца, и там выступает за край подошвенной пластинки. Более медленное изнашивание когтевой пластинки по сравнению с подошвенной обусловливает сохранение остроты когтя.
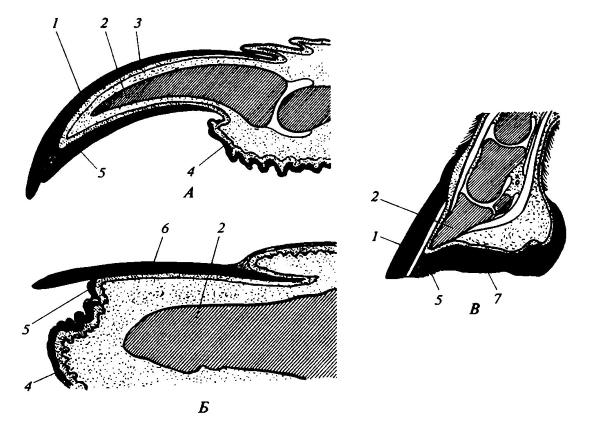
Продольные вертикальные разрезы через типичный коготь (А), ноготь человека (Б) и копыто лошади (В): 1 — когтевая пластинка; 2 — костный шип концевой фаланги пальца; 3 — дерма; 4 — подушечка пальца; 5 — подошвенная пластинка; 6 — ногтевая пластинка; 7 — стрелка копыта.
Когти и их видоизменения защищают конечные фаланги пальцев от механических повреждений, служат орудиями нападения и защиты, лазания и рытья. У роющих форм когти удлинены и уплощены, древесных — остры и сильно изогнуты. Особой остроты достигают втяжные когти кошек (Felidae) (кроме гепарда (Acinonyx jubatus)): коготь вместе с концевой фалангой особыми сухожилиями притягивается к дорсальной поверхности предпоследней фаланги. Вследствие наличия такого аппарата когти кошек не только не тупятся при ходьбе, но и облегчают захват и удержание подвижной добычи. Втяжные когти также присутствуют у некоторых виверр (Viverridae), мадагаскарских хищников (Eupleridae), азиатских линзангов (Prionodontidae) и вымерших нимравов (Nimravidae).
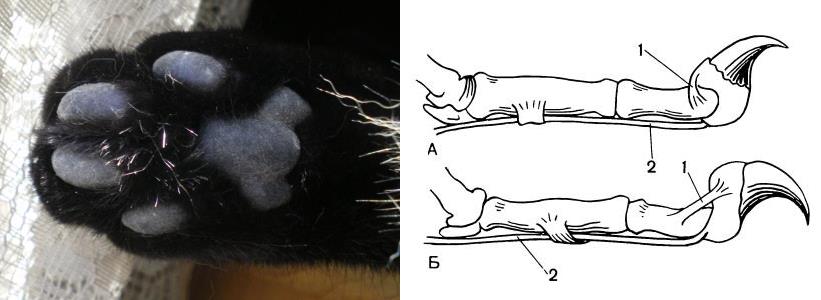
Слева — лапа домашней кошки (Felis silvestris catus), справа — скелет пальца с втянутым (А) и выпущенным (Б) когтем (1 — эластичная связка; 2 — сухожилие).
У некоторых млекопитающих отдельные когти приобретают специфическое назначение и изменяют соответственно свой внешний вид. Таковы, например, раздвоенные клещеобразные когти бобров (Castor), служащие в первую очередь для выжимания воды из ости, а также для смазывания меха жирным выделением анальных желез и вычесывания из него эктопаразитов. У кенгуру (Macropodiformes) подобные когти несут только последнюю функцию.
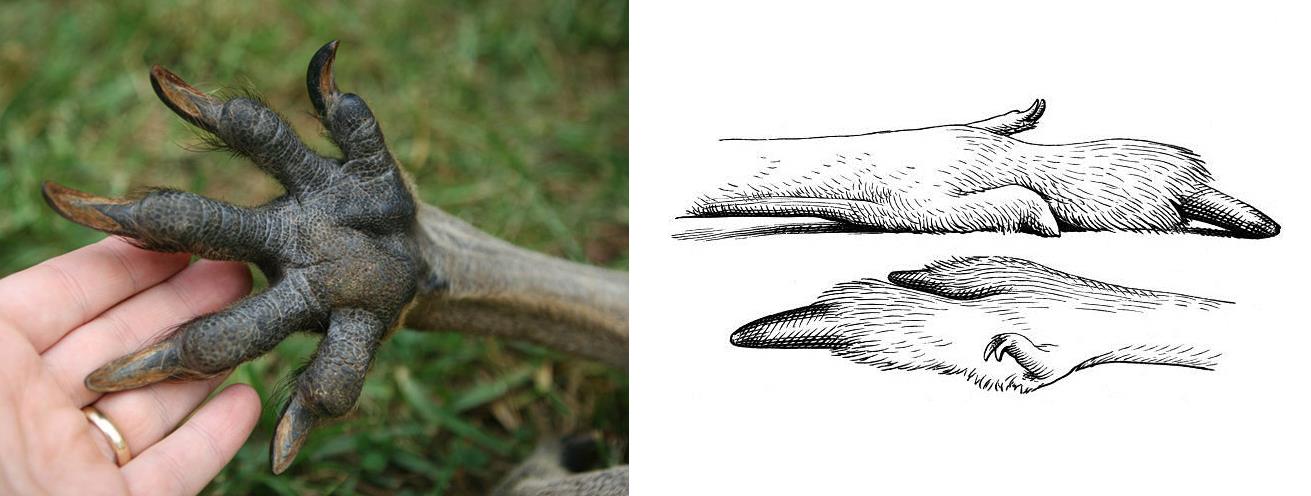
Лапы восточного серого кенгуру (Масropus giganteus): слева — передняя, справа — задняя с чесальными двойными когтями на недоразвитых сращенных II-м и III-м пальцах.
У водных и летающих форм когти претерпевают редукцию. Они недоразвиты на передних конечностях сирен и ластоногих. У китообразных когти либо полностью отсутствуют, либо зачаточны; у некоторых видов они имеются в виде провизорных зачатков только у зародышей. Передние конечности рукокрылых утратили когти на большинстве пальцев.
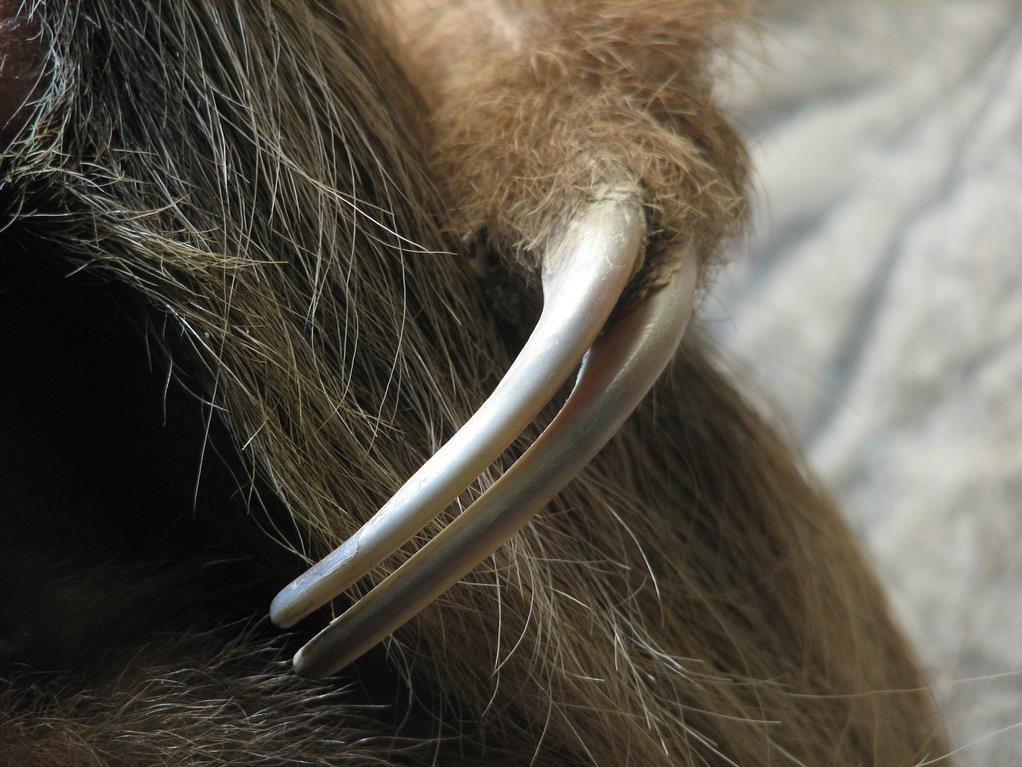

Слева — лапа ленивца Гоффмана (Choloepus hoffmanni), справа — передний ласт американского ламантина (Trichechus manatus).
У многих приматов когти преобразовались в ногти. Ноготь отличается расширением и уплощением когтевой пластинки, прикрывающей концевую фалангу только сверху, наличием в его начале кожистой оторочки — когтевого валика, значительной редукцией подошвенной пластинки и сильным развитием выдающейся за ее пределы мягкой подушечки, повышающей осязательные способности и цепкость пальцев.

Слева — кисть руконожки (Daubentonia madagascariensis), справа — кисть обыкновенного шимпанзе (Pan troglodytes).
Наоборот, у наземных травоядных, у которых число пальцев уменьшено и тело при передвижении опирается на кончики оставшихся пальцев, усложнение когтей привело к формированию толстых роговых копыт. Их мощные когтевая и подошвенная пластинки, а у лошадей и ороговевшая подушечка пальца (стрелка) в совокупности образуют футляр, почти целиком охватывающий концевую фалангу. При этом передняя стенка копыта соответствует когтевой пластинке, его подошва — подошвенной пластинке, тогда как относительно твердая стрелка вдается в копыто угловатым выступом сзади. Когтевой валик в копыте совершенно не выражен. Непосредственно с грунтом контактирует дугообразный или V-образный дистальный конец копыта. Особенно хорошо копыта развиты у быстро бегающих копытных — лошадей, антилоп, козлов. При этом у лесных видов (например, у оленей), часто ходящих по болотам, копыта более широкие и плоские. У степных (антилопы) и особенно у горных видов (козлы, бараны) копыта маленькие, узкие; площадь опоры у них значительно меньшая, чем у лесных копытных, ходящих зачастую по размягченному грунту или снегу. Так, весовая нагрузка на 1 кв. см подошвы у центральноазиатского горного козла (Саpra sibirica) равна в среднем 850 г, у лося (Alces alces) — 500 г, у северного оленя (Rangifer tarandus) — всего 140 г.
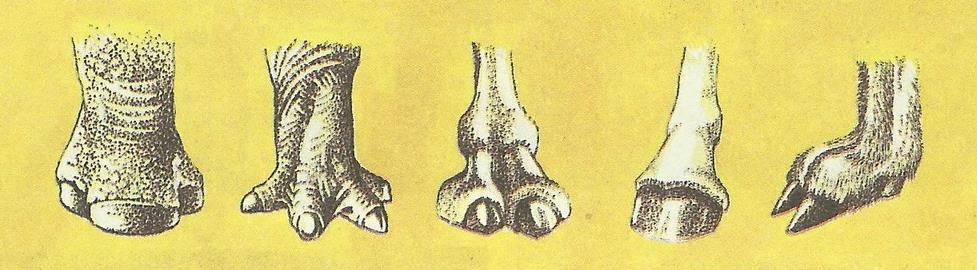
Различные типы копыт, слева направо — носорог, карликовый бегемот, верблюд, лошадь, олень.
У значительной части млекопитающих когти подвержены сезонным изменениям. Так, у копытного лемминга (Dicrostonyx torquatus) летом когти тонки, зимой же вследствие разрастания подошвенной пластинки и ороговения подушечки пальца они превращаются в род копытец (приспособление к разрыванию твердого снега); весной наросшая часть отпадает. Когти монгольской пищухи (Ochotona pallasi), песца (Vulpes lagopus) и др. зимой длиннее и острее, чем летом. Значительным сезонным изменениям подвержены также копыта, особенно у северного оленя (летом они менее развиты, чем зимой, когда служат для раскапывания снега).
В пяточной области задней конечности однопроходных имеется роговая шпора с костной основой, особенно сильно развитая у самцов. Шпора пронизана каналом, связанным с т. н. голенной железой. У самцов утконоса (Ornithorhynchus anatinus) во время репродуктивного сезона эта железа увеличивается и начинает вырабатывать яд. Его сила такова, что может убить не только мелкое, но и средних размеров животное, например, динго. Свои шпоры утконосы используют в основном в турнирных сражениях, но они могут выполнять и защитную функцию — например, введенный человеку яд вызывает сильную боль и вздутие тканей. У самцов ехидн (Tachyglossidae) голенные железы в период спаривания также увеличиваются, но поскольку ехидны не способны наносить удары своими шпорами, случаи интоксикации секретом их желез неизвестны. Все же удалось установить, что самцы ехидн используют вырабатываемый железами секрет для запахового информирования о своем репродуктивном статусе самок или самцов-конкурентов. Представляется вероятным, что голенная железа всех однопроходных некогда вырабатывала яд. У утконоса ядовитый аппарат сохранился, а у ехидн преобразовался в средство запаховой коммуникации. Что же касается самок однопроходных, то у них шпоры никак не используются и нередко утрачиваются еще в молодом возрасте.

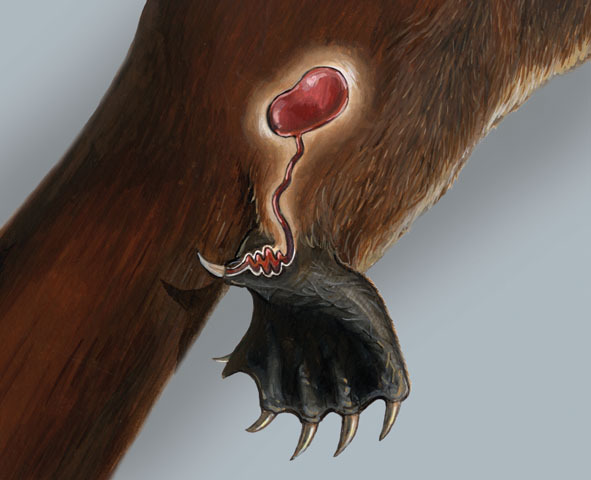
Роговая шпора (слева) и схема ядовитого аппарата (справа) самца утконоса (Ornithorhynchus anatinus).
Помимо однопроходных шпоры обнаружены у представителей других архаичных групп млекопитающих и млекопитающеобразных — кюнеотениев (Kuehneotheria), эвтриконодонтов (Eutriconodonta), спалакотероидов (Spalacotheroidea), многобугорчатых (Multituberculata), докодонтов (Docodonta). Судя по всему, шпорами обладал и общий предок класса млекопитающих, однако сумчатыми и плацентарными они были утрачены.
Кроме того, роговыми образованиями эпидермиса являются хвостовые шпоры львов (Panthera leo) и леопардов (Panthera pardus), а также колючки и шипики, свойственные половому члену многих грызунов (например, тушканчиков (Dipodidae)), а также кошек (Felidae) (тот же лев (Panthera leo), тигр (Panthera tigris)). У пустынных землероев (сумчатых кротов (Notoryctidae), златокротов (Chrysochloridae)) на кончике морды развивается роговой колпачок для раздвигания сухого, каменистого грунта. Роговые чехлы, покрытые сверху кожей, защищают от повреждений челюсти однопроходных. Небные роговые образования у млекопитающих представлены в виде утолщений небных валиков (хищные, копытные, сирены) или бахромчатых пластин (китовый ус беззубых китов).
На голове у многих копытных имеются различного рода рогообразные выросты. Часто степень их развитости или само их наличие является половым признаком. Как правило, рога сильнее развиты у самцов, которые используют их для внутривидовых демонстраций и в качестве турнирного оружия. Также рога служат защитой от хищников. Для копытных, лишенных острых зубов и когтей, это очень существенно. Появившись в виде небольших наростов, рога в процессе эволюции постепенно становились все более крупными; аналогичное изменение имеет место и в онтогенезе. Значительное разнообразие рогов по положению, величине и форме делает их важным диагностическим признаком.
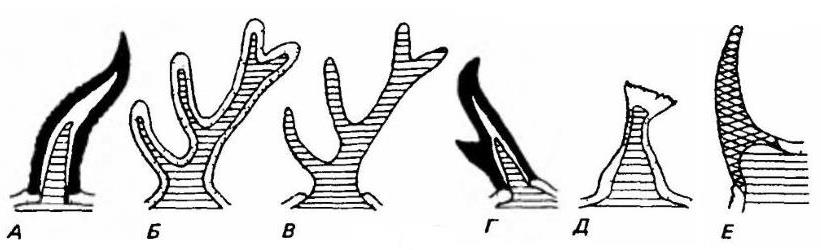
Схемы рогов разных типов в продольном разрезе. Рога обращены передней стороной влево. А — рог полорогого с роговым чехлом на костной основе. Б — олений рог, покрытый кожей; когда рога завершают свой рост, эта кожа отмирает и отпадает. В — тот же рог без кожи. Г — рог вилорога. Д — рог жирафа. Е — рог носорога. На всех рисунках костный отросток черепа заштрихован, кожа покрыта точками, роговое вещество зачернено, а непарный рог носорога покрыт сеткой.
Чисто эпидермальные образования представляют собой массивные рога носорогов (Rhinocerotidae). Они состоят из склеенных в сплошную роговую массу кератинизированных волосовидных сосочков, образующих прочный конус, возвышающийся над шероховатыми носовыми и лобными костями. Костного стержня такие рога лишены. Длина переднего рога белого носорога (Ceratotherium simum) может достигать 160 см. Поскольку рога обладают неограниченным ростом, носороги часто трутся ими о жесткие объекты с целью сохранения нужной формы и величины.
У бронтотериев (Brontotheriidae) костные poгoобразные выросты помещались на носовых костях и могли достигать большой величины. У многих диноцерат (Dinocerata) и у арсинойтериев (Arsinoitherium) из эмбритопод (Embrithopoda) парные костные рога имелись как на носовых, так и на лобных костях. Рога указанных животных были обтянуты кожей либо дополнительно укреплялись роговыми чехлами.
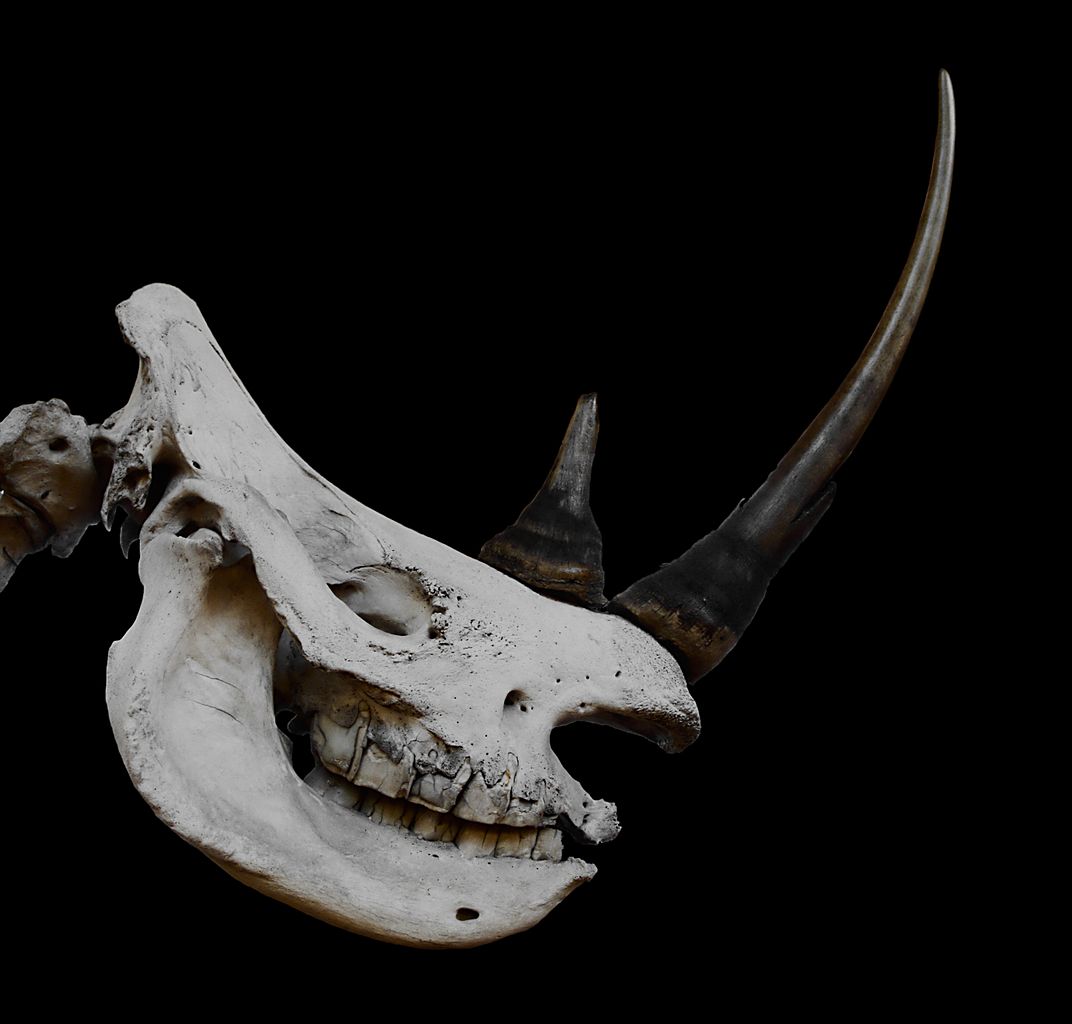
Череп и рога белого носорога (Ceratotherium simum).
У жвачных (Ruminantia) основу рогов формируют кожные кости (cornua), образованные дермой и в большинстве случаев тесно срастающиеся с надкостницей лобных костей. У оленей (Cervidae) выступ лобной кости дает начало постоянному основанию рога — т. н. пеньку, который всегда скрыт под кожей. От пенька растет образованный кожной костью неветвящийся стержень, именуемый штангой. С возрастом на штанге появляются боковые отростки, со временем их становится все больше. Рога благородного оленя (Cervus elaphus) достигают максимального развития на 9-й год; даже менее крупным видам, таким, как лань (Dama dama) и косуля (Capreolus capreolus), требуется 5-6 лет, чтобы их рога полностью созрели. В течение нескольких последующих лет их размер и форма остаются довольно постоянными; разумеется, при условии, что олени получают достаточное питание. По мере того, как животное стареют, их рога теряют отростки и уменьшаются в размерах. У некоторых оленей рога не ветвятся, оставаясь на стадии штанги в течение всей жизни. Обычно они свойственны только самцам, но у северного оленя (Rangifer tarandus) имеются и у самок, для которых являются важным средством адаптации к экстремальным условиям жизни. У архаичного водяного оленя (Hydropotes inermis) рога совсем отсутствуют.
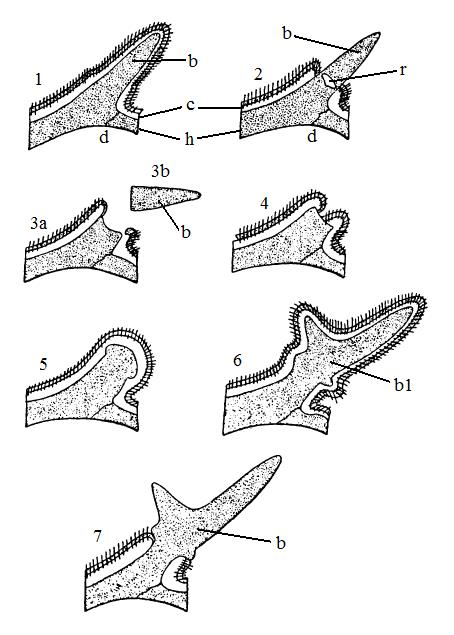
Развитие рога благородного оленя (Cervus elaphus): 1 — начало появления рога в виде выроста (a) лобной кости, он покрыт кожей и волосами; 2 — стержень рога (b) обнажился, в его основании в результате разрушения костного вещества образовалась полость (r); 3 — пенек рога (За) со сброшенным стержнем (Зb); 4 — процесс зарастания пенька кожей; 5 — образование венчика в пеньке, обросшем кожей; 6 — развитие вторичного рога (b1) под одетой волосами кожей; 7 — очистившийся от кожи рог с 2 отростками — вилка; е — эпидермис с волосами; с — дерма; h — кость; d — венечный шов.
Молодые рога оленей называются пантами. Они сложены из хряща, а сверху покрыты кожей, пронизанной многочисленными кровеносными сосудами и покрытой короткими волосами. По мере роста рога окостеневают в направлении снизу вверх. Их губчатое костное вещество также пронизывают сосуды — ветви временной артерии (arteria tempоralis). После окостенения рогов их питание кровью прекращается, а поверхностная кожа отмирает и слущивается. Для ускорения этого процесса олени трутся рогами о стволы деревьев, камни и землю. По окончании роста и отпадания кожи рога представляют собой чисто костные образования, на поверхности которых сохраняются углубления и шероховатости — следы сосудов. Такие рога регулярно сбрасываются. У ныне живущих оленей это происходит ежегодно зимой или ранней весной, и лишь у оленя Давида (Elaphurus davidianus) рога сбрасываются дважды в году. В пределах прилежащих к пенькам узких поясков костная ткань резорбируется под действием гидролитических ферментов, выделяемых специальными клетками остеокластами, и рога отпадают. Пеньки быстро зарастают особыми хрящевыми шапками, покрытыми кожей. Вскоре из них начинают развиваться молодые рога. Отрастание рогов продолжается примерно 5 месяцев и завершается к началу гона. Весь процесс проходит под влиянием эффективных биостимуляторов и ложится тяжелым бременем на обмен веществ. Животные в этот период вынуждены интенсивно потреблять известь и фосфаты. Жизнестойкость организма оленей поражает, ведь вес регулярно сбрасываемых рогов у благородного оленя (Cervus elaphus) достигает 13 кг, у лося (Alces alces) — 20 кг при размахе 2 м, а у недавно вымершего большерогого оленя (Megaloceros giganteus) — даже 40 кг при размахе в 3,7 м.

Рост рогов у белохвостого оленя (Odocoileus virginianus).
Несколько иной характер имеют постоянные рога жирафов (Giraffidae). Они почти всегда имеют небольшие размеры и не ветвятся, в течение жизни полностью или частично покрыты кожей с волосами. Исключение составляли сиватерии (Sivatheriinae) с крупными ветвистыми рогами, которые могли покрывать роговые чехлы. Обычно одна или две пары рогов располагается на лобных или частично на теменных костях жирафов; иногда, кроме того, имеется еще непарный рог на лобных или носовых костях. Степень пневматизации костных стержней рогов различна.

Череп жирафа (Giraffa camelopardalis). С возрастом у самца формируется костное утолщение на лбу. Оно оказывается жизненно необходимым, поскольку защищает мозг, который иначе мог бы быть поврежден в яростных соударениях самцов. Кость нарастает со скоростью 1 кг в год.
Полорогие (Bovidae) имеют неветвящиеся рога в виде полых роговых чехлов, образованных кератинизованным эпидермисом и сидящих на костных стержнях. Такой стержень формируется из небольшого выроста лобной кости и очень рано срастающейся с ним кожной кости. В большей или меньшей степени плотное костное вещество имеется только с поверхности лобного отростка, в его глубине оно мелкогубчатое или даже заменено крупными полостями (синусами), разделенными прочными костными перегородками. Полости эти в разной степени глубоко проникают в вершину отростка и большей частью связаны с воздухоносными полостями лобной кости. Вырост лобной кости соответствует пеньку в оленьем роге, а нарастающую над ним кожную кость можно сравнить со штангой.

Череп муфлона (Ovis orientalis) с роговыми чехлами, спирально изогнутыми и покрытыми складками роста.
Костная внутренняя часть рога с годами уплотняется и утолщается. В соответствии с этим утолщается и роговой чехол рога, в медленно растущем основании которого каждый год налагается новая складка (зимой рост рогов замедляется). Таким образом, в противоположность рогам оленей, рога полорогих растут в своей нижней части. Со временем темп их роста все более замедляется, а покрывающие чехол складки сморщиваются и сливаются. Рога сильно различаются по величине и форме: у мелких антилоп тонкие и изящные, у крупных антилоп прямые или саблевидно изогнутые, у козлов и баранов толстые, часто спиралевидные или винтообразные, у овцебыка (Ovibos moschatus) и африканского буйвола (Syncerus caffer) очень массивные, образующие на лбу крепкий панцирь. Все они отличаются чрезвычайной прочностью и выдерживать сокрушительные удары. Рога полорогих постоянны и никогда не сбрасываются: стираются лишь поверхностные слои рогового вещества, которые постепенно возобновляются изнутри.

Рост рогов у толсторогого барана (Ovis canadensis).
У вилорога (Antilocapra americana), единственного современного представителя семейства Antilocapridae, роговые чехлы ежегодно сбрасываются и формируются заново. Чехлы эти имеют небольшую боковую ветвь. Плотные неветвистые костные стержни — основа чехлов — не сбрасываются.
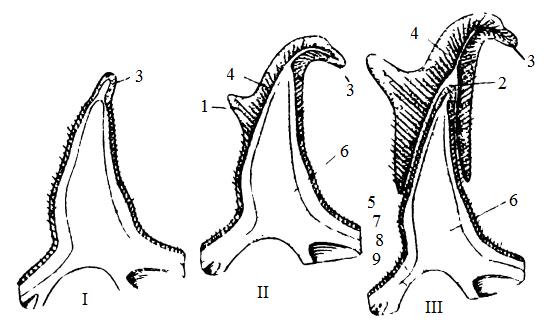
Развитие рога у вилорога (Antilocapra americana): I — вскоре после сброса роговой оболочки (чехла); II — более поздняя стадия с образованием переднего рогового отростка (1); III — под старым роговым чехлом, отделившимся от его основы, возникает новый волосяной покров и новый роговой чехлик (2); 3 — первый роговой отросток; 4 — второй роговой отросток; 5 — основание рогового чехла; 6 — костный лобный вырост; 7 — эпидермис; 8 — кожный покров; 9 — лобная кость.