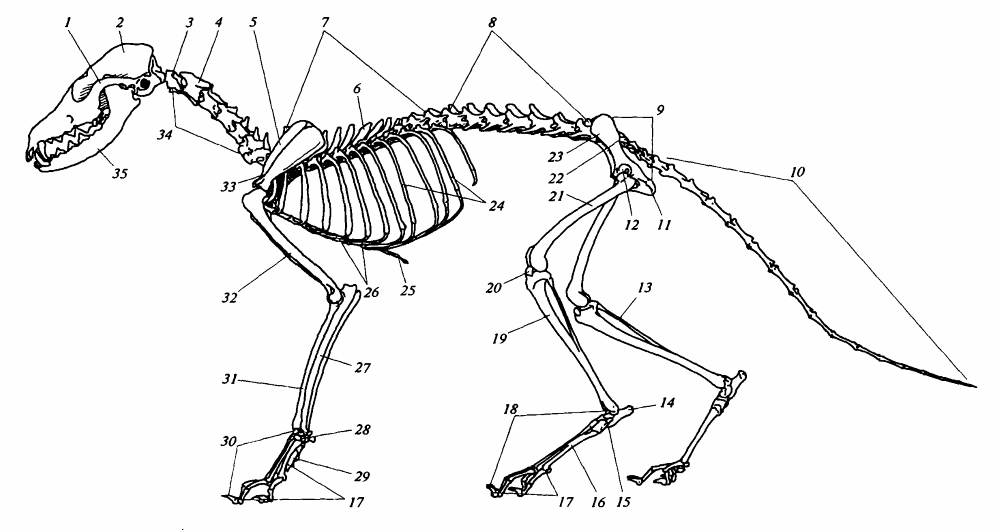
Скелет лисицы (Vulpes vulpes): 1 — скуловая дуга; 2 — череп (cranium); 3 — атлант (atlas); 4 — эпистрофей (epistropheus); 5 — лопатка (scapula); 6 — остистый отросток; 7 — грудные позвонки (vertebrae thoracales); 8 — поясничные позвонки (vertebrae lumbales); 9 — таз (pelvis); 10 — хвостовые позвонки (vertebrae caudales); 11 — седалищная кость (ischium); 12 — вертлужная впадина; 13 — малая берцовая кость (fibula) (приросшая); 14 — пяточная кость; 15 — предплюсна (tarsus); 16 — плюсна (metatarsus); 17 — фаланги пальцев(phalangae digitorum); 18 — стопа; 19 — большая берцовая кость (tibia); 20 — коленная чашечка (patella); 21 — бедренная кость (femur); 22 — крестцовые позвонки (vertebrae sacrales); 23 — подвздошная кость (ileum); 24 — позвоночные отделы ребер; 25 — грудина (sternum); 26 — грудинные отделы ребер (хрящевые); 27 — локтевая кость (ulna); 28 — запястье (carpus); 29 — пясть (metacarpus); 30 — кисть; 31 — лучевая кость (radius); 32 — плечевая кость (humerus); 33 — лопаточная ость (spina scapulae); 34 — шейные позвонки (vertebrae cervicales); 35 — нижняя челюсть (mandibula).
Позвоночный столб
Позвоночный столб (columna vertebralis), или позвоночник, представляет собой образованный позвонками (vertebrae) членистый стержень. У млекопитающих он состоит из четко выраженных шейного, грудного, поясничного, крестцового и хвостового отделов. По своему общему контуру позвоночный столб похож на таковой терапсид. У среднестатистического млекопитающего шейные позвонки идут от головы косо вниз. Выше плечевой области, приблизительно там, где начинаются спинные позвонки, кривая позвоночника подымается, а затем опускается через поясничную область к крестцу и хвосту.
Позвонки состоят из тел (corpus vertebrae), дуг и отростков. За исключением первых 2 шейных, свободные сочленения между позвонками отсутствуют. Вместо этого их тела соединены эластичными прокладками — межпозвоночными дисками, или менисками из волокнистого хряща, которые сминаются при взаимных поворотах позвонков. Внутри менисков, в виде студенистых ядер, сохраняются остатки хорды. Тела позвонков обладают плоскими торцевыми поверхностями и являются по этому признаку платицельными; только у копытных их строение приближаются к опистоцельному (выпукло-вогнутому) типу. Сверху к телу позвонка примыкает хорошо развитая верхняя, или невральная, дуга (arcus neuralis). В совокупности верхние дуги образуют позвоночный канал — защитный футляр для спинного мозга. Кроме того, у позвонка имеются остистый и поперечные отростки. Остистый отросток отходит от тела позвонка вверх. Поперечные отростки отходят от боковых оснований верхних дуг и только в поясничной области иногда сидят прямо на телах позвонков. Кроме того, как и у всех тетрапод, у основания дуги с каждой стороны располагаются сочленовные отростки, которые соединены скользящими суставами с сочленовными отростками смежных позвонков. У китообразных взаимная связь позвонков посредством суставных отростков ослаблена. В ней нет необходимости, как это требуется для сухопутных животных — у водных позвоночных потеря в весе, равная весу вытесненной телом воды, сильно уменьшает нагрузку на позвоночник и суставные отростки позвонков. У ряда китообразных в хвостовом отделе позвоночника дополнительно присутствуют также нижние, или гемальные, дуги.
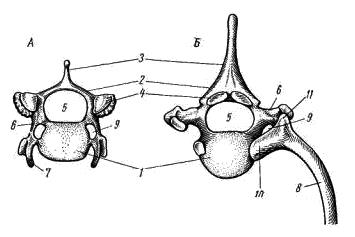
Позвонки лисицы (Vulpes vulpes), вид спереди: А — шейный позвонок; Б — грудной позвонок; 1 — тело позвонка, 2 — верхняя дуга, 3 — остистый отросток, 4 — сочленовные поверхности верхних дуг (для сочленения соседних позвонков), 5 — канал для спинного мозга, 6 — поперечный отросток, 7 — рудимент шейного ребра, 8 — ребро, 9 — отверстие для кровеносных сосудов, 10 — головка ребра, 11 — бугорок ребра.
Для млекопитающих характерно постоянное число шейных позвонков — 7, от длины тел которых и зависит длина шеи. Сильно укороченная и негибкая шея китов слагается из буквально листовидных позвонков, которые к тому же нередко срастаются между собой. Укороченный шейный отдел и небольшая подвижность головы также характерна для норников, особенно землероев. Хорошо развита шея у хищников. Наиболее сильно шейный отдел развит у копытных, для которых подвижность головы очень важна при добыче пищи. Но даже чрезвычайно длинная шея жирафа (Giraffa camelopardalis) поддерживается 7 очень вытянутыми позвонками. Исключение из общего правила составляют двупалые ленивцы (Choloepus) с 5 или 6 шейными позвонками, ламантины (Trichechus) — с 6, тамандуа (Tamandua) и панголины (Manidae) — с 8, а также трехпалые ленивцы (Bradypus), у которых их бывает от 8 до 10.
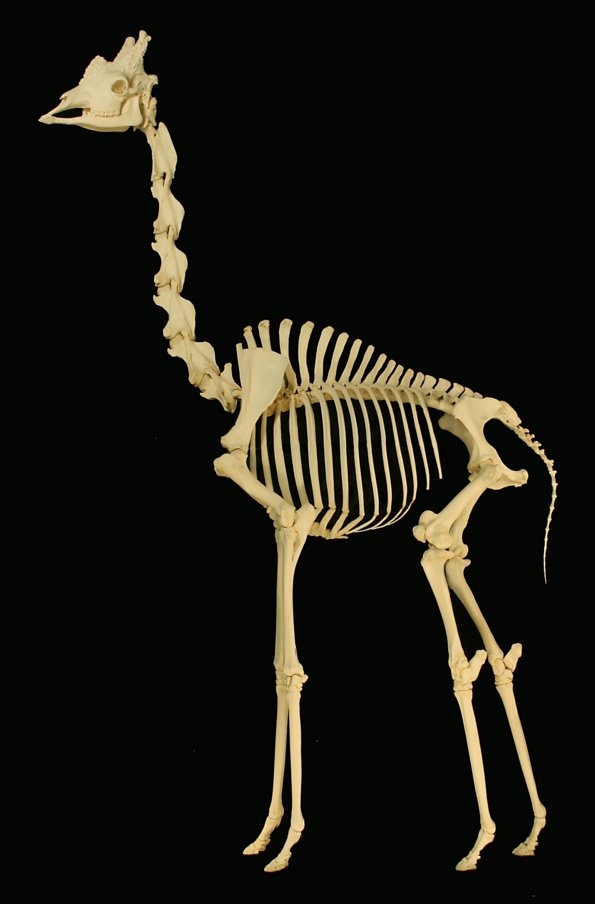
Смонтированный скелет жирафа (Giraffa camelopardalis). Длина шеи взрослого животного достигает 2,4 м.
Как и у других амниот, 2 передних шейных позвонка сильно специализированы. Атлант (atlas) представляет собой кольцо, состоящее из невральной дуги и интерцентра. Спереди атлант несет 2 сочленовные поверхности для соединения с парными мыщелками затылочной кости. За ним следует более крупный эпистрофей (epistropheus, или axis), имеющий на своем переднем конце гладкий конический зубовидный отросток (processus odontoideus) — приросшее тело атланта. Этот отросток входит в ямку атланта, который на нем и поворачивается. Вращательная подвижность здесь особенно важна, поскольку причленение черепных мыщелков к атланту такой степени свободы не дает. Таким образом, относительно атланта голова может двигаться вверх и вниз; боковые движения головы и ее ротация вдоль продольной оси происходят преимущественно за счет движения атланта относительно эпистрофея. Благодаря специфике строения сочленовных поверхностей шейных позвонков голова ленивцев может поворачиваться на 270 °.

Смонтированный смонтированный скелет древнего бизона (Bison antiquus).
В грудном отделе позвоночника насчитывается 12-15 позвонков; в большинстве случаев их 13. Исключение составляют бутылконосы (Hyperoodon) и один род броненосцев с 9 грудными позвонками, а также двупалые ленивцы (Choloepus), у которых этих позвонков 24. Число поясничных позвонков варьирует от 2 до 9, чаще всего их 5-7. В этих двух отделах, особенно в грудном, хорошо развиты остистые отростки позвонков, которые у большинства грудных позвонков наклонены назад, а на поясничных позвонках — вперед. Но все они оказываются вертикальными и максимально эффективны в качестве рычагов в начале толчка обеими задними конечностями, когда позвоночник выгнут вверх и энергично распрямляется усилием специальной мышцы. У ксенартр на последних грудных и на поясничных позвонках имеются своеобразные придаточные сочленения.

Смонтированный скелет длиннохвостого панголина (Phataginus tetradactyla).
Крестцовых позвонков, в значительной степени срастающихся между собой, может быть от 1 до 10, но наиболее часто их 4 (у хищных — 3). Однако только 2 передних позвонка, сросшиеся с подвздошными костями, считаются истинно крестцовыми, остальные же представляют собой приросшие к ним хвостовые позвонки. У утконоса (Ornithorhynchus anatinus), как и у рептилий, только 2 крестцовых позвонка. Крестец (sacrum) предоставляет жесткую опору для тазового пояса.
Хвостовой отдел позвоночника формирует хвост. Во многих случаях он является важным органом, выполняющим ряд разнообразных функций. Каждой из этих функций соответствует своя длина, толщина и общий вид хвоста. В других случаях хвост совсем не используется и подлежит сильной редукции. В этой связи число хвостовых позвонков подвержено большой изменчивости: так, у землеройкового тенрека (Microgale principula) их 47, длиннохвостого панголина (Phataginus tetradactyla) — до 50, а у гиббонов (Hylobatidae) и некоторых летучих мышей — всего 3.
Смонтированный скелет паукообразной обезьяны (Ateles sp.).
Степень подвижности отдельных позвонков различна. У мелких бегающих и лазающих зверьков она велика по всей длине позвоночника, поэтому их тело может изгибаться в разных направлениях и даже свертываться в клубок. Остистые отростки у этих видов низкие и тонкие. У большинства других млекопитающих наибольшей подвижностью обладают шейный и хвостовой отделы позвоночника, менее подвижны поясничный и грудной отделы и совершенно неподвижен крестцовый. Гибкость шеи связана с многообразием ее функций, прежде всего с участием в захватывании пищи. У копытных наиболее крупные позвонки с развитыми остистыми отростками располагаются в передней части грудного отдела, где к ним прикрепляется мощная мускулатура шеи и отчасти передних конечностей. У млекопитающих, передвигающихся на задних ногах (кенгуру (Macropodidae), тушканчики (Dipodidae), прыгунчики (Macroscelididae)), наоборот, позвонки крупнее в крестце и основании хвоста, а далее вперед их величина последовательно убывает.
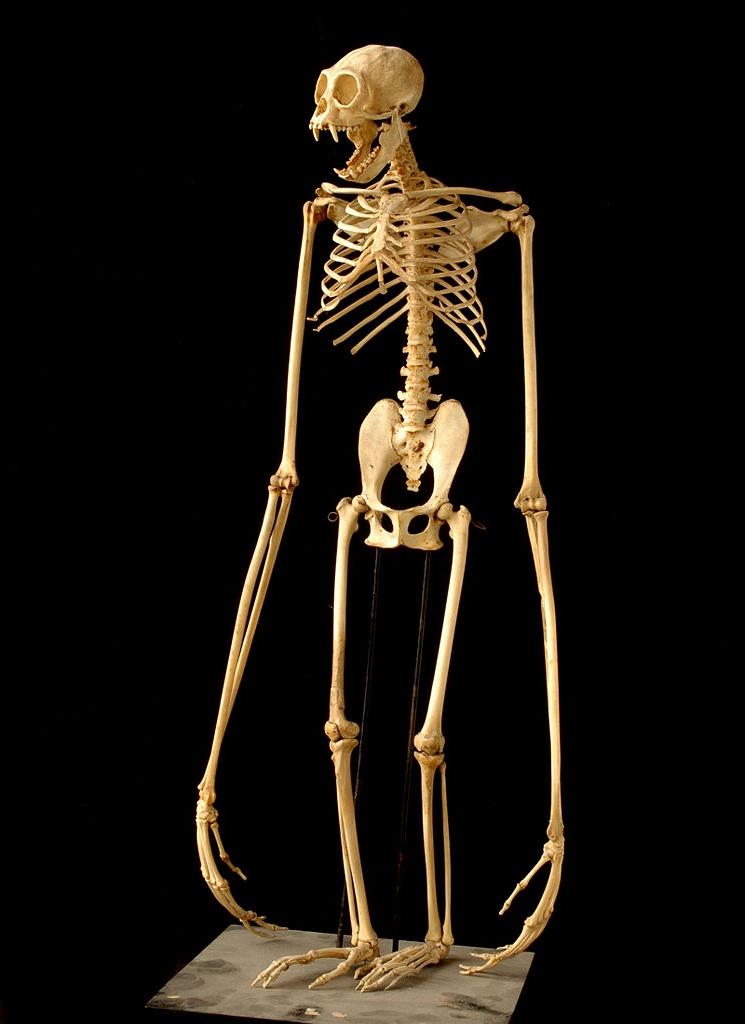
Смонтированный скелет гиббона (Hylobates sp.).
У белозубок-броненосок (Scutisorex) поперечные отростки позвонков сильно разрастаются и посредством многочисленных выростов сочленяются с передними и задними позвонками. Эти дополнительные сочленения настолько укрепляют позвоночный столб, что эти зверьки способны выдержать без вреда для себя вес взрослого человека. Данное приспособление позволяет броненоскам охотиться на насекомых в каменистых осыпях без риска быть раздавленными.

Поясничный отдел позвоночника угандской белозубки-броненоски (Scutisorex somereni).
У водных млекопитающих, использующих в основном движущую силу хвоста, его позвонки отличаются большими размерами и обладают длинными толстыми отростками для крепления сильных мышц. Таково строение хвостового отдела ондатры (Ondatra zibethicus), бобров (Castoridae), выдр (Lutrinae), китообразных. Хвост прыгающих и древесных форм, выполняющий прежде всего функцию балансира, тоже длинный, но позвонки тонкие, и отростки у них также тонкие и длинные. Хвостовая мускулатура этих животных по сравнению с предыдущей группой слабая, так как воздух оказывает меньшее сопротивление, чем вода. Подобным хвостом обладают тушканчики (Dipodidae), белки (Sciuridae), куницы (Mustelinae) и большинство приматов, ведущих древесный образ жизни. Однако цепкохвостые обезьяны, такие, как капуцины (Cebidae), ревуны (Alouattinae) и коаты (Atelinae), у которых хвост служит для опоры и хватания, имеют, подобно водным млекопитающим, мощные хвостовые позвонки с толстыми отростками для прикрепления мышц. У части зверей (собак (Canidae), копытных и т. д.) укороченный хвостовой отдел позвоночника служит только для отпугивания насекомых и подачи коммуникационных сигналов. Типичное строение при этом сохраняют лишь первые позвонки, а лежащие позади них принимают вид палочковидных тел. При полной утрате своих функций хвост в большей или меньшей степени редуцируется (зайцы (Leporidae), пищухи (Ochotonidae), слепыши (Spalacidae), медведи (Ursidae), многие приматы и др.).
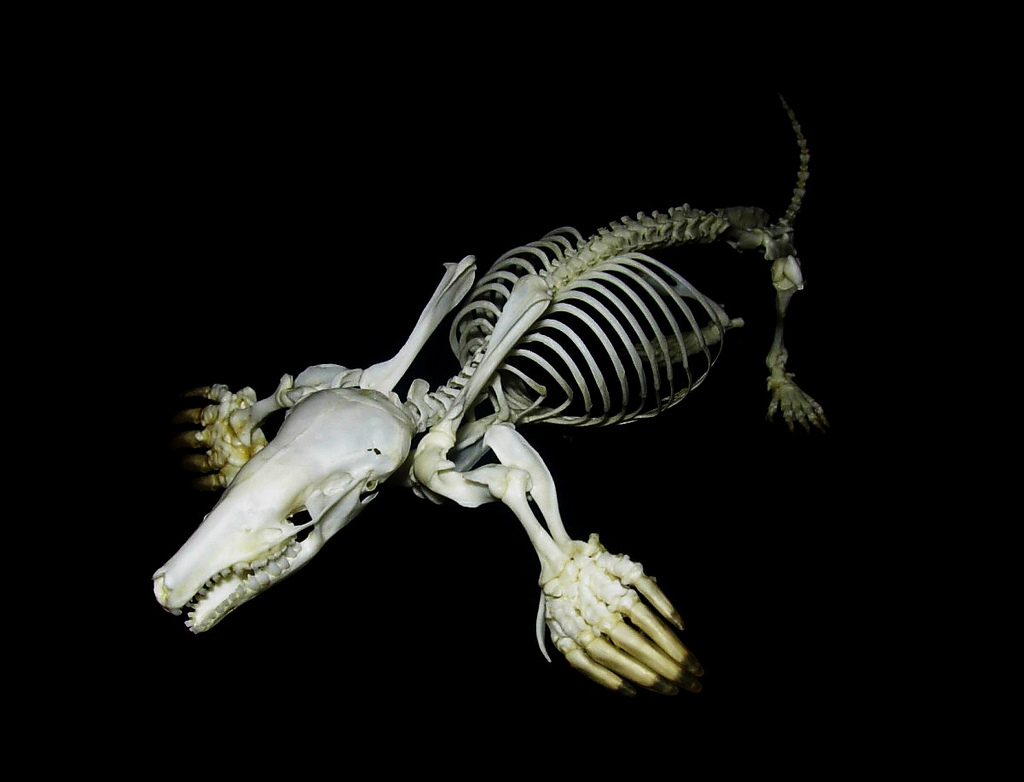
Смонтированный скелет обыкновенного крота (Talpa europaea).
Ребра
Ребра (costae) — парные дугообразные плоские кости. Подобно другим тетраподам, ребра млекопитающих имеют двойное сочленение с позвоночником: находящаяся на конце реберная головка (capitulum) причленяется между двумя соседними телами позвонков, а несколько отступающий от нее бугорок (tuberculum) — к нижней части поперечного отростка. У ребер однопроходных бугорков нет и они причленяются к позвонкам только посредством головок.
Шейные ребра у млекопитающих обычно не заметны, однако эмбриональное развитие показывает, что поперечные отростки шейных позвонков на самом деле включают приросшие снизу короткие двухголовчатые ребра; небольшое отверстие, пронизывающее отросток, представляет собой промежуток между собственно головкой и бугорком ребра. Из мезозойских групп свободные шейные ребра известны у могрганукодонтов (Morganucodonta), по крайней мере, у некоторых эвтриконодонтов (Eutriconodonta), симметродонтов (Symmetrodonta), многобугорчатых (Multituberculata), однако отсутствуют у эвгарамиид (Euharamiyida). Среди ныне живущих млекопитающих обособленные, хотя и неподвижные шейные ребра имеются лишь у однопроходных, у которых они синостозируют с поперечными отростками позвонков с образованием в них поперечного отверстия (foramen transversarium).
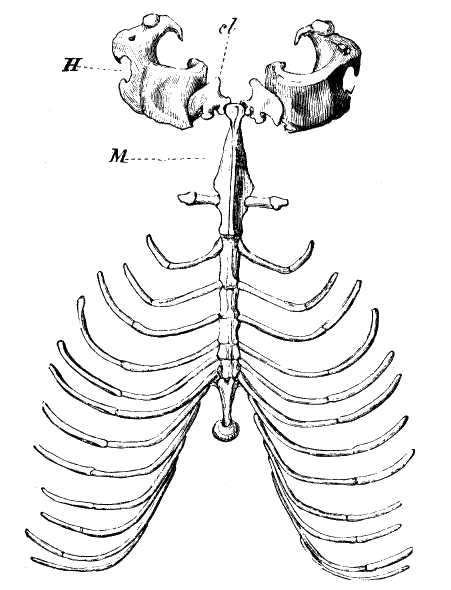
Часть плечевого пояса и ребра обыкновенного крота (Talpa europaea), вид снизу: H — плечо; cl — ключица; M — рукоятка грудины.
Наибольшего развития достигают «истинные» ребра (costae verae) грудного отдела. Их верхние концы крепятся к грудным позвонкам (обычно к первым 7), а нижние загибается вперед и через посредство реберных хрящей (cartilagines costales) соединяются с грудиной. У однопроходных и ксенартр реберные хрящи окостеневают. «Истинные» ребра и грудина образуют грудную клетку (thorax), которая обеспечивает дыхательные движения и служит защитой для органов грудной полости. Последующие грудные позвонки несут «ложные» ребра (costae spuriae), которые скреплены с последними «истинными» ребрами через хрящ или связочные соединения.
Позади предыдущих в грудном отделе могут иметься еще и «колеблющиеся» ребра (costae fluctuantes), заканчивающиеся свободно. У однопроходных и некоторых других древних групп ребра имеются также в поясничном отделе позвоночника. У более продвинутых млекопитающих они редуцируются и срастаются с поперечными отростками позвонков, которые вследствие этого называются плеврапофизами (pleurapophyses). В хвостовом отделе позвоночника ребер у млекопитающих никогда не бывает.

Смонтированный скелет гобиконодона (Gobiconodon ostromi).
Пояс передних конечностей
Пояс передних конечностей, или плечевой пояс, у млекопитающих упрощен и связан с осевым скелетом только мышцами и связками. Он образован грудиной, парными лопатками и ключицами.
Истинные ребра, путем слияния своих дистальных концов, дают начало грудной кости, или грудине (sternum), которая обычно узка и сильно вытянута. В грудине различают три следующих друг за другом отдела. Расширенный передний отдел, рукоятка (manubrium sternum) связан с первой парой ребер и ключицами (если таковые имеются). Средний отдел, или тело (mesosternum), состоит из нескольких сегментов, связывающих между собой концы соответствующих пар ребер. Наконец, в нижней части грудина заканчивается удлиненным мечевидным отростком (processus xiphoideus). У рукокрылых грудина теряет ясно выраженную сегментацию и в большинстве случаев несет в своей средней части небольшой килевой выступ, служащий, как и настоящий киль у птиц, для прикрепления мощных грудных мышц. У форм с хорошо развитыми для рытья передними конечностями (например, у кротов) на грудине развивается выполняющий ту же функцию маленький бугорок.
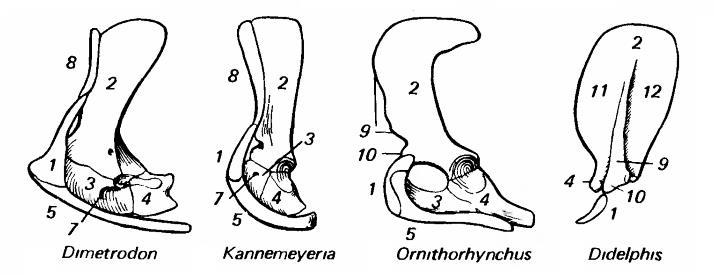
Плечевой пояс зверообразных (пеликозавр диметродон (Dimetrodon), дицинодонт каннемеерия (Kannemeyeria)) и млекопитающих (однопроходное утконос (Ornithorhynchus), сумчатое опоссум (Didelphis)): 1 — ключица (clavicula); 2 — лопатка (scapula); 3 — прокоракоид (procoracoideum); 4 — коракоид (coracoideum); 5 — межключица (interclaviсulа); 6 — грудина (sternum); 7 — коракоидное отверстие (foramen coracoideum); 8 — клейтрум (cleithrum); 9 — ость лопатки (spina scapulae); 10 — акромиальный отросток (processus acromialis); 11 — предостная ямка (fossa supraspinаtа); 12 — заостная ямка (fossa infrаspinаtа).
Основой плечевого пояса служат лопатки (scapulae) — расположенные дорсально плоские кости, форма и очертания которых крайне варьируют. Наиболее часто лопатка имеет вид вытянутого неправильного треугольника. Основа ее уплощена, а вдоль всей верхней части, ближе к наружному краю, тянется хорошо развитый продольный гребень — лопаточная ость (spina scapulae), увеличивающая поверхность для крепления мышц. Перед остью на лопатке находится предостная (fossa supraspinаtа), а за остью — заостная (fossa infrаspinаtа) ямки. Спереди ость лопатки заканчивается акромиальным отростком (processus acromialis), к которому подвижно причленяется ключица. Судя по связи с ключицей, лопаточная ость соответствует клейтруму (cleithrum), расположенному на переднем крае лопатки зверообразных. Месторасположение ости по переднему лопаточному краю сохранилось до наших дней у однопроходных. У териевых млекопитающих впереди ости располагается новоприобретенная часть лопатки — дополнительная площадь для прикрепления мышц. На суженном вентральном конце лопатки находится вогнутая суставная ямка для головки плечевой кости, а над ней возвышается искривленный коракоидный отросток (processus coracoideus).
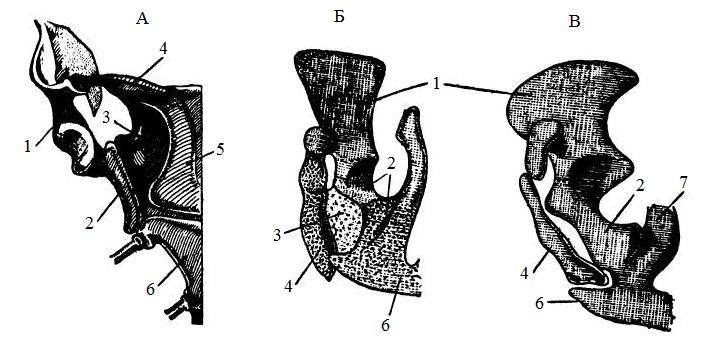
Правая половина плечевого пояса утконоса (Ornithorhynchus anatinus) (А), зародыша (Б) и детеныша (В) кузу (Trichosurus sp.), вид снизу: 1 — лопатка (scapula); 2 — коракоид (coracoideum); 3 — прокоракоид (procoracoideum); 4 — ключица (clavicula); 5 — межключица (interclavicula); 6 — грудина (sternum); 7 — ребро (costa).
Ключицы (claviculae) — расположенные вентрально относительно тонкие, слегка искривленные кости кожного происхождения. У многих генерализованных сумчатых и плацентарных хорошо выраженные ключицы подвижно сочленяются с рукояткой грудины. Значительного развития они достигают у форм, передние конечности которых приспособлены к движениям в различных плоскостях и у которых наличие ключицы обеспечивает более прочное причленение плечевой кости и упрочение всего плечевого пояса (кроты, рукокрылые, обезьяны). В то же время, ключицы обнаруживают тенденцию к редукции у бегающих и прыгающих форм, у которых полная свобода лопатки от осевого скелета нужна для ослабления толчков, передаваемых с передних конечностей на тело, и для увеличения функциональной длины конечности за счет поворотов лопатки. Наконец, они полностью утрачены у животных, передние конечности которых совершают только или преимущественно однообразные движения в параллельной главной оси тела плоскости. Ключицы неразвиты у бандикутов, зайцеобразных и некоторых грызунов; рудиментарны, заменены связками или отсутствуют у хищных; утрачиваются у копытных, китообразных, хоботных и сирен.
Смонтированный скелет большой панды (Ailuropoda melanoleuca).
Пояс задних конечностей
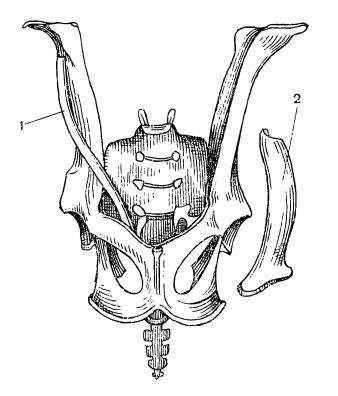
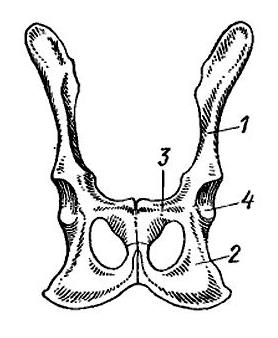
Пояс задних конечностей, или таз (pelvis), в эмбриональный период состоит из типичных для этого отдела 3 парных костей — подвздошной (ilium), седалищной (ischium) и лобковой (pubis). В месте их схождения находится вертлужная впадина (acetabulum) — большое округлое углубление, в которое входит головка бедренной кости. По сравнению с предшественниками, в тазовом поясе у прогрессивных зверообразных и млекопитающих произошли заметные изменения, связанные с изменением положения конечностей и преобразованием мускулатуры. Мышцы, а также кости таза, от которых они начинаются, заметно повернуты против часовой стрелки, если смотреть с левой стороны. Подвздошная кость, которая первично была направлена верхним концом назад, теперь на своем пути вверх к крестцу отклоняется далеко вперед. Лобковая и седалищные кости, напротив, сдвинулись назад. В передней части вертлужной впадины нередко развивается небольшое окостенение — т. н. вертлужная кость (асеtabulare). Подвздошная кость млекопитающих первично представляет собой довольно тонкий стержень треугольноrо сечения. У тяжеловесных копытных (например, у современных лошадей (Equidae), быков (Bovini), а также у слонов (Elephantidae)) и двуногих форм (например, у человека), у которых имеются особенно мощные ягодичные мышцы, идущие к бедру от подвздошной кости, последняя сильно расширяется. Запирательное отверстие для одноименного нерва, которое находится между ветвями лобковой и седалищной костей, на стадии прогрессивных зверообразных увеличивается и образует широкое запирательное окно (fenestra obturatoria).
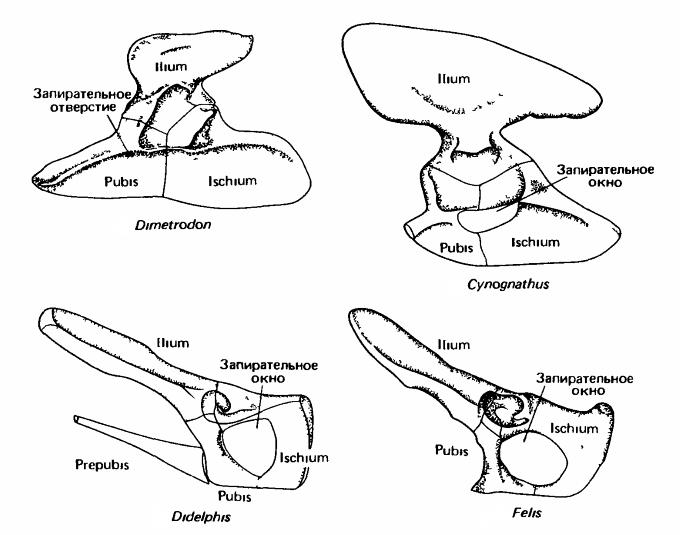
Тазовый пояс зверообразных (пеликозавр диметродон (Dimetrodon), цинодонт циногнат (Cynognathus)) и млекопитающих (сумчатое опоссум (Didelphis), плацентарное кошка (Felis)).
После завершения роста все тазовые кости обычно сливаются с каждой стороны в одну безымянную кость (innominatum). Внизу безымянные кости срастаются друг с другом по средней линии, формируя симфиз (symphysis pelvina) — обязательный атрибут закрытого таза. У большей части млекопитающих широким сращением соединены лобковый и седалищный отделы; у броненосцев, приматов и большинства насекомоядных (например, кротов (Talpidae) и землероек (Soricidae)) вентральный симфиз отсутствует и сращение имеется лишь в передней, лобковой части тазового пояса. Вытянутые подвздошные кости связаны синдесмозом (прослойкой соединительной ткани) с двумя истинно крестцовыми позвонками. Таз и крестец образуют костное кольцо, ограничивающее тазовое выходное отверстие, через которое, прежде чем покинуть тело, проходит содержимое половой, выделительной и пищеварительной систем. Размеры выходного отверстия являются важным параметром живородящих форм, приносящих крупных детенышей, в связи с чем оно часто шире именно у самок. Кроме того, в некоторых случаях симфиз ко времени родов разрыхляется под воздействием гормонов (главным образом между лобковыми костями).
Тазовый пояс сирен и китообразных в значительной мере редуцирован, у второй группы он даже утрачивает связь с позвоночником. При этом остаток таза китообразных связан с анальной мускулатурой и половыми органами, представляя собой вполне функциональную структуру.

Слева — расположение таза у сейвала (Balaenoptera borealis).
Справа, сверху вниз — таз кашалота (Physeter macrocephalus), сейвала (Balaenoptera borealis) и финвала (Balaenoptera physalus); у последнего также имеется рудимент бедра.
У ряда мезозойских групп (тритилодонты (Tritylodontidae), многобугорчатые (Multituberculata) и др.), а также у однопроходных и метатериев к лобковым костям подвижно причленяется пара предлобковых костей (prepubica), которые тянутся свободными концами вперед и расположены в брюшной стенке. Сначала предполагалось, что предлобковые элементы дают опору сумке для детеныша, из-за чего они и были названы сумчатыми костями (marsupialia). Затем, однако, было показано, что, сообщаясь с некоторыми мышцами бедра и живота, эти кости играют важную роль в локомоции. С развитием сумки они напрямую не связаны, поскольку имеются у представителей обоих полов и у таксонов, у которых сумка вообще отсутствует. В то же время, в строении предлобковых костей наблюдается половой диморфизм, так что их дополнительной функцией у самок является обеспечение поддержки при переносе массы детенышей. С другой стороны, эти кости рудиментарны и представлены волокнистыми хрящами у некоторых сумчатых, обладающих хорошо выраженными сумками — например, у сумчатого волка (Thylacinus cynocephalus) и сумчатого крота (Notoryctes typhlops).

Предлобковые элементы также присутствуют у базальных эвтериев и, по некоторым данным, у ряда низкоорганизованных современных плацентарных. В последнем случае они имею вид небольших хрящиков и занимают соответствующее положение по отношению к мускулатуре. У более продвинутых форм эти хрящи исчезают. Согласно другому предположению, редуцированные предлобковые элементы сливаются друг с другом и образуют непарные кости полового члена и клитора, имеющиеся у некоторых плацентарных; впрочем, доказательств у этой точки зрения весьма немного.
вперед
в оглавление