Строение и функции
Внешний облик
Сумчатые кроты — это единственные ныне живущие сумчатые, приспособленные к роющему образу жизни. Размеры их мелкие. Длина головы и туловища составляет 7-14 см, хвоста — 1,2-2,6 см. Вес варьирует в пределах 30-70 г. Половой диморфизм в размерах не выражен.

Большой сумчатый крот (Notoryctes typhlops).
Голова короткая и массивная, конусовидной формы. Рот находится на нижней ее поверхности. Язык может высовываться изо рта по меньшей мере на 2 см. Ноздри небольшие, щелевидные, расположенные вертикально. Коричневато-розовая голая кожа на конце мордочки (ринариум) прямо над ноздрями переходит в желтый щиток, покрывающий верхнюю часть носа до межглазничной области и примерно посередине разделенный поперечной бороздой. Он образован сильно утолщенным эпидермисом, в то время как дерма под ним тоньше, чем на других частях тела (за исключением, разве что, так же покрытого плотным эпидермисом хвоста). Твердый роговой щиток помогает раздвигать головой почву. Рудиментарные глаза скрыты под кожей, через которую немного просвечивают в виде темных пятнышек. Наружные слуховые отверстия имеют диаметр до 2 мм и окружены плотным шерстным покровом, ушные раковины отсутствуют. Вибриссов также нет.
Шея короткая и внешне неразличимая. Туловище вальковатое, несколько удлиненное и сплюснутое сверху. Наивысшей точкой тела является слегка выгнутая спина, голова и задняя часть туловища наклонены вниз. В области плечевого и тазового поясов имеются скопления подкожного жира. Короткий цилиндрический хвост покрыт кольцевыми чешуями, сформированными толстым эпидермисом, сильнее выраженными сверху и по бокам и уменьшающимися по окружности от основания к концу. Последний хвостовой позвонок несет несколько расширенный тупой ороговевший колпачок. Весь хвост жесткий.

Большой сумчатый крот (Notoryctes typhlops).
Конечности хорошо приспособлены для рытья. Они короткие и мощные, особенно сильно укорочена передняя пара. Широкие лапы имеют по 5 пальцев, в значительной мере редуцированных, но между собой не сращенных. Когти развиты неравномерно. На кисти когти I-го и II-го пальцев длинные, тонкие и узкие. Когти III-го и IV-го пальцев очень сильно увеличены, уплощенные и с притупленными кончиками. Коготь III-го пальца самый крупный, длиной около 15 мм. Коготь IV-го пальца по сравнению с предыдущим короче, но шире у основания, так что приобретает треугольную форму. V-й палец оснащен маленьким расширенным когтем. Когти III-го и IV-го пальцев настолько большие, что закрывают собой всю остальную кисть. У внутренней стороны этих пальцев, отделенная от них глубокой бороздкой, находится твердая, морщинистая ладонь, из которой выступают когти I-го и II-го пальцев. Таким образом, пальцы кисти составляют 2 ряда со щелью между ними. Дорзальная поверхность кисти обращена внутрь. На стопе II-й, III-й и IV-й пальцы несут удлиненные, а короткие I-й и V-й пальцы — небольшие плоские когти. На проксимальной фаланге I-го пальца развита хрящевая лопасть, увеличивающая площадь стопы. Она настолько вывернута в передне-наружном направлении, что V-й палец оказывается впереди и подошвенная поверхность обращена наружу.
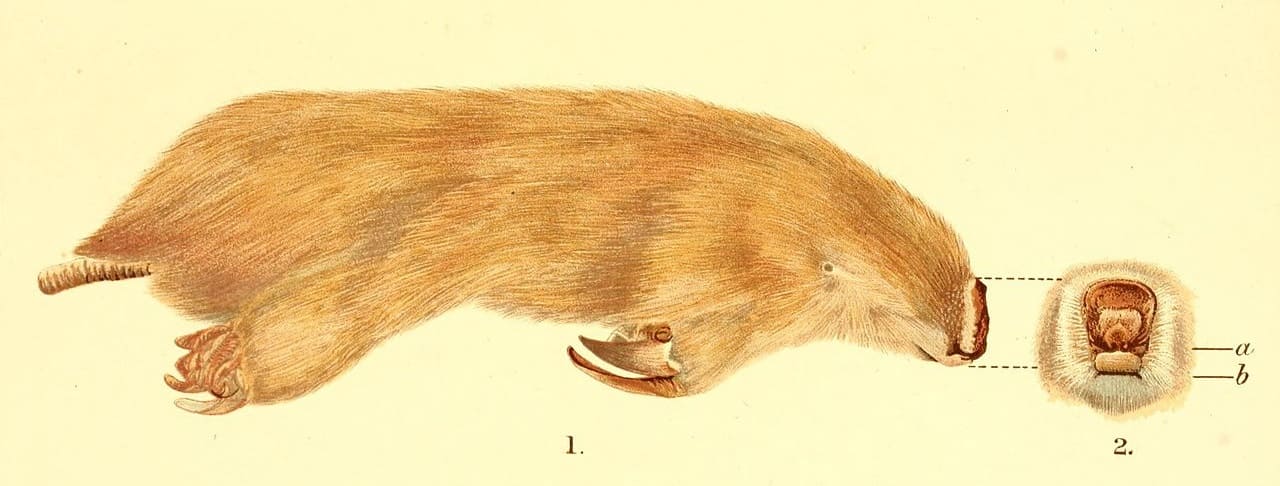
Внешний облик большого сумчатого крота (Notoryctes typhlops), сбоку и спереди.
Волосяной покров сравнительно короткий, очень густой и мягкий. Такая структура меха способствует продвижению в толще грунта. В зависимости от места обитания его окраска варьирует от почти белой до розовато-светло-коричневой и насыщенной желтовато-красной, с характерным переливающимся отблеском. Красновато-коричневый оттенок шерсти придают оксиды железа, которыми богата австралийская почва. При содержании в террариуме без соответствующего грунта красноватый окрас вскоре утрачивается, но вновь приобретается после его внесения. Вполне возможно, что изменчивость меховой окраски имеет маскировочное значение.
Волосы сгруппированы в небольшие пучки, каждый из которых растет из собственного фолликула. Пучок содержит 1 длинную ость и от 9 до 20 пуховых волос. 3-5 пучков образуют группу. Наиболее плотно волосяные пучки располагаются на подбородке, по сторонам головы и на спине в крестцовой области. Именно в нижней части крестца имеется т. н. ишиотергальное пятно диаметром около 10 мм, на котором волосы обычно сильнее спутаны и темнее окрашены. На конечностях волосы растут только до кисти и ступни, на хвосте они также отсутствуют. Отверстие клоаки окружено одиночными крупными волосками. Сальные железы сопровождают волосы по всему телу. Они в меньшей степени развиты на плече и бедре, и в большей — на верхней части головы, ишиотергальном пятне, в сумке и вокруг клоаки.
Скелет
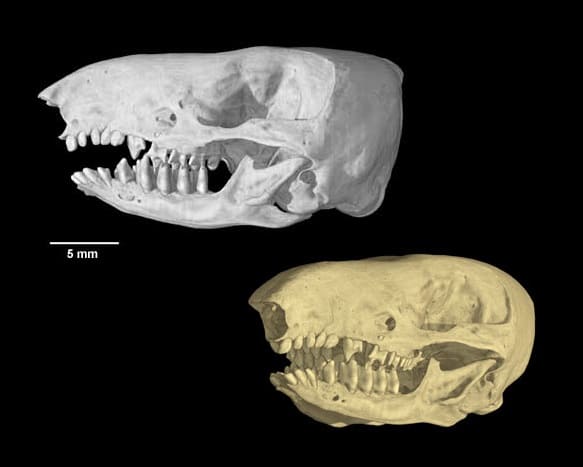
Компьютерные модели черепа с нижней челюстью большого сумчатого крота (Notoryctes typhlops).
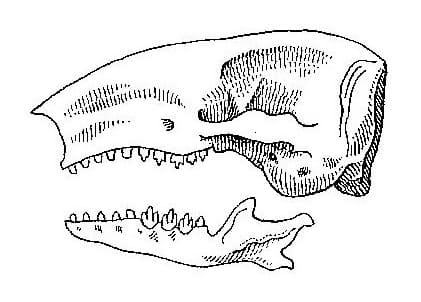
Череп конической формы со суживающейся мордой и расширенным мозговым отделом. От затылочной линии кпереди до начала носовых костей верхняя поверхность черепа плоская, а на носовых костях скошена вниз под углом приблизительно 60 °. Носовые кости клинообразно выдаются вперед и вверх, нависая над носовым отверстием. Глазничное отверстие и заглазничная дуга полностью отсутствуют. Имеется межглазничное сужение. Скуловая дуга сильно развита, максимальную ширину череп имеет в месте ее подъема на скуловой кости. Вздутый слуховой барабан лучше всего заметен при взгляде сбоку. Швы между костями мозгового отдела исчезают рано. Стреловидного гребня нет. 2 низких гребешка отходят от задней вершины средней линии черепа и дугообразно спускаются вниз к началу задней части скуловой дуги. Задняя часть черепа, его затылочная область, плоская и поднимается вертикально к затылочной линии. Большое затылочное отверстия расположено посредине задне-нижнего края черепа и наиболее хорошо заметно при взгляде снизу.
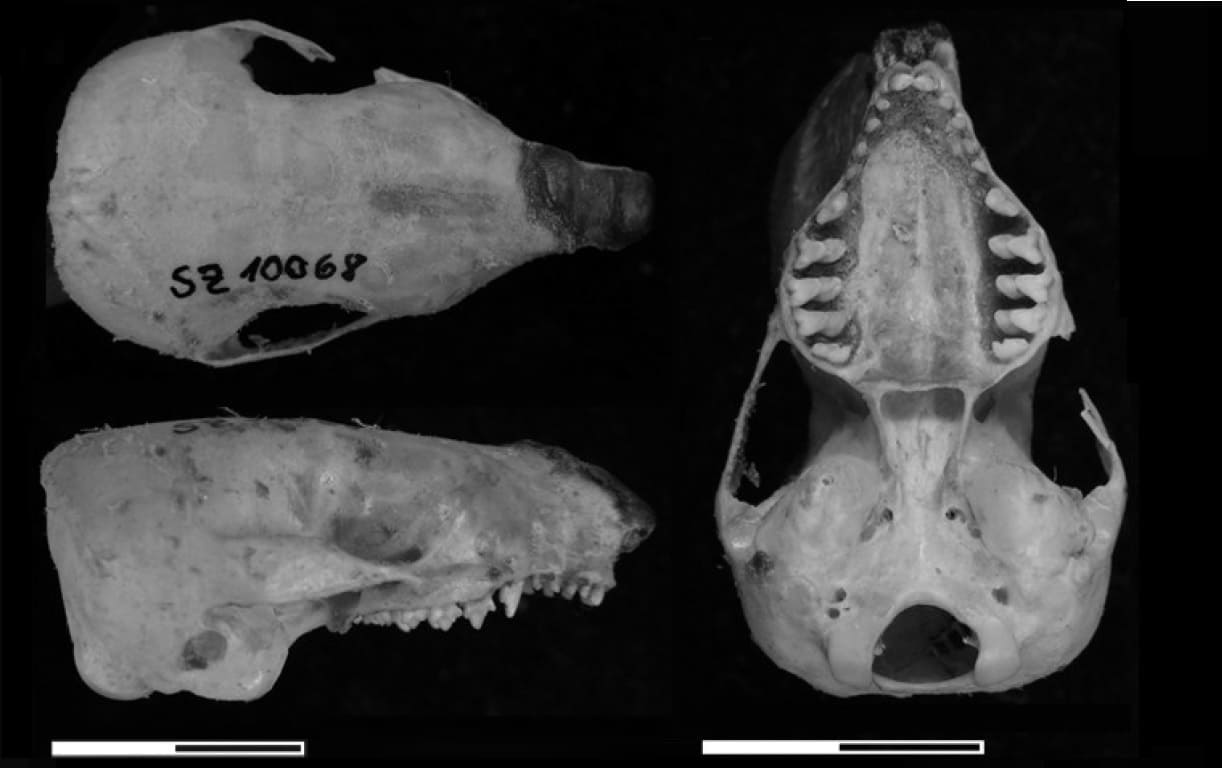
Череп большого сумчатого крота (Notoryctes typhlops). Длина масштабной линейки — 1 см.
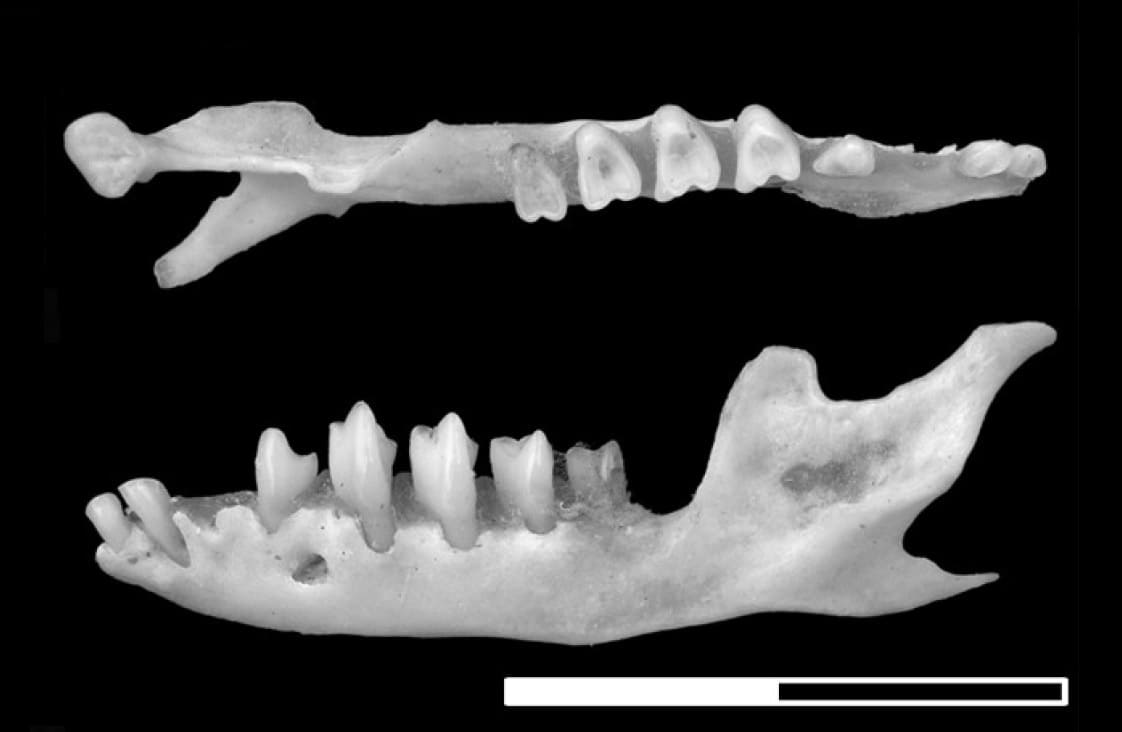
Нижняя челюсть большого сумчатого крота (Notoryctes typhlops). Длина масштабной линейки — 1 см.
Нижняя челюсть за симфизом заметно расширяется. Угловой отросток на конце сужается в точку и загнут внутрь. Суставный и венечный отростки соединены в нижней половине своей длины. На внешней поверхности этого объединения имеется хорошо заметная ямка для прикрепления жевательных мышц. Венечный отросток поднимается почти вертикально, отклоняясь назад и внутрь лишь незначительно. Суставный отросток отклоняется назад под углом около 60 ° и заканчивается расширенной и уплощенной сверху суставной головкой, лишь незначительно смещенной в боковом направлении относительно передне-задней оси зубного ряда.
Прорисовки черепа и нижней челюсти большого сумчатого крота (Notoryctes typhlops), сбоку и сверху.
Зубная формула довольно изменчива: I3-4/i2-4, C1/c1, P2-3/p2-3, M3-4/m3-4 = 38-48 (обычно 42-44). Число зубов в разных половинах челюстей может не совпадать даже у одной и той же особи. Все зубы четко отделены друг от друга. Резцы, клыки и большинство предкоренных имеют различные, но всегда небольшие размеры, простую колышковидную форму и обычно тупые жевательные поверхности со слитыми параконом и метаконом. Только последний верхний предкоренной более крупный и несет 2 бугорка. Крупные верхние коренные трехбугорчатые, залямбдодонтного типа, с отсутствующим талонидом и сохранившимся протоконом. Нижние коренные более мелкие, с маленьким талонидом и утраченным талонидным бассейном. Поскольку у щечных зубов корни составляют лишь около 1/3 их длины, допускается, что слишком твердые пищевые частицы в рацион входить не могут.
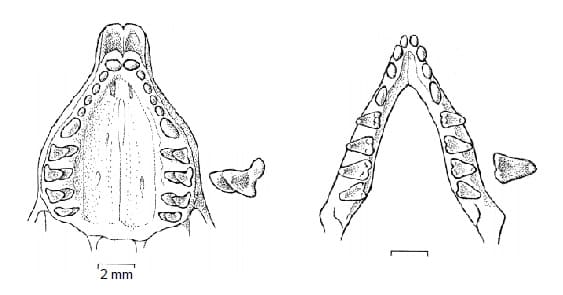
Прорисовки жевательных поверхностей верхнего и нижнего зубных рядов большого сумчатого крота (Notoryctes typhlops).
Позвонков 7 шейных, 15 грудных, 4 поясничных, 6 крестцовых и 12-13 хвостовых. Шейные позвонки с II-го по V-й сращены. Атлант имеет обычное строение, за исключением круглого сочленения вверху и посредине задней поверхности невральной дуги. Зубовидный отросток эпистрофея хорошо выражен. 5 сросшихся позвонков имеют небольшие невральные дуги и остистые отростки, а также несут короткий направленный вперед отросток, соединяющийся с суставной поверхностью на задней стороне невральной дуги атланта. Хотя голова способна вращаться вокруг оси и наклоняться, верхнее сочленение между атлантом и эпистрофеем препятствует ее подъему выше горизонтальной плоскости. Шея изгибается назад и вниз под таким углом, что ее VI-й позвонок сзади сочленяется не только с VII-м шейным позвонком, но и с передним ребром. У VII-го шейного позвонка нижний прогиб преобразован в верхнюю дугу.

Смонтированный скелет большого сумчатого крота (Notoryctes typhlops).
I-й грудной позвонок несет очень длинный остистый отросток и короткое, но при этом массивное ребро, сочленяющееся с VII-м шейным позвонком, а снизу — с крупной рукояткой грудины. Грудные позвонки с I-го по VI-й имеют большие остистые отростки, из которых самый длинный находится на I-м позвонке. Позвоночник выгибается вверх от сочленения VII-го шейного с I-м грудным и до VI-го грудного позвонка, затем идет горизонтально до конца крестцового отдела, а с I-го хвостового позвонка начинает опускаться вниз. Поперечные отростки IV-го поясничного позвонка образуют подвижное сочленение с поперечными отростками I-го крестцового позвонка и подвздошной костю. Крестцовые позвонки сращены вместе, а V-й и VI-й позвонки поперечными отростками прирастают к седалищной кости. Хвостовые позвонки с I-го по IX-й хорошо сформированы, но дальше становятся трудноразличимыми, числом от 3 до 4. На середине хвоста имеются 2 боковых выступа, так что он здесь шире, чем у корня.

Смонтированный скелет большого сумчатого крота (Notoryctes typhlops).
Ребер бывает 14 либо 15 пар. Ребра с I-го по VII-е сочленяются непосредственно с грудиной. Остальные ребра посредством хрящей соединены каждое со своим предшественником. I-е ребро короткое, совершенно окостеневшее и прочно сочлененное с VI-м шейным, I-м грудным позвонками и рукояткой грудины. Последняя представляет собой массивную щитообразную конструкцию с хорошо выраженным направленным вперед килем. По внешнему виду рукоятка сходна с таковой у некоторых летучих мышей (Microchiroptera). Стержневидное тело грудины состоит из 5-6 сегментов. Мечевидный отросток заканчивается плоским хрящевым расширением.
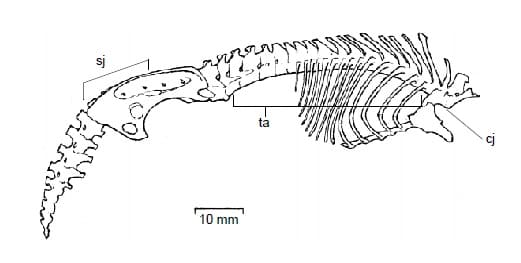
Прорисовка позвоночного столба большого сумчатого крота (Notoryctes typhlops): cj — шейно-грудной переход; ta — грудопоясничный изгиб; sj — крестцово-каудальное сочленение.
Ключица представляет собой тонкую изогнутую кость, слегка расширяющуюся на сближенном с лопаткой конце. К грудине она присоединяется посредством связок, прикрепленных к передне-боковому краю ее рукоятки.
Строение лопатки уникально. Из места расположения суставной ямки, коракоида и подмышечного края к позвоночному краю лопатки по ее верхней стороне тянутся V-образные возвышения. На протяжении нижних 2/3 лопатки эти гребни превосходят по высоте ее ширину. Большая часть поверхностей для прикрепления мышц располагается именно вдоль костных возвышений, а не в надостной и подостной ямках. Между суставной ямкой и акромиальным отростком расположена глубокая узкая выемка. От вынесенного вперед акромиального отростка выше уровня суставной ямки поднимается гребень. Подмышечный край лопатки поворачивает вверх вдоль выемки, увеличиваясь по высоте к суставной ямке и формирует еще один гребень. Верхушки обоих гребней так тесно сближены, что пространство между ними почти замыкается в трубку. Позвоночный край лопатки изогнут, утолщен и слегка приподнят.
Плечевая кость является коротким, массивным элементом. Она несет много более или менее выраженных бугров, а также большой гребень для прикрепления дельтовидных и других мышц плеча, простирающийся на половину ее длины. Внутренний надмыщелок выступает из дистального конца плечевой кости на расстояние, примерно равное половине ее длины. Длинный, изогнутый локтевой отросток почти равен по длине телу локтевой кости. Лучевая кость по длине также почти равняется телу локтевой кости. Кисть пятипалая. I-й, II-й и III-й пальцы имеют по 2 фаланги, а IV-й и V-й — по 1. В дистальном отделе I-й пястной кости находится большая сесамовидная косточка.
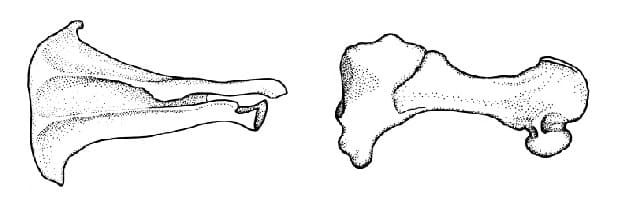
Прорисовки лопатки (слева) и плечевой кости (справа) большого сумчатого крота (Notoryctes typhlops), снаружи.
Таз расположен почти горизонтально и сориентирован параллельно продольной оси грудопоясничной части позвоночного столба. Подвздошная кость спереди доходит до уровня IV-го поясничного позвонка. Сверху слившаяся с отростками крестцовых позвонков тазовая кость расширяется в боковом направлении и формирует арочный свод. Спереди он достигает IV-го поясничного позвонка и заканчивается под его поперечным отростком. Сзади свод сливается с седалищной областью и на уровне поперечного отростка II-го хвостового позвонка расширяется в задне-боковом направлении. Сверху тонкая подвздошная кость полностью накрывает пещеристую область, лежащую между ней и арочным сводом. Запирательное отверстие круглое. Вертлужная впадина расположена впереди и выше запирательного отверстия и имеет сходный с ним размер. Несмотря на комплектность вертлужной впадины, ее передне-верхняя стенка утолщена и поднята значительно выше остального края впадины, ориентируя ее в задне-нижнем направлении. Предлобковые (сумчатые) кости редуцированы до коротких и тонких стержней, составленных волокнистыми хрящами или окостеневающих.
Головка бедренной кости представляет собой овальное утолщение на проксимальном эпифизе, выраженная шейка у нее отсутствует. Отчетливо выраженный малый вертлуг расположен дистальнее головки. Большой вертлуг вздымается выше уровня головки, полого опускаестся спереди назад и тянется в виде отчетливо различимого гребня на протяжении проксимальной 1/4 тела кости. Дистальная суставная поверхность бедра вполне обычна, за исключением того, что мыщелковые поверхности расширяются кпереди для сочленения с очень большой коленной чашечкой. Большеберцовая кость уплощается в задне-переднем направлении с образованием на ее передней грани хорошо заметного гребня, который тянется, дистально понижаясь, на протяжении проксимальных 2/3 тела кости. Проксимальной гребень образует обширное сочленение с большой коленной чашечкой. Проксимальный конец малоберцовой кости расширяется спереди назад и может свободно сочленяться с направленным вверх отростком коленной чашечки. Малоберцовая кость уступает по массивности большеберцовой, но примерно равна ей по длине. Стопа построена из вполне обычных костных элементов. Имеется 5 пальцев. Как и в кисти, в области пясти присутствует большая сесамовидная косточка. Эта косточка совместно с расширенной V-й пястной костью образует паз. В стопе синдактилии (сращения пальцев) не наблюдается.
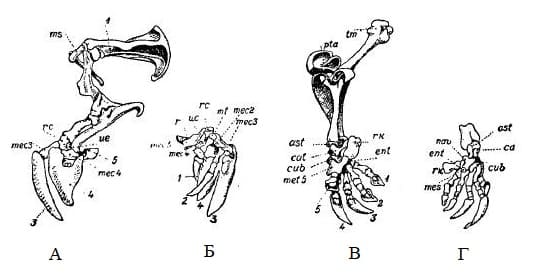
Прорисовки скелета передней (А, Б) и задней (В, Г) конечностей большого сумчатого крота (Notoryctes typhlops), с наружной (А, В) и внутренней (Б, Г) стороны: rc — лучевая кость; uc — локтевая кость; mt — запястная кость; mec — пястные кости; tm — большой вертлуг; ast — таранная кость; cat — пяточная кость; nav — ладьевидная кость; mes — заплюсневая кость; met — плюсневые кости; 1-5 — пальцы.
Таким образом, вся скелетная система сумчатых кротов является прекрасно приспособленной к роющему образу жизни. При поступательных движениях в толще грунта давление распространяется кзади от головы, в связи с чем компрессионному сжатию подвержены позвоночный столб и грудная клетка. Часть шейных позвонков срослась для усиления жесткости шеи во время рятья. Хорошо развитые остистые отростки позвонков грудопоясничной области служат для прикрепления глубоких сильных мышц, облегчающих продвижение вперед посредством сгибания позвоночника в поясничном отделе. Обширные поверхности для прикрепления мышц на тазовых костях, сверху бедренной кости, коленной чашечке и большеберцовой кости также обеспечивают мощный толчок в переднем направлении. Плечевой пояс с килеватой грудиной и трубчатой лопаткой демонстрирует максимальный уровень специализации. Расширенная кисть отлично приспособлена для рытья, а стопа служит для отбрасывания накопанной почвы назад. Жесткий хвост способен упираться в субстрат, выполняяя роль упора для позвоночника и всего тела.
Мускулатура
Среди характерных миологических признаков можно выделить следующие.
Большая подкожная мышца (m. panniculus carnosus) развита слабо. Наличие мускульных прикреплений в задне-наружной части седалищной области и на пояснице обусловливает возможность ишиотергального скольжения. Насколько известно, эта особенность присуща исключительно сумчатым кротам.
Остисто-трапециевидная мышца (m. spinotrapezius) обоими своими концами прикрепляется независимо от других порций трапециевидной мышцы (m. trapezius). Ромбовидная мышца груди (m. rhomboideus thoracis) представляет собой толстый и широкий мускульный пласт. Часть ее отходит от длинных невральных отростков I-го и II-го грудных позвонков, также имеется очень широкое прикрепление к позвоночному краю лопатки, а часть волокон тянется вперед для прикрепления на преаксиальном расширении лопаточной ости.
Среди мышц ключично-грудинно-черепной области отсутствует ключично-сосцевидная мышца (m. cleidomastoideus). В то же время, имеется грудинно-сосцевидная мышца (m. sternomastoideus), начинающаяся от переднего края рукоятки грудины. Ключично-дельтовидная мышца (m. clavodeltoideus) мала, наличие ключично-трапециевидной мышцы (m. clavotrapezius) несколько сомнительно. Значительное развитие крепящейся к ключице и связанной с ней мускулатуры объясняется главным образом сильным расширением передней конечности в латеральной плоскости.
Широчайшая мышца спины (m. latissimus dorsi) прикрепляется исключительно к остистым отросткам грудных позвонков с III-го по XII-й. Вместо плечевой кости она заканчивается посередине предплечья на апоневрозе локтевого сгибателя запястья (m. flexor carpi ulnaris).
Хорошо развитая большая грудная мышца (m. pectoralis major) делится на 3 части. 2 поверхностные передние порции начинаются на рукоятке и теле грудины и прикрепляются к вентральной поверхности массивного дельтовидного гребня плечевой кости. III-я, более глубоко расположенная и отодвинутая кзади порция, берет начало на вентральных поверхностях ребер и прикрепляется к большой бугристости плечевой кости.
Большая круглая мышца (m. teres major) не связана с широчайшей мышцей спины; малая круглая (m. teres minor) и клювовидно-плечевая (m. coracobranchialis) мышцы отсутствуют. Подостная мышца (m. infraspinatus) проходит через глубокую канавку, образуванную костным гребнем и полукружно изогнутым подмышечным краем лопатки.
В трехглавой мышце плеча (m. triceps brachii) ее длинная головка (cap. longuin) очень широким сухожилием начинается от подмышечного края лопатки на всей его протяженности, а также от дополнительного гребня, сформированного по этому краю. Латеральная (caput laterale) и медиальная (caput mediate) головки отходят от тела плечевой кости по всей его длине. Все головки прикрепляются к большой выпуклой поверхности локтевого отростка. По-видимому, часть длинной головки вместе с широчайшей мышцей спины заканчивается на предплечье.
Мускулатура предплечья и кисти претерпела существенные изменения. Сильно развитый короткий лучевой разгибатель запястья (m. extensor carpi radialis brevis) прикрепляется к проксимальным 3/5 лучевой кости. Круглый пронатор (m. pronator teres) также хорошо развит, тогда как квадратный пронатор (m. pronator quadratus) отсутствует. Мощный локтевой сгибатель запястья (m. flexor carpi ulnaris) покрыт прочным апоневрозом, к которому прикрепляется широчайшая мышца спины.
Вся внутрення поверхность вогнутости локтевого отростка и пространство внутри нее занято глубоким сгибателем пальцев (m. flexor digitorum profundus). Эта мышца тянется вниз по медиальной половине тела локтевой кости и прикрепляется к большой сесамовидной косточке, находящейся в дистальном отделе I-й пястной кости. Из внутренних мышц кисти сохраняется ороткая мышца, отводящая большой палец (m. abductor pollicis brevis), короткий сгибатель большого пальца (m. flexor brevis pollicis) и короткий сгибатель указательного пальца (m. flexor brevis digiti secundi).
Мускулатура тазовой области и задних конечностей остается практически не изученной.
Пищеварительный тракт
Пищевод имеет короткую задиафрагмальную часть, входящую в желудок посредине его малой кривизны. Желудок имеет наибольшую длину 17,5 мм и наибольшую ширину 12,5 мм, но может изрядно сжиматься. Длина кишечного тракта от пилорического клапана до отверстия клоаки составляет около 290 мм. Передняя его часть достигает в диаметре 5 мм, тогда как задняя 3 мм. Выраженная граница между тонким и толстым кишечником отсутствует. Примерно за 100 мм до клоаки находится мешковидное расширение с подобием клапана, но является ли оно слепой кишкой, неясно.
Поджелудочная железа, по-видимому, занимает обычное положение, протянувшись вдоль большой кривизны желудка и маленькой части кишки сразу за его привратником. Печень делится на 2 больших и 2 маленьких доли; большие доли лежат вентральнее меньших. Желчного пузыря нет. Общий желчный проток открывается в тонкий кишечник 2,5 мм ниже пилорического клапана.
Кровообращение
Сердце занимает обычное положение, его достаточно острая верхушка направлена влево. Дорсально сердце отделено от диафрагмы частью правого легкого, но вентрально перикард связывается непосредственно с диафрагмой. Правый желудочек не участвует в образовании верхушки сердца. Легочная артерия выходит из правого желудочка и представляет собой короткий, толстый сосуд, почти сразу разветвляющийся на левую и правую легочные артерии. Правая ветвь короче и шире, чем левая.
Аорта выходит из левого желудочка примерно на том же уровне, что и легочный ствол из правого. Дугообразно изгибаясь вперед и влево, аорта продолжается в заднем направлении по обычной для млекопитающих схеме.
В районе аортальной дуги, однако, наблюдаются индивидуальные различия в ответвлении сонных и подключичных артерий. В одном случае 2 сонные артерии выходят из общего короткого ствола, от основания которого также ответвляется правая подключичная артерия, а левая подключичная артерия отходит от дуги аорты несколько дальше и левее. В другом случае безымянный ствол разделяется на правую сонную и подключичную артерии, левая сонная артерия ответвляется недалеко от основания этого ствола, а левая подключичная артерия, как и в предыдущем варианте, отходит от дуги аорты дальше и левее.
Еще одной особенностью артериальной системы является то, что средняя крестцовая артерия ответвляется незадолго до разделения брюшной аорты на 2 общие подвздошные артерии. Средняя крестцовая артерия может быть представлена 1-3 небольшими сосудами или полностью отсутствовать. В обычной для большинства млекопитающих схеме, средняя крестцовая артерия представляет собой небольшую ветвь брюшной аорты.
2 легочные вены объединяются в общий ствол, равный легочному артериальному стволу по толщине, но примерно вдвое превосходящий его в длину. Легочный ствол входит в левое предсердие. 3 полые вены — задняя, левая и правая — входят в тесное соприкосновение до впадения в правое предсердие, но при этом сохраняют отдельные выходящие отверстия.
Остальная часть венозной системы соответствует обычной для млекопитающих схеме. И венозные и артериальные сосуды, обслуживающие передние конечности, в достаточной мере расширены. Селезенка занимает стандартное положение и имеет V-образную форму.
Органы дыхания и выделения
Легкие сумчатых кротов имеют большие размеры, общее их строение вполне обычно.
Почки субовальной формы с гладкой внешней поверхностью. Надпочечник расположен на передне-внутренней стороне. Мочеточники отходят от почечных ворот и открываются на вентральной поверхности мочевого пузыря, очень близко от его выводного протока. У самцов мочеполовой канал открывается длинной щелью на верхней стороне пениса. У самок мочеиспускательный канал соответствует типичной для сумчатых схеме.
Никаких особенных механизмов для экономного расходования воды, которые встречаются у большинства обитателей пустынь, у сумчатых кротов не выявлено.
Работа эндокринных желез практически не изучена.
Репродуктивная система
Выводковая сумка у самок небольшая, но хорошо сформированная. Она обращена назад, что препятствует попаданию внутрь частиц почвы. Складка в нижней части брюха разделяет сумку неполной перегородкой на 2 "кармана", в каждом из которых находится по 1 соску. Самцы обладают рудиментом выводковой сумки, образованным небольшой поперечной складкой кожи. Сальные железы в сумке хорошо развиты, особенно те из них, которые расположены вокруг сосков.
У особей обоих полов имеется рудиментарная клоака. Мочеполовой синус отделен от анального отверстия в большей степени, чем у однопроходных, но не столь выраженно, как у териев, и, в отличие от последних, является неразделенным. Слизистая оболочка клоаки, особенно вблизи ее наружного отверстия, весьма богата железами, вероятно апокриновыми по своей природе. Как и у однопроходных, обычных кожных желез и связанных с ними проток клоака лишена. По бокам от передней половины клоаки располагаются парные анальные железы. Каждая из них представляет собой округлое тельце диаметром 2,4 мм, находящееся в фиброзной капсуле и окруженное поперечно-полосатых мышечными волокнами. Секрет мелких апокриновых структур собирается в центральной полости железы и выбрасывается в клоаку через соединяющий их проток.
Подобно однопроходным и в отличие от остальных современных териев, пенис у самцов находится внутри клоаки. Семенники овальной формы расположены примерно на уровне переднего края лобковой кости, т. е. впереди клоаки. В отличие от остальных сумчатых, мошонка отсутствует, но, вопреки частому утверждению, семенники не лежат в брюшной полости, а находятся между кожей и брюшной стенкой. Семявыносящие протоки и семенные кровеносные сосуды через паховые кольца выходят в брюшную полость. Семявыносящие протоки впадают в мочеполовой канал примерно на том же уровне, что и мочеточники впадают в мочевой пузырь. Приблизительно на этом же уровне расположена железистая масса, по-видимому, представляющая собой простату. В основании пениса имеется пара бульбоуретральных желез. Многочисленные мелкие железки, характерные для мочеполового канала териев, отсутствуют.
Нервная система
Головной мозг устроен крайне примитивно даже для сумчатых. Он выраженно макросматический. Обонятельные луковицы очень крупные — больше, чем у однопроходных. В отличие от териев и подобно однопроходным, обонятельные луковицы смещены от средней линии вентро-латерально. Обонятельные бугры тоже развиты очень хорошо, но уступают по размерам луковицам. И те и другие представляет собой разграниченные, отчетливо выраженные структуры. Расположенный позади них средний мозг своеобразно сжат в передне-заднем направлении, так что он вздыбливается вертикально по отношению к остальному мозгу.
Полушария переднего мозга имеют большую грушевидную долю, отвечающую за распознавание запахов. Наоборот, новая кора развита меньше, чем у любого другого современного млекопитающего. Она лежит поверх грушевидной доли и совсем не имеет извилин, что также не характерно для териев, но типично для однопроходных.
Сплюснутый сверху вниз листообразный мозжечок полностью покрывает заднюю выпуклость нижнего двухолмия среднего мозга. Мозжечок также сжат в задне-переднем направлении, равно как и продолговатый мозг, так что последний достаточно четко отграничен от спинного мозга. Эпифиз расположен сверху посредине, на III-м желудочке головного мозга.
От нижней поверхности головного мозга отходят очень большие корни V-й пары черепных нервов. Гипофиз расположен на средней линии между обонятельными буграми. Какие-либо следы II-й, III-й, IV-й и VI-й пар черепных нервов отсутствуют. VII-я, VIII-я, IX-я, X-я, XI-я и XII-я пары черепных нервов имеются. Из них особенно хорошо развита XII-я пара.
Спинной мозг достигает около 70 мм в длину. Он утолщен в плечевой и крестцовой областях, причем плечевое утолщение сильно приближено к продолговатому мозгу.
Органы чувств
Из сенсорных систем ведущую роль играет обоняние, что вполне ожидаемо для подземных млекопитающих. Слышимая человеком вокализация ограничивается высоким резким писком, издаваемым в случае недовольства. Благодаря особенностям строения, среднее ухо может быть приспособлено для восприятия низкочастотных звуков и почвенных колебаний. Данная адаптация является важной для определения местоположения добычи, хищника или своего сородича. Она отмечена у самых разных живущих под землей позвоночных, включая златокротов, однако у сумчатых кротов ее наличие пока не подтверждено.
В связи с жизнью в среде, лишенной зрительных стимулов, потребность в зрении утрачена. Расположенное под кожей рудиментарное глазное яблоко имеет вид чечевицеобразной пластинки 1 мм в диаметре, лишено хрусталика и зрачка. Пигментный слой — вероятно, остаток сетчатки — окрашивает пластинку в почти черный цвет, но отдельные палочки или колбочки распознать невозможно. Глазное яблоко лежит на надкостнице слезной кости и прикрыто сверху кожей, через которую слегка просвечивает в виде темного пятнышка. Зрительный и глазодвигательный нервы отсутствуют. Сохраняются рудиментарные глазные мышцы, иннервируемые глазничной ветвью V-й пары черепных нервов.
Слезная железа очень хорошо развита, как и конъюнктивальный мешок. Вомероназальный орган (орган Якобсона) содержит значительное количество железистой ткани, пронизанной густой сетью кровеносных сосудов. Судя по всему, крупные слезные железы с их широкими протоками, утратив какое-либо значение для глаз, увлажняют носовую полость (а также вомероназальный орган) и препятствуют ее загрязнению частичками грунта.
В районе ишиотергального пятна, в коже наверху головы и в области сумки находятся специализированные группы клеток. Несмотря на то, что эти клетки расположены под роговым слоем эпидермиса, они в общих чертах напоминают вкусовые почки. Участвуют ли данные клетки в химической или тактильной чувствительности, неизвестно. Вибриссы полностью отсутствуют.
Энергообмен
Подобно многим другим живущим под землей млекопитающим, температура тела сумчатых кротов невысокая и в значительной мере зависит от температуры окружающей среды, колеблясь от 15 до 30 °C. Температура ниже 16 °C переносится плохо — животные начинают дрожать и могут погибнуть от переохлаждения. В состоянии покоя уровень обмена веществ может существенно снижаться, что способствует экономному расходованию калорий, воды и кислорода.
Энергоэффективность процессов жизнедеятельности сумчатых кротов кажется более низкой, чем у роющих плацентарных, также обладающих лабильной температурой тела и способных снижать свой метаболизм. В частности, большой сумчатый крот (Notoryctes typhlops) прокладывает новый туннель со скоростью 2-18 м/ч, теряя при этом до 81 Дж/м, тогда как у пустынного златокрота (Eremitalpa granti) эти показатели составляют соответственно 15-40 м/ч и 73 Дж/м. В обоих случаях энергоемкость способа передвижения примерно одинакова, но каким образом сумчатый крот поддерживает свой более расточительный энергетический баланс, кроме как за счет потребления очень большого количества пищи, остается неясным.
Генетика
Согласно данным молекулярных часов, около 70 тыс. лет назад род сумчатых кротов прошел через т. н. бутылочное горлышко, значительно сократив свою популяцию.