Строение и функции
Внешне дейнотерии напоминали предшествовавших им продейнотериев, но были при этом гораздо крупнее их. Длина тела у представителей различных видов колебалась в пределах 3,5-7 м, рост в холке достигал 3-5 м (в среднем — 3,5-4 м), а вес составлял 8-12 т. Дейнотерии, несомненно, являлись крупнейшими наземными животными на протяжении позднего миоцена - раннего плиоцена. Внешне они несколько напоминали современных слонов, однако отличались от них своими пропорциями. Как правило, туловище дейнотериев было сравнительно укороченным и держалось на высоких, но массивных колоннообразных конечностях. Тело заканчивалось довольно длинным хвостом.
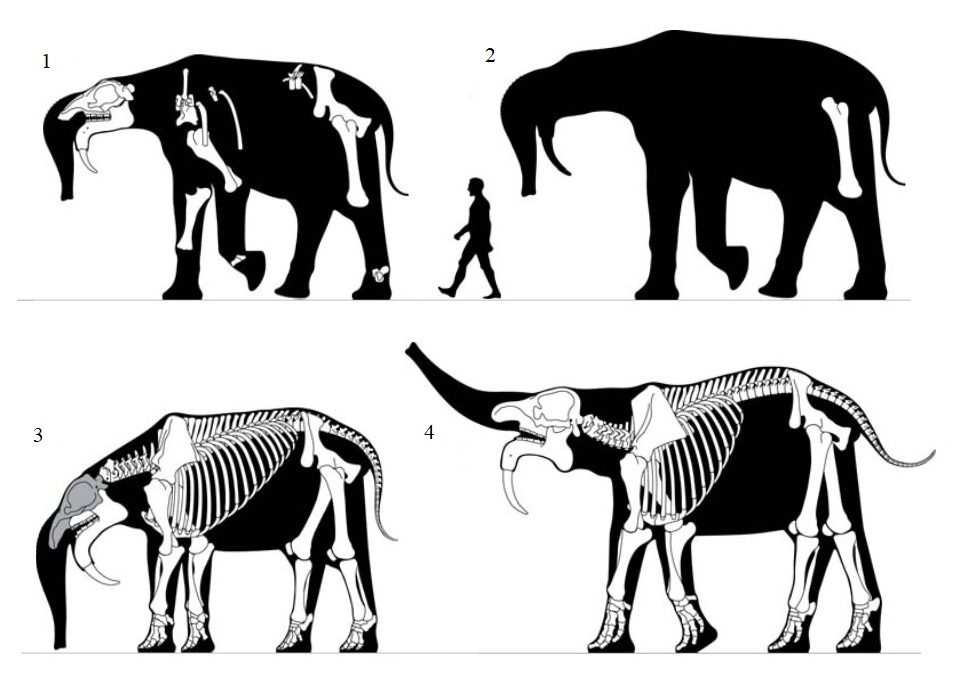
Прорисовки скелетов 4 особей дейнотериев: 1, 2 — Dеinotherium giganteum; 3, 4 — Dеinotherium proavum (3 — "Dеinotherium thraceiensis").
По сравнению с массивным телом, череп дейнотериев относительно небольшой. Его лицевой отдел длинный, почти равен по длине невысокому мозговому. Череп в большей или меньшей степени уплощен в дорсо-вентральном направлении. В профиль и сверху он имеет клиновидные очертания. Воздушные полости в черепе развиты умеренно. Вытянутые предчелюстные кости расположены параллельно изгибающемуся вниз нижнечелюстному симфизу. Альвеолы для верхних бивней отсутствуют. Впадина наверху на предчелюстных костях большая, широкая. Носовое отверстие отодвинуто от переднего конца черепа; его задний край расположен примерно на уровне переднего края глазниц. Маленькая глазница смещена вверх. Скуловая впадина большая. Сагиттальный и затылочный гребни отсутствуют. Затылок низкий, затылочная площадка сильно увеличена, расширена в латеральном направлении, уплощена в верхней части и отклонена вперед. Чешуя затылочной кости образует уступ. Верхняя часть чешуи затылочной кости в центре несет хорошо выраженные рельефные образования в виде сильно развитых бугорков, которые служили местами прикрепления мощных мышц. Следует также отметить, что уплощение затылка с наклоном вперед при высоком положении мыщелков дает животному возможность двигать головой вверх по очень большой дуге, почти под прямым углом к шее. Вероятно, подобные движения были связаны с использованием направленных вниз бивней.

Череп и нижняя челюсть дейнотерия (Deinotherium giganteum) из окрестностей Эппельхайма, Германия. Высота черепа составляет 1,2 м, ширина — 0,9 м.
Очень характерно для дейнотериев строение нижней челюсти. Обе половины нижней челюсти срастаются по симфизу до исчезновения шва. Хорошо выраженный желобок сагиттальной линии симфиза позволяет предполагать наличие длинного, мускулистого языка. Нижний край тела нижней челюсти под зубным рядом практически прямой. Восходящая ветвь широкая, отчетливо отклоняется наружу в верхнем направлении. Тело и восходящая ветвь нижней челюсти образуют прямой угол, в отличие от таковой представителей Еlephantiformes. В отличие от слонов тело нижней челюсти у дейнотериев длинное, массивное и слегка вздутое в средней части. Кроме того, их нижняя челюсть очень вытянута в передней части, а удлиненный симфизный отдел почти под прямым углом к телу нижней челюсти загибается вниз и назад и несет 2 изогнутых назад бивня, представляющие собой видоизмененные i2, округлые в сечении. Длина выступающей из челюсти части бивня может доходить до 1 м, но, как правило, меньше.
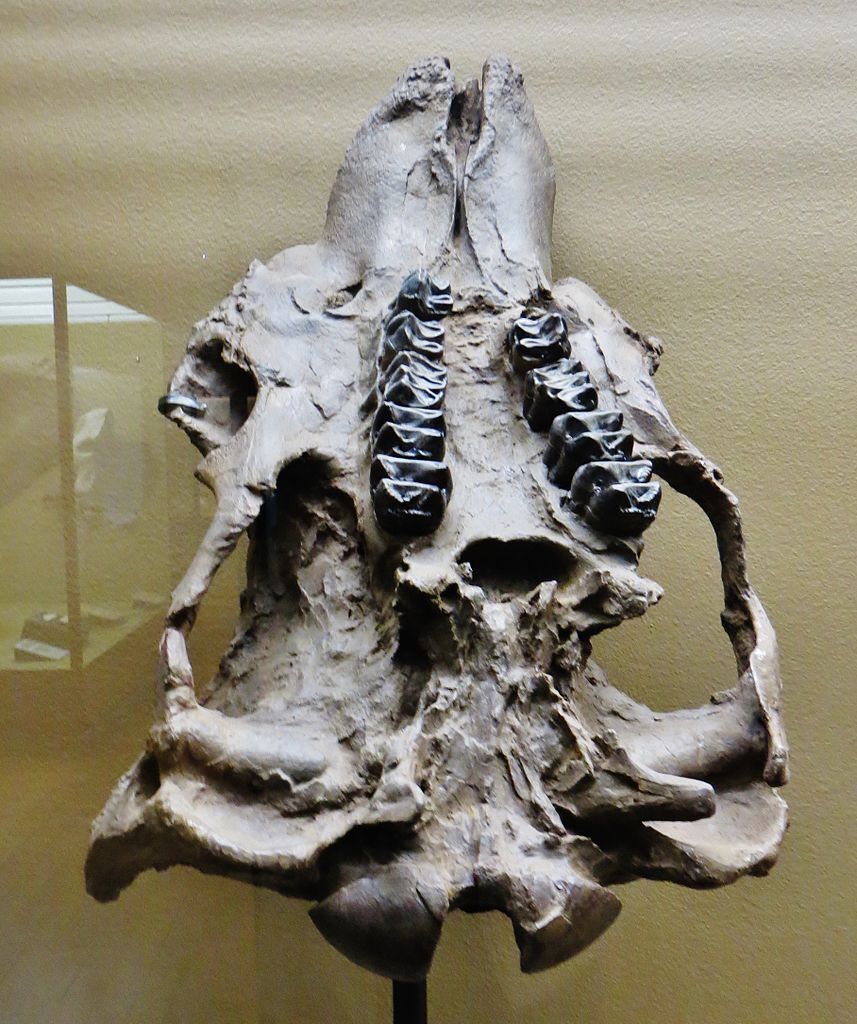

Слева: череп дейнотерия (Dеinotherium giganteum); снизу.
Справа: левая ветвь нижней челюсти дейнотерия (Dеinotherium giganteum) с острова Крит, Греция; снаружи.
Относительно назначения в жизни дейнотериев своеобразных нижних бивней высказывались самые различные предположения. Согласно воззрениям ранних авторов, причислявших данных животных к сиренам или к ленивцам, бивни якобы использовались ими в качестве якоря во время сна у речных берегов, либо служили при лазании для цепляния за ветви деревьев. Последовавшие в дальнейшем более полные находки, позволившие установить принадлежность дейнотериев к хоботным, заставили отказаться от подобных экзотических версий и позволили выдвинуть другие, более или менее вероятные. Несостоятельным выглядит утверждение, что бивни применялись прежде всего для защиты от хищников — во-первых, форма и расположение существенно ограничивают область использования бивней в качестве эффективного оборонительного оружия, а во-вторых, лучшей защитой дейнотериев являлись их гигантские размеры. Было высказано мнение, что большая величина бивней являлась вторичным половым признаком, однако фрагментарность подавляющей части находок не позволяет его подтвердить, как, впрочем, и опровергнуть. В любом случае, не исключено, что бивни играли немаловажную роль в социальной жизнь этих животных, например, выступая в качестве турнирного оружия самцов в брачный сезон.
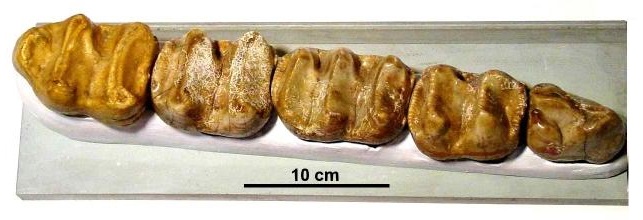
Ряд нижних щечных зубов дейнотерия (Deinotherium giganteum).
Все же наиболее вероятной выглядит точка зрения, согласно которой дейнотерии использовали свои бивни преимущественно в процессе питания. Животные могли пригибать и обламывать ими ветки деревьев, а также сдирать кору с древесных стволов. Нередко подобным образом поступают и современные слоны (правда, их щечные зубы гораздо лучше приспособлены к потреблению столь жесткого корма, как кора). В вязи с этим примечательно, что в некоторых случаях на отогнутой назад стороне бивней дейнотериев наблюдается характерный износ эмали, однако на других находках подобная истертость отсутствует. Кроме того, бивни в ряде случаев истерты на концах, с наружной и внутренней сторон, а иногда и вовсе сломаны при жизни животного. Таким образом, представляется вероятным, что использование бивней зависело от конкретной окружающей дейнотериев кормовой базы. На ряде черепов скуловые дуги несут хорошо развитые ограничивающие гребни. Направляясь назад и вниз снаружи сочленовных отростков нижней челюсти, они ограничивают ее передне-задние движения и защищают во время открывания, когда бивни подвергались значительным нагрузкам.
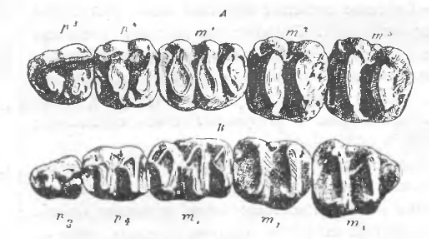
Прорисовки жевательных поверхностей верхнего и нижнего щечных зубных рядов дейнотерия (Dеinotherium giganteum) из окрестностей Эппельхайма, Германия.
Зубная формула дейнотериев: I0/i1, C0/c0, P2/p2, M3/m3. Между нижними резцами-бивнями и предкоренными имеется длинная диастема. Щечные зубы имеют очень примитивное строение — они низкокоронковые (брахиодонтные), с поперечными гребнями, образованными рядами из 2 бугорков (бунолофодонтные). Аналогичная морфология жевательной поверхности известна у тапира. Р3 и Р4, кроме 2 поперечных гребней, несут соединяющий их продольный наружный гребень. Молочные предкоренные сменялись постоянными. Pd/рd и M1/m1 имеют по 3 гребня (трилофодонтные), на M2/m2 и M3/m3 — по 2 (билофодонтные). Так как последние молочные предкоренные и M1/m1 существовали одновременно до смены, то у молодых дейнотериев могло одновременно функционировать по 2 расположенных рядом трилофодонтных зуба. Молодые предкоренные сохраняют на своей внутренней стороне бугры, которые при стирании сливаются с внешними буграми и образуют гребни. Нижние щечные зубы несколько более удлинены, чем верхние. Все они имеют по 2 корня и коронку, последняя состоит из дентина и покрыта эмалью.
Структура зубов и особенности сочленения нижней челюсти с черепом свидетельствуют о преобладании в челюстном суставе движений в сагиттальной плоскости, сопровождающихся лишь сравнительно небольшими боковыми смещениями. Все 20 щечных зубов функционировали одновременно. P1-2/p1-2 и M1/m1 использовались для раздавливания, а M2-3/m2-3 — для разрезания пищи, то есть пережевывание растительной массы было давящим и режущим, а не перетирающим. Цемента на коронках нет, однако жевательная поверхность зубов в большинстве случаев не несет следов значительного износа. Все это свидетельствует о преимущественно мягком и сочном растительном корме.
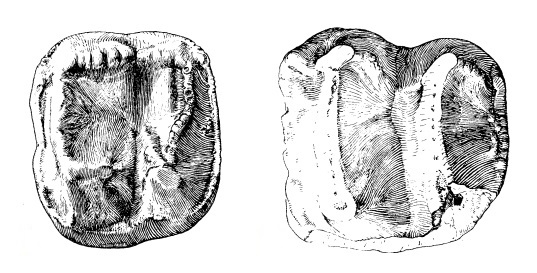
Прорисовки жевательной поверхности щечных зубов дейнотерия (Dеinotherium giganteum) из Ставрополя, Россия: слева — левый Р2, справа — левый М2.
Сравнительно с прочими хоботными, у дейнотериев мелась довольно длинная и гибкая шея, строение которой позволяло поднимать и подгибать голову вентрально, а также поворачивать ее из стороны в сторону. На такую способность указывает очень крупный атлант овальной формы, имеющий в отличие от остальных хоботных сильно развитые и массивные поперечные крылья и ряд других уникальных морфологических особенностей. Эпистрофей также очень массивен, его развитый зубовидный отросток, накладывающийся своим основанием на тело атланта, слабо уплощен, удлинен и направлен в дорсо-краниальном направлении, что отличает его от таких отростков других хоботных. Подобное его строение тоже связано с возможностью двигать головой вверх и вниз, активно работая бивнями. При нормальном положении голова располагалась горизонтально и находилась в одной осевой плоскости с шеей животного. Позвонки относительно прямые и изогнуты в меньшей степени, чем у слонов. Степень изгиба ребер и большой таз указывают на наличие объемистой грудной клетки и брющной полости.
Как и у настоящих слонов, конечности дейнотериев длинные, столбообразные. В передней конечности локтевая кость преобладает над лучевой с тонким верхним концом. В задней конечности большая берцовая кость значительно преобладает над малой, также имеющей гораздо более тонкий верхний конец, чем нижний. Кисть и стопа пятипалые, с короткими метаподиями и еще более укороченными фалангами. Карпальные кости — рядового расположения, не чередуются. Пяточная кость низкая, надпяточная совершенно плоская. I-е пальцы несколько редуцированы.

Частичный скелет дейнотерия (Dеinotherium giganteum).
Вместе с тем, в строении передней конечности и плечевого пояса наблюдаются специфические черты, указывающие на их несколько иные по сравнению с другими хоботными функции. Хорошо развитая ость делит дорсальную поверхность широкой лопатки дейнотериев на 2 выраженные ямки, имеется развитый гребень. Это указывает на хорошее развитие мышц, как выносящих переднюю конечность вперед, так и вращающих ее. Плечевая кость очень массивная, строение ее суставной головки говорит о приспособленности конечности к силовой нагрузке, действующей в сагиттальной плоскости. Имелась хорошо развитая дельтовидная мышца, способная слегка вращать плечевую кость и отодвигать ее наружу. Локтевая кость прямая, массивная и, как у других хоботных, не истончается в дистальном направлении. Локтевой отросток хорошо выражен, шероховатый, что указывает на значительное развитие разгибателя предплечья — трехглавой мышцы, крепившийся к поверхности отростка с помощью мощных сухожильных образований. В целом морфология локтевого сустава свидетельствует о значительном развитии мышц-сгибателей кисти. Форма крючковидного отростка свидетельствует о том, что при согнутой в локтевом суставе конечности животное могло совершать движения в разные стороны. У дейнотериев отмечается выраженное увеличение опорной нагрузки на наружный отдел кисти и повышение прочности запястья и запястно-пястных суставов.
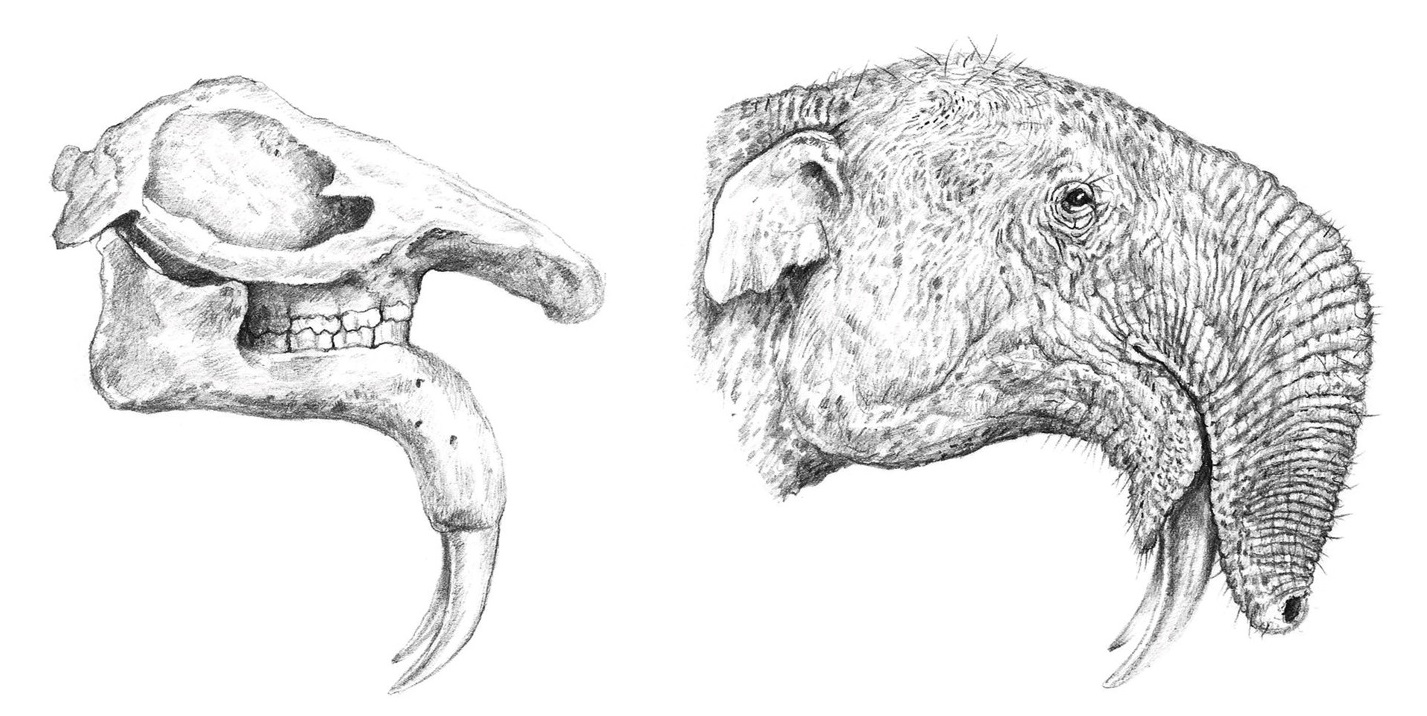
Рисунок черепа с нижней челюстью и портретная реконструкция дейнотерия (Deinotherium giganteum). Автор — В. Симеоновски.
Таким образом, передние конечности дейнотериев обладают рядом морфофункциональных особенностей, которые, по всей вероятности, были связаны с образом жизни и средой обитания этих животных. Прежде всего, в отличие от других хоботных, их передние конечности находились в отчетливо полупронированном положении с преимущественной опорой на наружные отделы. Это позволяет думать, что они были приспособлены не только к действию в сагиттальной плоскости, но могли выполнять разнообразные движения. Кисть относительно удлинена, с хорошо развитыми короткими мышцами, что позволяло животным значительно расширять ее опорную поверхность. Морфологические особенности передних конечностей дейнотериев свидетельствуют о их повышенных силовых качествах (значительное развитие мышц-сгибателей и мышц-разгибателей, отчетливо выраженная асериальность в сочленениях запястья и запястно-пястного суставов, выраженность костных упоров гребней и др.). Очевидно, это было связано с большой массой тела животных, а также, возможно, со специфическим типом использования конечностей во время добывания пищи.
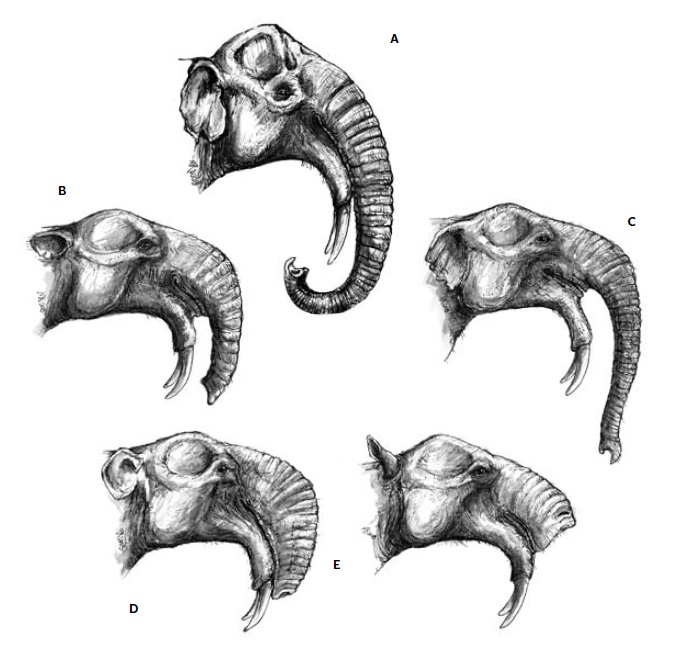
Портретные реконструкции дейнотерия (Dеinotherium giganteum) согласно представлениям различных исследователей: А — самые старые реконструкции; В — по Осборну (1910 г.); С — по Абелю (1922 г.); D — по Энгессеру с соавт. (1996 г.) и Маркову с соавт. (2001 г.); Е — по Грегору с соавт. (2000 г.).
Значительный размер и специфическая форма отодвинутого назад носового отверстия, а также присутствие выраженных следов канала VII-й пары лицевых нервов предполагают наличие более или менее развитого хобота. Хобот дейнотериев конвергентен таковому слонов и развился самостоятельно (по крайней мере, разделение предковых форм произошло еще до формирования полноценного хобота у слоновых). Очевидно, хобот дейнотериев был образован не только носом (как у слонов), но и мясистой верхней губой, хотя полной уверенности в этом нет. Его длина и форма и по сей день служат у исследователей предметом споров. На реконструкциях различных авторов можно увидеть как дейнотериев совсем без хобота, так и с длинным хоботом, напоминающим слоновий, а также всевозможные промежуточные варианты. При более внимательном рассмотрении выясняется, что дейнотерии существенно отличаются от слоновых (Elephantidae) строением предчелюстной кости, которая у них сплющена с верхней стороны и ориентирована более-менее горизонтально. Такая ее морфология не обеспечивает площади, достаточной для крепления мощной мускулатуры, образующей длинный хобот, подобный таковому у современных слонов. Поэтому некоторые реконструкторы изображают дейнотерия с коротким хоботом, несколько напоминающим хоботок тапира, или же помещают на носовой части его черепа мясистое образование, представляющее собой разросшуюся верхнюю губу.

Один из вариантов портретной реконструкции дейнотерия (Deinotherium giganteum).
Все-таки, настоящий хобот у дейнотериев вероятнее всего все же был, но при этом он не являлся столь же длинным и гибким, как у современных слонов. Очевидно, длинные конечности и шея, а также использующиеся для притягивания пищи нижние бивни компенсировали относительно небольшую длину хобота. В любом случае, ее должно было хватать для удаления застрявшей при кормлении между бивнями растительности и для использования на водопое. Учитывая явные адаптации дейнотериев к питанию строго качественным древесным кормом, представляется вероятным, что на конце хобота, как и у слонов, имелось особое хватательное устройство из одного или нескольких пальцевидных отростков. Вероятно, что они были развиты даже еще больше, так как являлись, по сути, единственным инструментом, дающим животным возможность точно манипулировать с мелкими кормовыми объектами.