в оглавление
назад
Водные формы (продолжение)
В полном смысле водных форм, или гидробионтов, дали два отряда млекопитающих — сирены и китообразные. Разумеется, приспособление этих животных к строго водному образу жизни шло постепенно. Их древнейшие представители уверенно передвигались на четырех конечностях и значительную часть времени проводили на суше, современные же киты и сирены водную среду не покидают.

Морда дюгоня (Dugong dugon).
По степени специализации к жизни в воде сирены представляют как бы промежуточную ступень между ластоногими и китообразными. Для них характерны каплевидное тело с сильно укороченной шеей (благодаря уплощению и срастанию шейных позвонков), ластообразные передние конечности, короткие вибриссы на верхней губе, сдвинутые кверху морды ноздри, отсутствие ушных раковин, почти полная редукция шерстного покрова и задних конечностей, от которых в толще мышц сохраняются лишь рудименты тазовых и бедренных костей, а также развитие бесскелетного горизонтального хвостового плавника, служащего основным локомоторным органом. По способности к нырянию сирены не превосходят ластоногих — например, максимальное время пребывания под водой для американского ламантина (Trichechus manatus) равно 16 мин. Средняя скорость его передвижения составляет 5-8 км/час, хотя на коротких дистанциях он способен развивать скорость до 30 км/час.

Хвостовой плавник дюгоня (Dugong dugon).
Наивысшую степень приспособления к водной среде обитания обнаруживают китообразные. Торпедообразная форма тела и редукция всех выступающих частей создают у них совершенную обтекаемость. Среди современных представителей отряда шейный перехват в слабой степени выражен только у белухи (Delphinapterus leucas), у всех остальных китообразных голова полностью сливается с туловищем. Как и у большинства млекопитающих, шейных позвонков 7, но тела позвонков от II-го до VII-го сильно сплющены и сливаются так, что все вместе имеют ту же длину, что и I-й позвонок, а длина шеи не превосходит ее ширины. Парные грудные плавники представляют собой преобразованные пятипалые конечности наземных млекопитающих. При этом их проксимальные отделы сильно сокращаются, тогда как скелет кисти, собственно и образующий плавник, наоборот, вытягивается, составляя почти две трети длины всей конечности. От заднего пояса конечностей остаются лишь дериваты таза и ничтожные остатки костей самих конечностей. На хвостовом стебле из соединительной ткани развивается горизонтальный двулопастный хвостовой плавник. Хвостовые позвонки очень толстые и имеют длинные отростки, к которым прикрепляются сильные двигающие хвост мышцы. У многих видов также формируется не имеющий костной основы спинной плавник.

Плывущий кашалот (Physeter macrocephalus).
Чрезвычайно прочный эпидермис и исключительно толстый подкожный жировой слой уменьшает теплоотдачу и умеряет действие давления воды на организм китообразных. Шерстный покров у современных представителей отряда полностью утрачен (присутствуют лишь небольшие вибриссы), кожа гладкая и не имеет слизистых желез. Из других морфологических особенностей укажем уплощение очень толстой роговицы глаза; толстую склеру; шарообразный хрусталик; отсутствие слезных желез и каналов; сильное развитие гардеровых желез, жирный секрет которых защищает глаза от вредного действия соленой воды (то же значение имеет кератинизация поверхности роговицы); значительное смещение ноздрей в дорсальном направлении; особое устройство гортани, надежно страхующее дыхательные пути от попадания в них воды во время принятия пищи; огромную емкость и эластичность легких, имеющих сильно развитую мускулатуру даже в стенках альвеол.
По своей плавательной способности китообразные приближаются к рыбам. При плавании их тело интенсивно изгибается, а толчок производится мощным хвостовым плавником. С помощью вертикальных колебаний этого органа даже крупные киты могут развивать скорость движения до 40 км/ч, а максимальная скорость плавания белокрылой морской свиньи (Phocoenoides dalli) составляет 56 км/час. Находясь в движении, морские свиньи и дельфины порой кажутся совершенно неподвижными, так как вибрация их хвостового плавника происходит с колоссальной быстротой и человеческим глазом не улавливается. Помимо простых вертикальных взмахов, хвостовой плавник способен выполнять сложные винтообразные движения. Грудные плавники выполняют роль стабилизаторов и лишь в редких случаях служат органами ограниченного движения. Спинной плавник, в случае его наличия, помогает поддерживать равновесие. Помимо этого, плавникам присуща терморегуляторная функция.
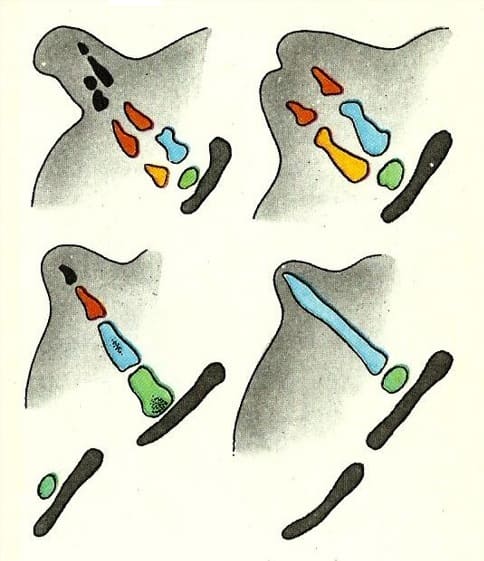
Примерно один из 10 тыс. кашалотов (Physeter macrocephalus) обладает выступающими на поверхность тела задними конечностями. Еще чаще встречаются особи с едва заметными выступами на месте задних конечностей. При анатомировании оказалось, что во всех случаях, у всех без исключения животных присутствуют тазовые кости (обозначена на рисунке серым). У самца эти кости связаны с мускулатурой пениса, а у самок — с мускулатурой влагалища и, несмотря на резко сокращенные размеры по сравнению с этими костями наземных млекопитающих, не могут считаться бесполезными, нефункционирующими. У части китов, помимо тазовой, встречаются и округлые, часто бесформенные бедренные кости, обозначенные на рисунке зеленым цветом, а также разные сочетания других костей: большой и малой берцовой (желтая и синяя), косточек заплюсны и плюсны (красные), а также пальцевых фаланг (черные). Те кости, которые встречаются не у всех особей, являются настоящими атавизмами — образованиями, не связанными с какими-либо видовыми адаптациями и проявляющимися лишь потому, что по каким-то причинам развитие данной структуры пошло по предковому пути.
При плавании на поверхности кожи китов и дельфинов формируются тонкие поперечные волны, которые гасят возникающие турбулентные завихрения и, таким образом, облегчают движение тела в водной среде. «Антитурбулентность» в данном случае обусловливается специфическим строением кожи, пронизанной большим количеством микроскопических ходов и трубочек, заполненных губчатым амортизирующим веществом. Кроме того, в процессе движения с кожи слущиваются поверхностные клетки и она еще больше сглаживается, что также уменьшает сопротивление воде. Утраченный подобным образом слой эпидермиса восстанавливается каждые 2-4 часа. Перечисленные особенности строения кожи китообразных, в сравнении с таковой наземных млекопитающих, приводят к снижению сопротивления водному потоку примерно в 100 раз. Широко известные прыжки из воды дельфины тоже совершают в целях экономии энергии — ведь воздух оказывает движущимся объектам меньшее сопротивление, чем вода. Длиннорылый продельфин (Stenella longirostris) способен на еще большее: выпрыгнув из воды, он совершает в воздухе до 7 поворотов вокруг собственной оси. Также дельфины отлично управляются с волнами: «оседлав» одну из них, они могут подолгу катиться на ее гребне, практически не совершая при этом никаких движений и почти не расходуя энергии.
При обычном плавании поступательное движение китообразных идет по волнообразной кривой, так как для возобновления запаса воздуха в легких животные вынуждены периодически всплывать на поверхность. Выдыхаемый с большой силой сжатый воздух образует специфические для крупных китов фонтаны в виде облака с примесью брызг (последние поднимаются в воздух в том случае, когда кит начинает выдох прежде, чем его ноздри успеют подняться над водой. Облако состоит не из конденсированных в холодном воздухе газов (оно образуется и в теплом воздухе тропиков), а из большого количества слизи, выбрасываемой из дыхательных путей кита.

Выпрыгнувший из воды длиннорылый продельфин (Stenella longirostris).
У кашалота (Physeter macrocephalus) вдох и выдох — процессы произвольные. Выдох он начинает еще находясь под водой. После часового погружения у кашалота наблюдают 30-40 фонтанов (выдохов) с интервалом в 25-30 с. У дельфинов дыхательный акт является безусловно-рефлекторным, действующим при смене среды вода-воздух. Благодаря такой регуляции дыхания дельфин продолжает дышать на поверхности воды даже тогда, когда находится в бессознательном состоянии. В целом надо отметить, что ритм дыхания ныряющих животных отличается нерегулярностью, т. е. имеет прерывистый характер. В то же время погружение под воду у большинства животных, включая сухопутных (кошки, собаки, куры), рефлекторно вызывает задержку дыхания — апноэ.
Благодаря своей плавучести киты в период покоя могут подолгу лежать без движения, выставив на поверхность лишь верхнюю часть туловища. С другой стороны, сделав соответствующий разгон, даже крупный кит (например, горбач (Megaptera nodosa)) способен выпрыгнуть из воды и описать в воздухе довольно большую дугу или даже сделать сальто. Несоразмерно крупную голову кашалота, которая вмещает огромный спермацетовый мешок, помимо прочего является аварийным поплавком, выносящим кита на поверхность, если запас кислорода иссяк у него на большой глубине.

Кормящийся полосатик Брайда (Balaenoptera brydei).
Киты обладают способностью глубоко нырять (по вертикали). Имеются, например, указания на случаи ныряния финвала (Balaenoptera physalus) на глубину до 360 м, клюворыла Кювье (Ziphius cavirostris) — до 3000 м, а кашалота (Physeter macrocephalus) — и более того. Время пребывания под водой у разных видов различно. Дельфины обычно погружаются не более чем на 20 мин, обычно выныривая для дыхания через промежутки не более чем 5 мин. Кашалот (Physeter macrocephalus) способен оставаться под водой до 80 мин, высоколобый бутылконос (Hyperoodon ampullatus) — до 120 мин, клюворыл Кювье (Ziphius cavirostris) — иногда до 222 мин. Усатые киты в данном отношении явно уступают зубатым: финвал (Balaenoptera physalus) может находиться под водой на протяжении 20 мин, синий кит (Balaenoptera musculus) — 50 мин, гренландский кит (Balaena mysticetus) — 60 мин. После длительного и глубокого ныряний большинству китообразных требуется достаточно долгий восстановительный период. Другим, как, например, клюворылу Кювье, достаточно провести на поверхности всего несколько минут.
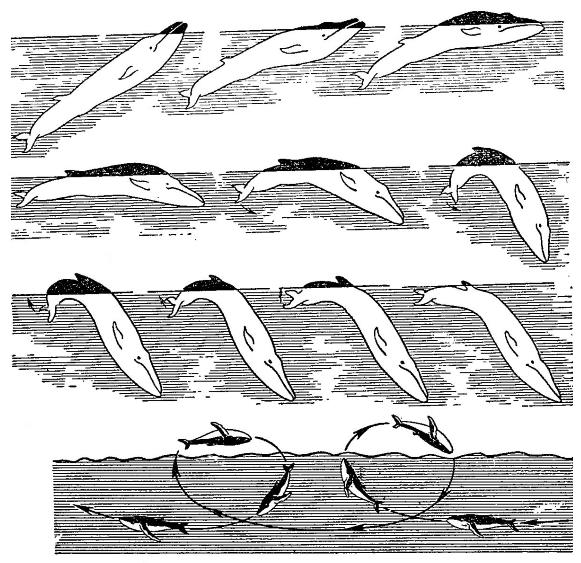
Последовательность движений финвала (Balaenoptera physalus), плывущего у поверхности воды, и горбача (Megaptera nodosa), совершающего сальто.
Ныряние связано с многократным повышением давления (на глубине 1000 м оно достигает 100 атмосфер) и длительной задержкой дыхания. Естественно, что у китообразных в связи с этим выработался целый ряд морфофункциональных адаптаций.
Количество легочных альвеол у дельфинов в три раза больше, чем у наземных животных. Как верхние, так и нижние дыхательные пути у китообразных имеют прочные стенки благодаря включению жестких хрящевых колец. Плевра у них также толстая. В легких, включая альвеолы, обнаруживаются запирательные мышечные элементы. Такая структурная особенность воздухоносных путей, легких и плевральной полости предохраняет воздух от сжатия при нырянии на большую глубину. Его давление остается близким к атмосферному, поэтому растворимость азота в крови не увеличивается. В противном случае у ныряющих животных после погружения развивалась бы кессонная болезнь — профессиональное заболевание аквалангистов.
Глубина погружения и длительность пребывания под водой некоторых водных млекопитающих (по Наумову, 1989)

Сами легкие китообразных очень вместительные — например, вычислено, что у синего кита (Balaenoptera musculus) весом в 122 т емкость легких составляет 14 тыс. л, а запас в них кислорода достигает 2800 л. Относительный объем легких значительно превышает таковой у сухопутных животных. Так, у дельфинов масса легких достигает 4 % от массы тела. Это в два с лишним раза больше по сравнению с наземными животными. Для примера, масса легких у домашнего быка (Bos primigenius taurus) равна 0,7 %, у свиньи (Sus scrofa domesticus) — 0,9 %, у лошади (Equus ferus caballus) — 1,0 %, у собаки (Canis lupus familiaris) — 1,4 %, у кошки (Felis silvestris catus) — 1,9 %. Помимо повышенной емкости, легкие китообразных отличаются высокой эластичностью. Благодаря этим особенностям они имеют увеличенную фракцию дыхательного воздуха в составе жизненной емкости легких. Например, если у наземных млекопитающих дыхательный воздух занимает около 10 % от жизненной емкости легких, то у водных он составляет 80-90 %.
Большой объем легких и их противостояние сдавливанию обеспечивает не только полную разовую вентиляцию легких, но и положительную плавучесть телу вторично водных животных. Это для них важно, поскольку у китообразных (а также ластоногих и других ныряющих животных) относительно тяжелый скелет. В костях превалирует компактное вещество при малой доле более легкого губчатого вещества. Масса костей составляет 25-30 % от живой массы водных животных.
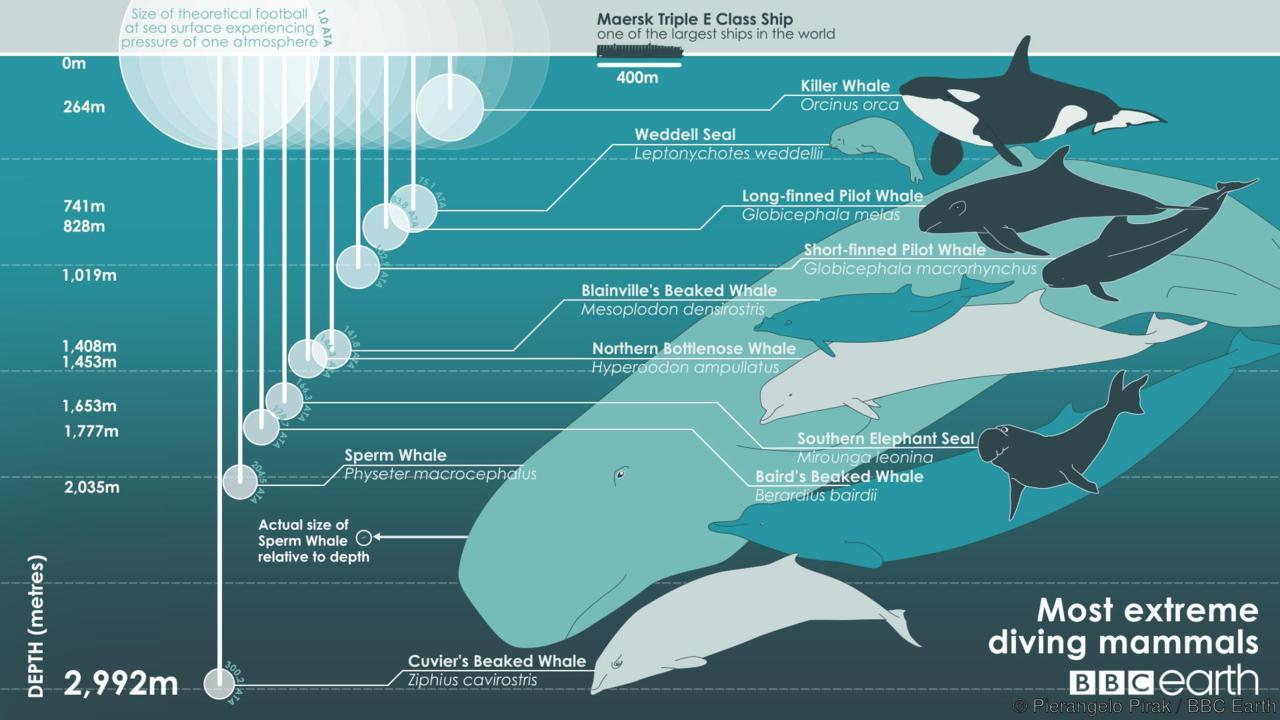
Зарегистрированная максимальная глубина погружения некоторых млекопитающих.
Наилучшие ныряльщики обладают дополнительным приспособлением для запасания атмосферного воздуха. Заполнение их легких воздухом происходит через левую ноздрю. Правая же ноздря сильно расширена и превращена в воздушный мешок, емкость которой равна емкости легких, т. е. 7-8 тыс. л у высоколобого бутылконоса (Hyperoodon ampullatus) и 12-15 тыс. л у кашалота (Physeter macrocephalus). Заполнение воздушного мешка воздухом происходит следующим образом. У поверхности воды кит производит выдох. После этого следует вдох через левую ноздрю с заполнением легких свежим воздухом. Затем левая ноздря закрывается, а воздух из легких перекачивается в воздушный мешок. Далее вдох повторяется и происходит окончательное заполнение легких атмосферным воздухом.
Сердце у водных зверей находится в условиях, которые предполагают выполнение меньшего объема работы (низкая гравитация, преобладание горизонтальных прямолинейных магистральных сосудов). Поэтому сердечный индекс у водных животных меньше, чем у сухопутных. Более того, крупные киты в хороших условиях питания накапливают большую массу жировой ткани. Поэтому за счет жировых накоплений в теле у них сердечный индекс понижается до 0,2-0,3 %. Несмотря на относительно малые размеры сердца вторичноводных млекопитающих, функционально оно выигрывает у сердца сухопутных представителей класса. Проявляется это в увеличенном систолическом выбросе и минутном объеме крови. Например, у тюленя после 10 погружений под воду систолический объем возрастает с 38 мл до 100 мл, а минутный — с 5,5 л до 14,7 л. У наземных животных эти характеристики сердца могут увеличиваться при функциональных нагрузках менее чем в два раза.
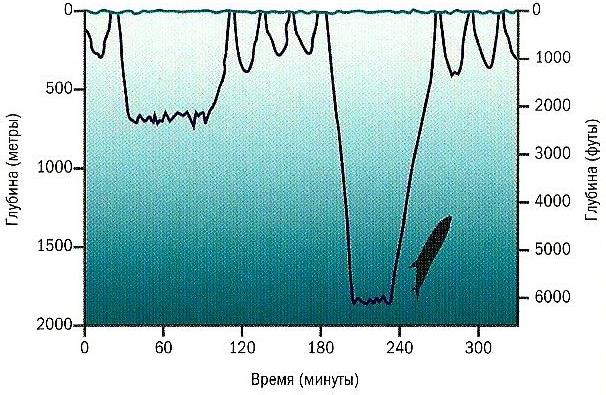
Кривая погружений клюворыла Кювье (Ziphius cavirostris).
Строение сердца ныряющих животных имеет особенности. Прежде всего бросается в глаза симметрия в развитии левой и правой половин. Если у собаки толщина стенки правого желудочка составляет всего 33 % от средней толщины стенки левого желудочка, то у кольчатой нерпы (Pusa hispida) эта разница более выражена и равняется 64 %. У гренландского тюленя (Pagophilus groenlandicus) левый и правый желудочки равны по объему и по массе. А у китов-гигантов правый желудочек больше левого. Объясняется это тем, что разветвленная капиллярная сеть огромных легких создает большое сопротивление току крови. Преодолеть это сопротивление позволяет мощная систола правого желудочка, который выглядит гипертрофированным на фоне левой половины сердца.
Ныряющие животные при нырянии проявляют функциональную брадикардию. При погружении под воду у крупных китов частота сердечных сокращений снижается с 30 до 15 ударов в минуту. У косатки (Orcinus orca) работа сердца при нырянии замедляется с 60 до 30 ударов. У тюленей брадикардия еще более выражена. У них частота сокращений сердца после погружения под воду рефлекторно понижается в 10 раз — с 60 до 6 ударов в минуту.

Дыхало синего кита (Balaenoptera musculus).
При нырянии вследствие задержки дыхания у китообразных и ластоногих диафрагма передавливает заднюю полую вену. В результате кровь скапливается в венозных сосудах, сплетениях и пазухах, разгружая таким образом сердечную мышцу. При этом ограничивается кровоснабжение и мышц тела. Скелетные мышцы переходят на потребление тканевого кислорода, который запасается при помощи миоглобина. К тому же под водой у ныряющих животных резко снижается мышечный тонус, что предполагает снижение потребления кислорода и выделения углекислого газа. Экспериментальные исследования, проведенные на тюленях, показали, что при насильственном погружении животных под воду в первую минуту отмечается повышение газообмена. Однако далее наступает длительное снижение потребления кислорода. В итоге за время пребывания под водой животные расходуют значительно меньше кислорода, чем на поверхности воды, т. е. одной из важнейших особенностей ныряющих животных является снижение кислородного запроса организма.
Интересно, что при погружении под воду у китов не изменяется снабжение артериальной кровью головного мозга и сердечной мышцы. Анастомозы кровеносных сосудов, образующие хорошо развитую «чудесную сеть», обеспечивают быстрый возврат крови к сердцу без потерь кислорода в таких мощных его потребителях, как скелетные мышцы. Таким образом, при нырянии кислород крови идет на обеспечение головного мозга и сердца. Кроме того, у водных млекопитающих отмечена низкая чувствительность мозга к углекислому газу и снижению водородного баланса крови. Поэтому морские звери могут остаться под водой до тех пор, пока не иссякнут запасы кислорода, не реагируя при этом на накопление в крови углекислоты.
У многих водных млекопитающих отмечается повышенная кислородная емкость крови. Но это справедливо не для всех ныряющих животных. Так, кислородная емкость крови китов даже ниже, чем у наземных животных. Большая кислородная емкость крови морских млекопитающих создается за счет повышенного содержания кислорода в одном эритроците, а не за счет повышения общего количества красных клеток, как в высокогорных условиях. Средние показатели гемоглобина зверей-ныряльщиков не намного выше показателей наземных животных: кашалот (Physeter macrocephalus) — 15-17 %; сейвал (Balaenoptera borealis) — 14-16 %; ламантин (Trichechus manatus) — 12-13 %.
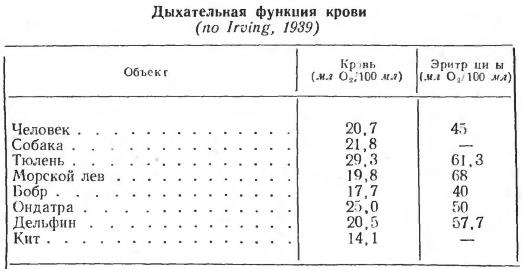
Тем не менее общие запасы кислорода в крови у ныряющих животных выше, чем у наземных. Этому способствует особое строение артериальной системы. У вторичноводных млекопитающих имеются специфические образования — артериальные сплетения, в которых депонируется насыщенная оксигемоглобином кровь. Поэтому общая кислородная емкость крови, например, у тюленя в 2 раза выше, чем у человека. У зверей-ныряльщиков имеется и более надежный способ создания запаса кислорода. Исследования показали, что у китов и ластоногих содержание миоглобина в несколько раз выше, чем у наземных млекопитающих. Так, у тюленя и быка различия по этому по этому показателю достигают 7-кратной величины. В отдельных мышцах концентрация миоглобина у вторичноводных животных в 10 раз превышает таковую у собаки.
вперед
в оглавление